20 책임저자:윤경식, 130-701, 서울시 동대문구 회기동
경희대학교 의과대학 생화학분자생물학교실 Tel: 02-961-0388, Fax: 02-965-6349
E-mail: [email protected]
접수일:2009년 2월 13일, 게재승인일:2009년 2월 26일
Correspondence to:Kyung-Sik Yoon
Department of Biochemistry and Molecular Biology, Medical Science and Engineering Research Center for Bioreaction to Reactive Oxygen Species, School of Medicine, Kyung Hee University, Hoegi-dong, Dongdaemun- gu, Seoul 130-701, Korea
Tel: +82-2-961-0388, Fax: +82-2-965-6349 E-mail: [email protected]
Effects of Tamoxifen and Ghrelin in ER-positive MCF-7 and ER-negative MDA-MB231 Breast Cancer Cells
Young Eun Choi1,2, Chang Hoon Cho1,2, Hye Ok Kim1,2, Yong Hwa Jo1,2, Jun-Kyu Song1,2, Hyung Hwan Baik1,2, Wonchae Choe1,2, Insug Kang1,2 and Kyung-Sik Yoon1,2
Departments of 1Biochemistry and Molecular Biology (BK21 Project), 2Medical Science and Engineering Research Center for Bioreaction to Reactive Oxygen Species, School of Medicine, Kyung Hee University, Seoul 130-701, Korea
Tamoxifen (Tam) is widely used in chemotherapy of breast cancer. It inhibits proliferation and induces apoptosis of breast cancer cells by estrogen receptor (ER)-dependent modulation of gene expression. ER involves with AMPK and MAPK pathway. Ghrelin is growth hormone and binds to growth hormone secreatagogue receptor1 (GHSR-1) that have estrogen receptor similar pathway. We studied to Tamoxifen, Estadiol & Ghrelin effects by MTT assay for cell viability and western blot for ERK/p38 phosphorylation in ER-positive MCF-7 (ER+) and ER-negative MDA-MB231 (ER−). We showed to MCF-7 cell death induction by Tamoxifen dose-dependent manner. But we can see inhibition of Tamoxifen effect by Estradiol and Ghrelin. ERK phosphorylation increased 2.9∼18.5-fold by dose-dependent Tamoxifen in both ER+ MCF-7 cells and 7.8-fold by 7μM Tamoxifen treatment in ER− MDA-MB231 cells. ERK phosphorylation was induced by Estadiol and Ghrelin in only ER+ MCF-7 cells, but p38 phosphorylation was not affected by Estradiol, Ghrelin and Tamoxifen in breast cancer cells. In conclusion, ERK activation of MAPK by Tamoxifen for cell death and Ghrelin in ER-positive MCF-7 cell line was observed. (Cancer Prev Res 14, 20-27, 2009)
Key Words: MCF7, MDA-MB231, Breast cancer, Tamoxifen, Ghrelin, Estrogen receptor
INTRODUCTION
Estradiols (E2) are key regulators for normal growth and differentiation of mammary glands as well as malignant progression of breast cancer.1) Actions of estrogen are mediated through an interaction with its intracellular receptor, a member of the steroid/thyroid/retinoid receptor gene superfamily.2) E2
exerts its effects by binding to Estrogen receptor (ER) that functions as transcription factor controlling cell proliferation and differentiation through MAPK and Akt.3) ER was thought to be the only form of nuclear receptor able to bind estrogen and mediates its hormonal effects in their target tissues.4) ER−
positive (ER+) breast cancer is estrogen-dependent so reducing estrogen levels or altering the activity of the receptor can induce these cancers to regress.5)
Cumulative analysis of tumor biopsies has shown that ERs are present in 65% of all human breast tumors.6) It has long been known that breast cancer tumors which express the ER+
behave in a fundamentally different fashion than ER-negative (ER−) tumors with regard to their response to hormonal therapies.7) It has been proposed that ER is mediated through growth factor receptor.8) Growth factor receptor induces signal transduction in kinase pathways such as ERK which phosphorylates and activates nuclear ER.9)
Tamoxifen (Tam) is a nonsteroidal selective ER modulator
widely used in the chemotherapy of breast cancer. Use of Tam is especially indicated for postmenopausal women who have ER+
breast cancer. Tam acts primarily through ER by modulation of gene expression. It was reported that ERK phosphorylation induced cell death rapidly by Tam.10)
Ghrelin, a gut-brain peptide, was identified as an endogenous ligand for the growth hormone secretagogue receptor 1 (GHS-R1), and stimulates food intake and adiposity.11) Ghrelin’s basic pathway has been reported to induce energy balance and cell proliferation through AMPK and MAPK. However, research of Ghrelin rarely investigated the downstream effects in breast cancer. Ghrelin inhibits breast cancer cell proliferation in vitro.12) Cell growth induced by Ghrelin in breast cancer cells was observed in this study.
Our aim for the present work is to characterize the up-stream events leading to the previously observed cell death in breast cancer cells by Tam and study the effect of E2 on Tam-induced rapid death of breast cancer cells. We studied the activation of MAPK as a possible mechanism by which E2 opposes the effects of Tam. Activation of ERK1/2 is generally considered to lead to cell survival,13) and there is evidence that several cell types ERK1/2 may also transmit proapoptotic signals.14) In addition to phosphorylation of ERK within the cell, represents another level at which the sensitivity of the MAPK signaling pathway might be regulated13) In this study, as mentioned before, ghrelin inhibits cell proliferation in breast cancer cell line and E2 and ghrelin induced MAPK activation. Thus, effects opposing Tam through ER of ghrelin treatment through GHS-R1 were examined. So far, Tam and E2 co-treated effects are as well as studied but not Ghrelin with Tam or/and E2. Therefore, we investigated cell viability and MAPK family ERK, p38 phosphorylation levels with combined two molecules in ER+/−breast cancer cell lines.
MATERIALS AND METHODS 1. Cell culture
The MCF-7 and MDA-MB231 human breast tumor cell lines were originally obtained from the laboratory of molecular biology (Kyung Hee University School of Medicin, Seoul). The cells were cultured in RPMI 1,640 medium supplemented with 10% fetal bovine serum in a humidified 5% CO2 atmosphere at 37°C. For the treatment, breast cancer cells in serum-free medium were stimulated with Tamoxifen (T5648, Sigma, USA). When Estradiol (E8875, Sigma, USA) and Ghrelin (PGH-3627-PI,
Peptides international, USA) were used as treatment, breast cancer cells were grown in phenol red-free RPMI 1,640 culture medium supplemented (0.5%) with dextran charcoal-treated FBS (dcFBS) for two days and were stimulated with E2 and Ghrelin in a serum-free medium for 3 h.
2. Cell death analysis
The MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazo- lium bromide, Sigma, USA) assay was carried out to check cell viability. Cells were plated at a density of 1×105 cells/well in a 6-well plate and viability was assessed 20 min after Tam, E2 and Ghrelin treatment. MTT (5 mg/ml) was added to the medium for 30 min. Then, the supernatant was removed, and the cells were dissolved using DMSO. The absorbance was measured at 540 nm with an ELISA microplate reader (Multiskan EX; ThermolLabsystems, USA).
3. Western blot analysis
Whole-cell lysates of breast cancer cells were prepared in lysis buffer (50 mM Tris, 150 mM NaCl, 1% triton X-100, 50 mM NaF, 5 mM sodium pyrophosphate, 1 mM EDTA, 1 mM EGTA, 1 mM DTT, 0.1 mM PMSF, 5μg/ml soybean trypsin inhibitor, pH 7.4) and centrifuged to remove cellular debris. Samples were assayed for protein concentration using the Lowry (Bio-Rad DC protein assay kit) assay. Proteins samples (40μg for each) were separated by 10∼12% SDS- PAGE and transferred to nitrocellulose transfer membranes.
The membrane was blocked with 5% non-fat dry milk in TBS-T solution. The blots were incubated with primary antibodies for ERα, p-ERK1/2, ERK1/2, p-p38, p38 (cell Signaling, Beverly, MA) in TBS-T overnight at 4° or for 1h at room temperature, nitrocellulose transfer membranes were rinsed and incubated with the appropriate secondary antibodies.
The blots were detected using the Western blotting luminal reagent.
4. Density analysis
Image J (Image processing and analysis in java, USA) was used to analyze the results of the western blot.
5. Statistical analysis
All data were expressed as the average SD from three independent experiments and all calculations were down in triplicate for each experiment. Statistical analysis was performed
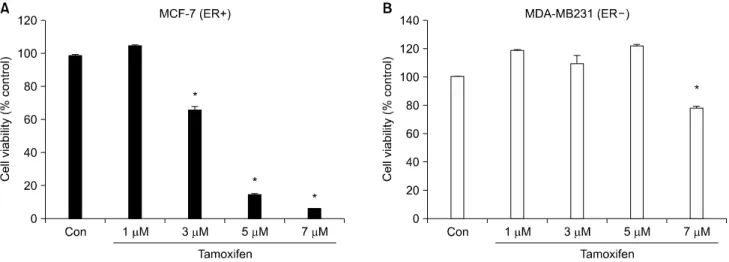
Fig. 1. Cell viability of MCF-7 and MDA-MB231 cells treated with Tam in a dose-dependent manner. MCF-7 (ER+) (A) and MDA-MB231 (ER−) cells were incubated in serum free medium for 3 h and treated with 1, 3, 5, 7μM concentrations of Tam for 20 min (B). Cell viability was analyzed by MTT assay. The results are representative of three independent experiments. *p<0.05 vs. control.
using the Student’s t-test. p-values of less than 0.5 were re- garded as statistically significant.
RESULTS
1. Tamoxifen treated ER+ MCF˗7 cells reduced cell viability in a dose˗dependent manner
The effect of Tam on cell viability in ER+ MCF-7 and ER−
MDA-MB231 breast cancer cell lines was examined. After in- cubating for 3 h MCF-7 and MDA-MB231 cells in serum-free medium, the cells were treated with 1, 3, 5, 7μM Tam for 20 min. At the end of 20 minutes, cell death was determined by the MTT assay and measured with a ELISA reader. Control cells were treated with vehicle (DMSO) only.
Cell viability of ER+ MCF-7 cell was not affected at a 1μM concentration of Tam but was reduced 30.3% (+/−0.14) by a 3μM concentration of Tam. 85.6% (+/−0.28) of the cells were dead at a 5 and 7μM concentration of Tam in MCF-7 (Fig. 1A). But cell viability of ER− MDA-MB231 was not affected by TAM at a 1, 3 and 5μM concentrations and the cell death by Tam at a 7μM concentration was only 20.4%
(+/−0.8) of the cells (Fig. 1B).
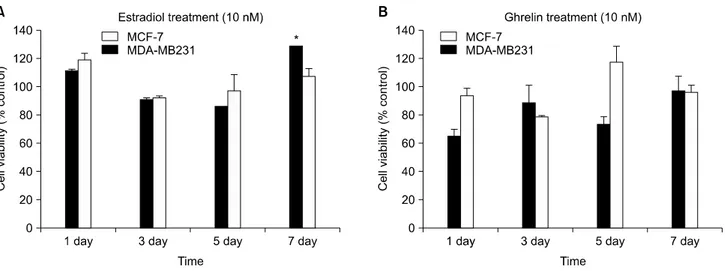
2. E2 induced cell proliferation in ER+ MCF˗7 but Ghrelin did not effect cell proliferation in breast cancer cells
ER+ MCF-7 and ER− MDA-MB231 cells were incubated
in phenol red-free medium with dcFBS for 3 h. The cells were treated with 10 nM E2 and 10 nM Ghrelin for 1, 3, 5 and 7 days. The concentrations of E2 and Ghrelin were within normal limits.10,15) Cell viability was determined by MTT assay and measured with a ELISA reader.
Cell proliferation was induced [21.2% (+/−1.2)] at day 7 in ER+ MCF-7 by E2 but not by ER− MDA-MB231 (Fig.
2A). Treatment with Ghrelin did not affect MCF-7 and MDA- MB231 cells (Fig. 2B). In other studies, cell proliferation was inhibited by Ghrelin treatment.16)
3. E2 and Ghrelin antagonize Tam˗induced cell death of ER+ MCF˗7 cells but not ER- MDA˗
MB231 cells
The co-effect of either E2 or Ghrelin with Tam on cell viability in ER+ MCF-7 and ER− MDA-MB231 breast can- cer cell lines was examined. ER+ MCF-7 and ER− MDA- MB231 cells were incubated with dcFBS (0.5%) for two days then treated with either E2 or Ghrelin and Tam for 20 min and afterwards changed to serum free medium. Cell death was determined by a MTT assay and measured with a ELISA reader.
Tam alone induced cell death by 47.4% (+/−3.2) in ER+
MCF-7 cells, but cell viability increased by co-treatment with either Tam and E2 or Tam and Ghrelin (Fig. 3A). Cell viability was not affected by any combination of Tam, E2 and Ghrelin in ER− MDA-MB231 cells (Fig. 3B). These results suggest
Fig. 2. Cell proliferation induced by E2 but Ghrelin did not effect in MCF-7 and MDA-MB231. The ER+ MCF-7 and ER−
MDA-MB231 cells were incubated with dcFBS (0.5%) for 1, 3, 5 and 7 days. The medium was changed every two days and treated with E2 and Ghrelin after each medium change. Cell viability was analyzed by MTT assay. The data are representative of three independent experiments. (A) 10 nM E2 treatment, (B) 10 nM Ghrelin treatment. *p<0.05 vs. control.
Fig. 3. Cell viability of MCF-7 and MDA-MB231 cells treated with either E2 or Ghrelin and Tam. ER+ MCF-7 and ER− MDA-MB231 cells were incubated with dcFBS (0.5%) for two days and then treated with either E2 or Ghrelin and Tam for 20 min after which the medium was changed to serum free. Cell viability was analyzed by MTT assay. Results are representative of three independent experiments. (A) MCF-7 (ER+), (B) MDA-MB231. *p<0.05 vs. control.
that E2 and Ghrelin antagonize Tam-induced cell death in ER+
MCF-7 cells.
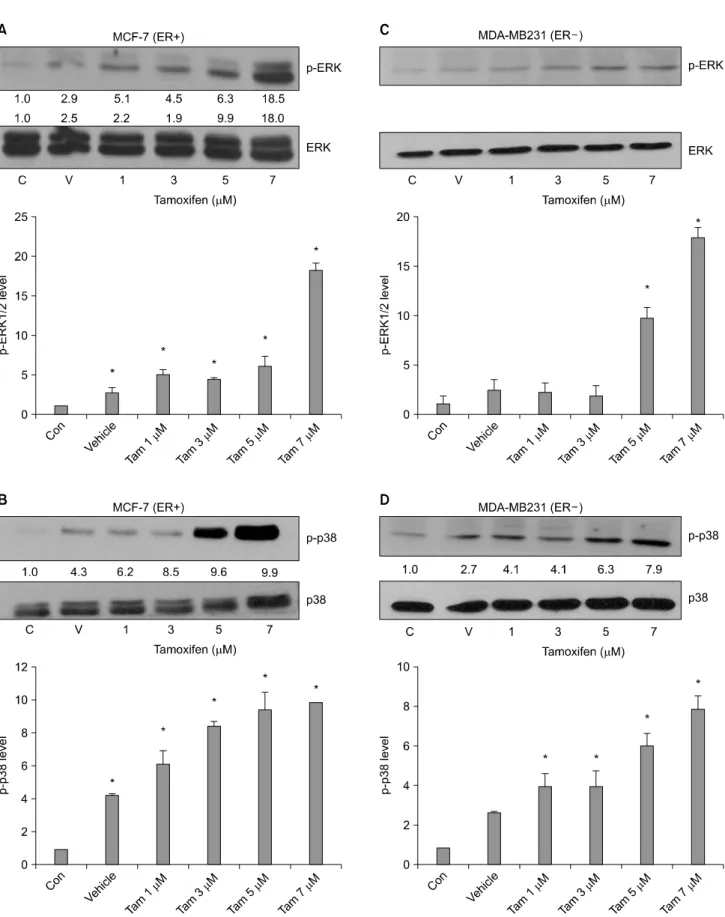
4. ERK and p38 phosphorylation were increased in a dose˗dependent manner by Tam in both ER+ and ER- breast cancer cells
To examine the effect of Tam on ERK and p38 pho- sphorylation in ER+ MCF-7 and ER− MDA-MB231 breast cancer cell lines, the cells were incubated in serum-free medium for 3 h. The cells were then treated with 1, 3, 5 and 7μM
concentrations on Tam for 20 min. Protein levels of ERK and p-38 phosphorylation were determined by western blot. We showed that non-treated sample density as a control. ERK phosphorylation increased 2.9∼18.5-fold in a dose-dependent manner by Tam in both ER+ MCF-7 cell (Fig. 4A) and 7.8-fold in ER− MDA-MB231 cells with treatment of 7μM Tam (Fig. 4C). Also, p38 phosphorylation increased 4.3∼
9.9-fold in a dose-dependent manner by Tam in ER+ MCF-7 cells (Fig. 4B) and 2.7∼7.9-fold in ER− MDA-MB231 cells (Fig. 4D).
Fig. 4. Tam induced phophorylation of ERK and induced p38 phosphorylation in ER+ MCF-7 and ER− MDA-MB231 cells. The ER+ MCF-7 and ER− MDA-MB231 cells were incubated in serum-free medium for 3 h and then treated with 1, 3, 5 and 7μM of Tam for 20 min. Whole-cell extracts were analyzed for ERK1/2 and p38 phosphorylation by Western blot. (A), (B) MCF-7 (ER
+)/(C), (D) MDA-MB231(ER−). Results are representative of three independent experiments. C: control, V: vehicle. *p<0.05 vs.
control.
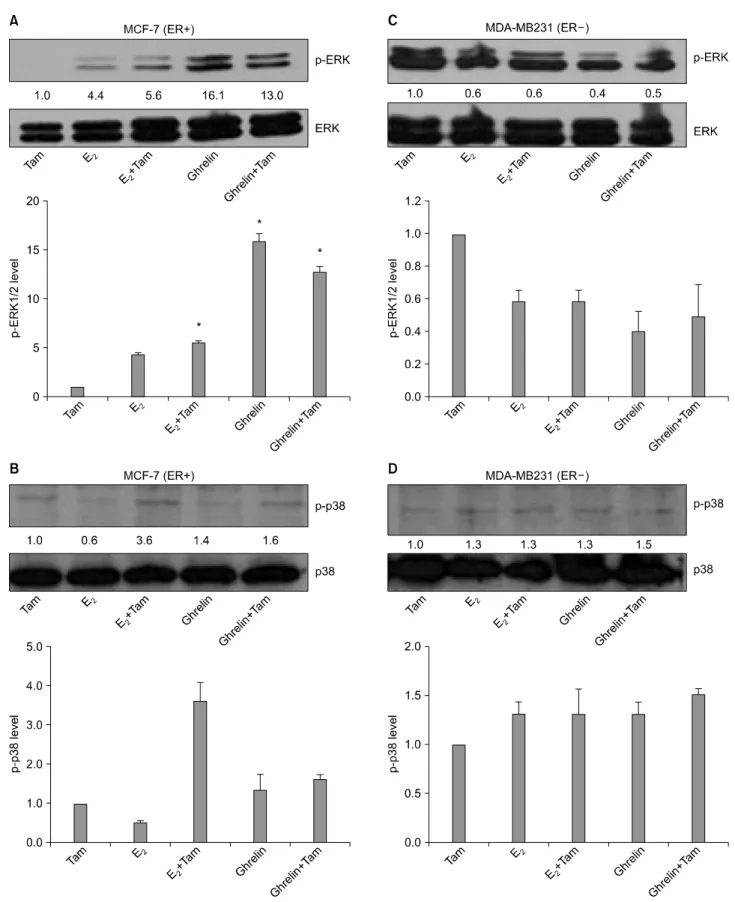
Fig. 5. Effects of E2, Ghrelin and Tam in ER+ MCF-7 and ER− MDA-MB231 cells on ERK and p38 phosphorylation. ER+
MCF-7 and ER− MDA-MB231 cells incubated with dcFBS (0.5%) for two days then treated with 10 nM E2, 10 nM Ghrelin and 5μM Tam for 20 min after which the medium was changed to serum-free medium. Whole-cell extracts were analyzed for ERK1/2 and p38 phosphorylation by Western blot (A), (B) MCF-7 (ER+)/(C), (D) MDA-MB231 (ER−). Results are representative of three independent experiments. *p<0.05 vs. control.
5. ERK phosphorylation was induced by Ghrelin in ER+ MCF˗7 cells only; but, p38 pho- sphorylation was not induced by E2, Ghrelin and Tam in breast cancer cells
To examine the co-effect of E2 or Ghrelin with Tam on ERK and p38 phosphorylation in ER+ MCF-7 and ER− MDA- MB231 breast cancer cell lines, ER+ MCF-7 and ER−
MDA-MB231 cells were incubated with dcFBS (0.5%) for two days then treated 10 nM E2 or 10 nM Ghrelin and 5μM Tam for 20 min after which the medium was changed to serum-free medium. We showed that 5μM Tam treated sample density as a control. Protein levels for ERK and p-38 phosphorylation were determined by western blot. ERK phosphorylation increased 11.3-fold by Ghrelin treatment in ER+ MCF-7 cells (Fig. 5A) and had no effect in ER− MDA-MB231 cells (Fig.
5C). But p38 phosphorylation levels were not changed significant by treatment with E2 and Ghrelin in ER+ MCF-7 cell (Fig. 5B) and ER− MDA-MB231 cells (Fig. 5D).
DISCUSSION
The purpose of this study was to investigate the effects of Tam and Ghrelin in ER+ MCF-7 and ER− MDA-MB231 breast cancer cell lines.
Our data suggests that cell death was induced in a dose-dependent manner when treated with Tam in ER+
MCF-7 cell. Cell death in ER− MDA-MB231 cells treated with 7μM Tam was noted. A 7.8-fold induction was observed.
Other studies have reported this effect before. But only cell viability was examined at the time.
So, we analyzed the protein levels in the dead cells induced by Tam. The proteins we targeted were MAPK family members (ERK, p38, JNK). The results showed that pho- sphorylated ERK and p38 was induced by Tam in ER+
MCF-7 and ER− MDA-MB231.
Consequently, ERK and p38 proteins were activated in MCF-7 and MDA-MB231 but activated proteins induced by Tam was dependent on whether it was ER+ or ER−. Thus, Tam not only initiates its toxic effect in the cells through the influence of ER but also other pathways in MDA-MB231 cells.
If cell death was not induced through the ER in ER+ MCF-7 and ER− MDA-MB231 cell lines, other pathways will be investigated to confirm the results that Tam induced cell death
was dose-dependent.
Cell proliferation was observed after 7 days by E2 treatment which suggests cell proliferation was induced through ER.
Co-treatment of cells with Tam and E2 showed that cell death was antagonized. This result was reported in other studies. We think competitive binding might play a role because Tam and E2 function through and activate downstream targets by the ER.
In our study, we showed the effects of Ghrelin in ER+
MCF-7 and ER− MDA-MB231 cells. ER+ MCF-7 and ER−
MDA-MB231 cells were ideal for this comparative study due to the ER+ or ER− groups. We looked at Ghrelin treatment and its different effects on our cell lines ER+ MCF-7 and ER−
MDA-MB231. GHS-R1 was expressed and active in ER+
MCF-7 and ER− MDA-MB231 cells. Ghrelin was a unique area of study. Various reports about its ability have been reported. But certain pathway and its targets have not been elucidated. It has been reported that Ghrelin induces proliferation in human aortic endothelial cells via ERK1/2 and PI3K/Akt activation.15) But it has also been reported that Ghrelin inhibits cell proliferation in regular breast cancer cell lines.16) Thus, we looked at the effects of Ghrelin and Tam in cells. Cell viability was reduced more when the cells were co-treated with Tam and Ghrelin than only Tam treatment in ER+ MCF-7 cell. ERK phosphorylation was induced by Ghrelin. ERK phosphorylation was lowered by co-treatment of Tam and Ghrelin than when the cells were only treated with Ghrelin. Our results show that cell viability did not change after Ghrelin only treatment from 1 to 7 days in ER+ MCF-7 and ER− MDA- MB231 cells.
As a result, ERK phosphrylation by Tam activated for Tam resistance.17) Cell proliferation may not be related strongly to the Ghrelin, but cell proliferation was inhibited by Ghrelin. In our experiment, cell proliferation was inhibited by Ghrelin in the breast cancer cell lines. It may be that Ghrelin antagonizes Tam resistance because ERK phosphorylation is reduced with treatment of Tam and Ghrelin more than when the cells are only treated with Ghrelin. The question about Ghrelin is not discussed in detail because of the lack research about Ghrelin.
Ghrelin may have a significant role in breast cancer. The effect observed in our breast cancer cell lines was that the activity of Ghrelin was through the MAPK pathway and comparable to the well-known E2 pathway. Based on this account, we think that we may solve the problem of the role Ghrelin gradually
based on this result.
Ghrelin must establish concrete roles breast cancer cell line because Ghrelin has various effects according cell lines.
Additional experiments will study the inhibition of ER and GHS-R1. Also, we will observe activation of specific sites of the MAPK pathway and its downstream targets.
CONCLUSION
Tam through ER induced cell death in ER+ MCF-7 cells and ER− MDA-MB231 cells. E2 and Ghrelin antagonized Tam- induced cell death. Tam, E2 and Ghrelin induced ERK activation of MAPK. Of note, ERK is related to cell prolife- ration but Tam resistance activated ERK in cell death.
We will take to elucidate the mechanisms of Tam/Ghrelin- induced rapid signaling for better understanding how Ghrelin interferes with the cell signaling network and affects normal physiological function.
ACKNOWLEDGEMENT
This research was supported by the Kyung Hee University Research Fund in 2004 KHU-20040348.
REFERENCES
1) Nadine Platet, Anne Marie Cathiard, Michel Gleizes, Marcel Garcia. Estrogens and their receptors in breast cancer pro- gression: a dual role in cancer proliferation and invasion. Crit Rev Oncol Hematol 51, 55-67, 2004.
2) Kyung-Chul Choi, Sung Keun Kang, Chen-Jei Tai, Nelly Auersperg, Peter C.K Leung. E2 up-regulates antiapoptotic Bcl-2 messenger ribonucleic acid and protein in tumorigenic ovarian surface epithelium cells. Endocrinology 142, 2351-2360, 2001.
3) Carolyn M Klinge. Estrogen receptor interaction with estrogen response elements. Nucleic Acids Res 29, 2905-2919, 2001.
4) Sietse Mosselman, Jan Polman, Rein DijTkema. ERβ:
identification and characterization of a novel human estrogen receptor. FEBS Lett 392, 49-53, 1996.
5) Wenlin Shao, Myles Brown. Advances in estrogen receptor biology: prospects for improvements in targeted breast cancer
therapy. Breast Cancer Res 6, 39-52, 2004.
6) Paik S, Hartmann DP, Dickson RB, Lippman ME.
Antiestrogen resistance in ER positive breast cancer cells.
Breast Cancer Res Treat 31, 301-307, 1994.
7) Murphy L, Cherlet T, Lewis A, Banu Y, Watson P. New insights into estrogen receptor function in human breast cancer. Ann Med 35, 614-631, 2003.
8) Edward J. Filardo, Jeffrey A. Quinn, A. Raymond Frackelton, Jr., Kirby I. Bland. Estrogen action via the G protein-coupled receptor, GPR30: Stimulation of adenylyl cyclase and cAMP-mediated attenuation of the epidermal growth factor receptor-to-MAPK signaling axis. Mol Endocrinol 16, 70-84, 2002.
9) Ellis R. Levin. Integration of the Extranuclear and Nuclear Actions of Estrogen. Mol Endocrinol 19, 1951-1959, 2005.
10) Aiping Zheng, Anu Kallio, Pirkko Härköe. Tamoxifen- Induced rapid death of mcf-7 breast cancer cells is mediated via extra- cellularly signal-regulated kinase signaling and can be abro- gated by estrogen. Endocrinology 148, 2764-2777, 2007.
11) Ichiro Sakata, Toru Tanaka, Mami Yamazaki, Takashi Tani- zaki, Zhao Zheng, Takafumi Sakai. Gastric estrogen directly induces ghrelin expression and production in the rat stomach.
J Endocrinol 190, 749-757, 2006.
12) Chrysanthia A Leontiou, Giulia Franchi, Márta Korbonits.
Ghrelin in neuroendocrine organs and tumours. Pituitary 10, 213-225, 2007.
13) Marshall CJ. Specificity of receptor tyrosine kinase signaling:
transient versus sustained extracellular signal-regulated kinase activation. Cell 80, 179-185, 1995.
14) Madalina Stanciu, Donald B. DeFranco. Prolonged nuclear retention of activated extracellular signal-regulated protein kinase promotes cell death generated by oxidative toxicity or proteasome inhibition in a neuronal cell line. J Biol Chem 277, 4010-4017, 2002.
15) Rossi F, Castelli A, Bianco MJ, Bertone C, Brama M, Santiemma V. Ghrelin induces proliferation in human aortic endothelial cells via ERK1/2 and PI3K/Akt activation. Pe- ptides 29, 2046-2051, 2008.
16) Paola Cassoni, Mauro Papotti, Corrado Ghe, Filomena Cata- pano, Anna Sapino, Andrea Graziani, Romano Deghenghi, Thomas Ressmann, Ezio Ghigo, Giampiero Muccioli.
Identification, characterization, and biological activity of specific receptors for natural (Ghrelin) and synthetic growth hormone secretagogues and analogs in human breast car- cinomas and cell lines. J Clin Endocrinol Metab 86, 1738-1745, 2001.
17) Riggins RB, Schrecengost RS, Guerrero MS, Bouton AH.
Pathways to tamoxifen resistance. Cancer Lett 256, 1-24, 2007.