Plasmid profiling of multi-drug resistant Vibrio sp. isolated from influent and effluent water samples of fish farms in Jeju, South Korea
7
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
수치

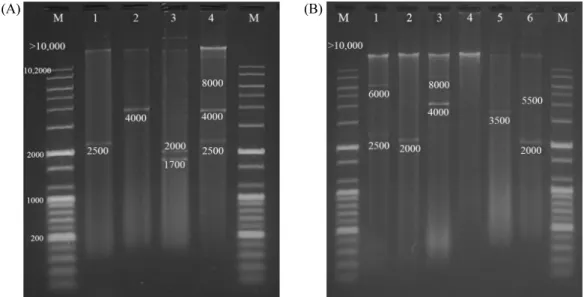
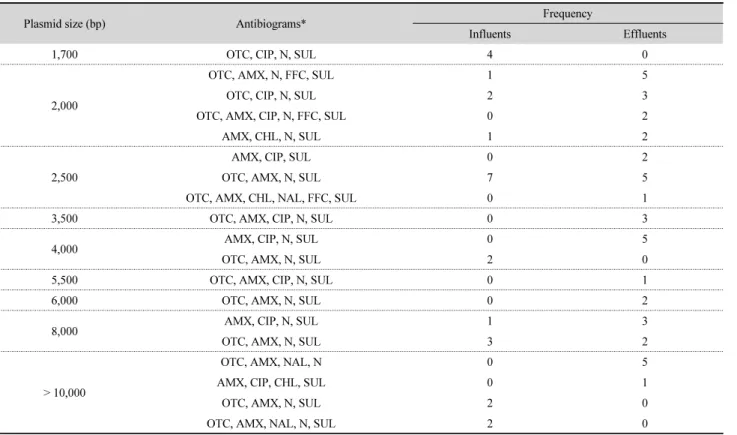
관련 문서
Key words : Aquaculture, Edwardsiella sp., Edwardsiellosis, Fish diseases, Flounder, Streptococcus sp., Streptococcosis, Vibrio sp., Vibriosis... The number
In addition, using the properties of water shown in diverse shapes, Ki Hyung-do's recognition on the reality and future consciousness from regression of
Multi-step method are methods that use information from more than one of the preceding steps,
Current water quality standards for phosphorus in the sewage effluent from 2012 0.2mg/L until tightened according to the existing sewage treatment facilities
A retrospective analysis of invasive tumor percentages of GSS tumor samples showed that samples with high tumor content correlated well with FNAB samples
Biological treatment of wastewater is an economical green technology, however, the effluent water quality of a biological wastewater treatment facility is
So, the result of the comparative analysis in the social studies object, curriculums and textbooks of South and North Korea indicate that the differences
Variations automatic water analyzer & laboratory manual data of suspended solids concentration form effluent of petroleum chemical plant · ·
