The
effects of
monoclonal
antibodies against
iC3breceptors
in mice with
experimentally induced
disseminated
candidiasis
K. H. LEE,M. S. YOON & W. H. CHUN DepartmentofDermatology andInstitutefor Immunologyand ImmunologicalDiseases,
Yonsei UniversityCollege of Medicine, Seoul,Korea
SUMMARY
CR3(iC3b receptor),composed of CD1 lb/CD18, isa
P2
integrin. A protein that shares antigenic and structural homology with the at-chain of CD1 lb/CD 18 has been isolated fromthe surface of Candida albicans. This molecule is thought to be essential in the pathogenesis of disseminated candidiasis. To evaluate the effects of anti-iC3b receptor antibodies on adhesionbetween human dermalmicrovascular endothelial cells(HDMEC) and C. albicans, and in treatment ofcandidal infection, a binding assay of C. albicans to cultured HDMECwas performed in vitro. An anti-iC3b receptor-specific monoclonal antibody was administered to mice infected with C. albicans. Themice were monitored for mortality and renal involvement by culture and histopathological findings. Flow cytometric analysis demonstrated surface expression of iC3b receptor on C.albicans. Theadherenceof C. albicansto HDMECwassignificantly decreased by treatmentwith
anti-iC3b receptor antibodies. Anti-iC3b receptor antibodies significantly increased the survival
time and rate while lowering the renal fungal burden. The iC3b receptors are involved in the adherence of C. albicans to vascular endothelial cells and are likely to be involved in the pathogenesisofdisseminatedcandidiasis. Theincreased survivalin mice infected with C.albicans
after treatmentwith anti-iC3b receptorantibodies indicates that thismodality may bebeneficial forfuture developmentofa new therapy forcandidiasis.
INTRODUCTION
Adherence of a pathogen to host tissue is essential in the
pathogenesis ofinfectious disease and presumablytakesplace
inall forms of candidiasis. Studiesconcerningthemodification
orprevention ofinfection in relation toadhesion mechanisms have been performed in animal and human
models."2
The virulence of a species of Candida can be correlated with itsability to adhereto various cell types, with the mostvirulent C. albicans beingmost adherentto
epithelial
cells. In animal models of invasive disease, the pathogenicities of candidalstrains in vivocorrelate directly with yeast adhesion in vitro.3
Endothelial attachment and subsequentpenetrationare requi-site stages in the induction ofhaematogenously disseminated candidiasis.
Receptors [complement receptor (CR)1, CR2 and CR3]
for C3 degradation fragments (C3b, iC3b, C3dg) are present
Received 14 January 1997; revised 14 May 1997; accepted
14May1997.
Abbreviations: CR, complement receptor; HDMEC, human
dermal microvascular endothelialcell; HUVEC,humanumbilicalvein endothelial cell.
Correspondence: Dr K. H. Lee, Department of Dermatology,
Yonsei University College of Medicine, C.P.O. Box 8044, Seoul,
Korea.
onmanyhuman cellsoperativein theimmuneand
inflamma-toryhostdefenceprocessand may becriticalin the attachment ofinvading micro-organisms.3 C3-bindingstructures are not
specific for mammalian cells ingeneralbut can also befound inmicro-organisms pathogenicfor humans. CR3 is presenton
thesurfaceofmammalianneutrophils, monocytesand
phago-cytes and has been known asCDllb/CD18, Mol,
Mac-i,
oriC3b receptor. It is composed ofan ao-chain of 165000MW and a 95000 MW
P-chain
common to all leucocytes. Thereare iC3b receptors on the surface of C. albicans which share
homology with the a-chain of the
neutrophilic
receptors foriC3b.-6
Westernblottingandimmunoprecipitation havecon-firmed the similar relative molecular weights of CR3 on the human neutrophil and the iC3b receptors isolated from C.
albicans.6.7
Since indisseminated candidiasis the
fungus
isdeliveredtotarget organs through the vascular system, targets for
adher-ence ofblood-borne
fungi
ultimately
include the endothelial cell.Experimentsonadhesion of C. albicanstocultured human umbilical vein endothelial cells(HUVEC)
demonstrate thatincreasing glucoseconcentrations increase adhesion and
mono-clonal antibodies (mAb) recognizing epitopes on neutrophil
CR3 inhibit glucose-enhanced adhesion of C. albicans to
HUVEC.8
However, the exact role of iC3b receptors inhaematogenously disseminated candidiasis in humans hasnot
beenelucidated.
1
1997BlackwellScience LtdWe assessed the role of iC3b receptors on C. albicans in theadhesion of yeast to cultured human dermal microvascular endothelial cells (HDMEC), and the inhibition of this adhesion by mAb against iC3b receptors. In addition, we observed the effects of intravenous injection of anti-iC3b receptor mAb with respect tomortality andlong-terminfection in mice withexperimentally induceddisseminated candidiasis.
MATERIALSANDMETHODS Organisms
One randomly selected pathogenic strain of C. albicans (American Type Culture Collection, Rockville, MD; ATCC 44808) recovered from human blood was used for these experiments. Yeast cells were grown to log-phase in Sabouraud'sbroth, asdeterminedbycolony counts and spec-trophotometric measurement of OD at 420 nm, and were then frozen at -700until use. On theevening before the experiment, aliquots of frozen yeasts were inoculated into RPMI-1640 media (Gibco Laboratories, Grand Island, NY). After attaining log-phase growth, the yeasts were pelleted, washed threetimes with phosphate-buffered saline (PBS) and diluted
spectrophotometrically to anOD of 0 5 at420nm.
Animals
BALB/c mice (Daehan Laboratory Animal Research Centre, Seoul, Korea) weighing around 25 g were used for all experiments.
Monoclonalantibodies
FourmAb directed against epitopes on a subunit of CR3 were Mol (mouse IgM, Coulter Immunology, Hialeah, FL), 44
(mouse IgGl, R&D Systems, Minneapolis, MN), Leu 5 (mouse IgG2a, Becton Dickinson, San Jose, CA), andM1/70. M1/70 (rat IgG2b) was purchased as a hybridoma (ATCC TIB 128) and was purified from supernatant by affinity chromatography on protein G-Sepharose (Hitrap Protein G
column, Pharmacia Biotech, Uppsala, Sweden). The mAb ECCD-2 (anti-E-cadherin, rat IgG2a) was obtained from M. Takeichi (Kyoto University, Kyoto, Japan). ECCD-2 was purified from hybridoma supernatants by protein G-Sepharose affinity chromatography. MEM 48 (mouseIgGI,
R&D Systems, Minneapolis, MN) is a mAb directed against the common ,B-chain ofCR3, leucocyte function associated
antigen-l (LFA-1).
Flowcytometricanalysis
Fifty million colony-forming units (CFU)/ml of C. albicans
werewashed in PBS with 0-5% bovine serum albumin (BSA), counted and aliquotted for staining. Primary antibody at the appropriate dilution was added to each tube. Either an isotype control mAborPBSwithout antibody was used as a first-step control with identical results. The cells were incubated for
30minonice, washed, and second-step antibody, diluted 1/20 in PBSwith 05% BSA, was added. After another 30 min of
incubation, the cells were washed and resuspended in 0-5 ml
PBSwith05% BSA.Propidiumiodide was added immediately before flow cytometric analysis to identify dead cells. Fluorescence was examined by a Becton Dickinson FACStar flow cytometer(Becton Dickinson, MountainView, CA) and
expressed aslogmean channelfluorescence.
Isolation and cultureofHDMEC
HDMEC were isolated from human neonatal foreskins by
trypsinization and Percoll gradient centrifugationasdescribed
previously.9"'0
Briefly, foreskins were cut into small pieces,treated with 0-3% trypsin (Sigma Chemical Co., St Louis,
MD), and 1% ethylenediamine tetraacetic acid (EDTA;Sigma) and individual segments were compressed with a scalpel bladetoexpress microvascular fragments. The microvascular
segments were layered onto a 35% Percoll (Pharmacia AB,
Sweden) gradient in Hank's balanced salt solution (HBSS) and spun at 400g for 15min at room temperature. The fraction with a density less than 1-048 g/mi was applied to
gelatin (Sigma) -precoated tissue-culture dishes and cultured in endothelial basal media (Clonetics Corp., San Diego, CA) withepidermal growth factor 1 ng/ml (Clonetics),
hydrocorti-soneacetate1
gg/ml
(Sigma), dibutyryl cyclicAMP5xI0-5M(Sigma), penicillin 100U/ml, streptomycin 100
gg/ml,
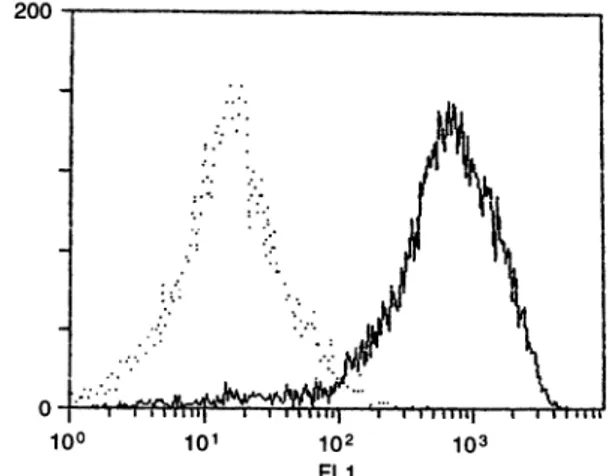
ampho-tericin B 250 jg/ml (Sigma), and 30% human serum (Irvine Scientific, Santa Ana, CA). The resulting cell cultures were free ofcontaminating fibroblasts asassessedby expression ofCD31 (Fig. 1) and uptake of acetylated low-density lipo-proteins. Experiments were conducted with endothelial cells
atpassages2-6.
Candida albicans-HDMEC adherenceassay
HDMEC were plated in gelatin-coated 96-well flat-bottomed culture plates. An isolate of C. albicans was grown in the
RPMI medium, labelledwith [35S]methionine, washed
exten-sively in PBS and resuspended in HBSS with calcium and magnesium, to which was added 1% BSA
(BSA-HBSS+).
Yeast cellswerethen heldat40 topreventgerminationbefore
incubation with HDMEC (5% germ tubes present for C. albicans). Then 4x104 CFU were added to each well of
confluentHDMEC monolayer,and incubated for 1hrat37°.
Forquantification ofnon-specific bindings, a 100-foldexcess ofunlabelledyeastcellswereaddedtolabelledyeastcells.The
supernatantwas removedfromeach well and themonolayers washed twice with BSA-HBSS+ + to remove non-adherent
yeasts.Thesewerecounted inascintillationcounter(Beckman Instruments, Inc., Palo Ato, CA). Adherent yeast cells and HDMEC monolayers were removed with 1% Triton-X
(Sigma), then solubilized in 0-5M NaOH and counted in a
200
100 101 102 103
FL1
Figure1. Flow-cytometric analysis showing the expression ofCD31 onHDMEC.
©1997 BlackwellScience Ltd,Immunology, 92, 104-1 10
P-scintillation
counter. Thepercentage binding of C. albicanswascalculatedas follows:
C.p.m. (adherent yeasts-non-specific binding)
100
C.p.m. (totalyeastsadded-non-specific binding)
Inhibition ofadhesion with mAb
C. albicans was grown in RPMI medium and was then
preincubated with 100pl of mAb to ac-subunit ofCR3 for 45min. 100g1 of20
gg/ml
ofmAbwereused. Afterpreincu-bation withantibodies, the adherence assaywasperformedas
described above in the continuous presenceof antibodies. All
the individual experiments were performed three times
independently. Experimental infection
Mice were divided into three experimental treatment groups for the assessment of anti-iC3b receptor antibody on the
resulting prevention of systemic candidiasis. The threegroups were: Group I, controls, received the vehicle for the mAb
(0-5% human serum albumin) and was inoculated with one dose of 106 CFU of C. albicans grown in RPMI-1640 per mouseintothe lateral tailveins ofagroupof 45normal mice;
Group II,mAb Ml/70, beginning before infection received
mAb M1/70 2mg/kg infused intravenously once per day for 5days (day0today 5). Group III, isotype control, beginning
before infection received mAb ECCD-2 2mg/kginfused
intra-venously once perday for 7 days (day 0today 5);The fourth groupreceived only equivalentamountsofmAbM1/70atthe same time. All animals were monitored for 30 days, and
mortality was recorded. Two groups of 20 mice each were
studied, an only C. albicans-inoculated group and a group
treated anti-M1/70mAb and inoculated with C. albicans as
previously described. On days 0, 2, 4 and 6, five mice per experimental group were selected at random and killed by
cervical dislocation. Thekidneysof each of these animalswere
removed aseptically,transferred into sterilepolyethylene bags,
and homogenized in 5ml of sterile saline. Samples were removed from eachhomogenate, seriallydilutedin0-9%NaCl, platedontoSabouraud dextroseagarplates,and incubatedat
250 for 16 hr. The colonies were then enumerated, and CFU werecalculatedpergramof eachorgan. Allexperimentswere
performed in triplicate. To document the administration of
M1/70mAb sufficientto produce antibodyexcess, samples of serum were analysed for the presence of residual M1/70 antibody. Bloodwascollected fromtheorbital,venousplexus. This wasassessedby indirect immunofluorescence analysis in
which C. albicanswere incubated in serumfor 30minat250,
andthen,afterwashing, incubatedin fluorescein isothiocyan-ate (FITC)-conjugated goat anti-rat immunoglobulin for an additional 30minat250. Antibody bindingto C. albicanswas
quantified byflowcytometry. Formalin-fixedportions of
kid-neys obtained at2, 4 and 6 days were stained with Periodic
acid-Schiff reagent(PAS) to searchfor fungi inkidneys.
Statisticalanalysis
Alldataareexpressedas mean+standarderror.Alldifferences in survival wereanalysed bythe Wilcoxon rank sumtest and Kruskal-Wallistest. Product limit methodwasusedtoanalyse
survival data andrepeated ANOVA wereappliedtocompare
themeanCFUpermeankidney weight.Statisticalsignificance wasdefined as aP-value of <0 05.
RESULTS Expression of iC3breceptoronC.albicans
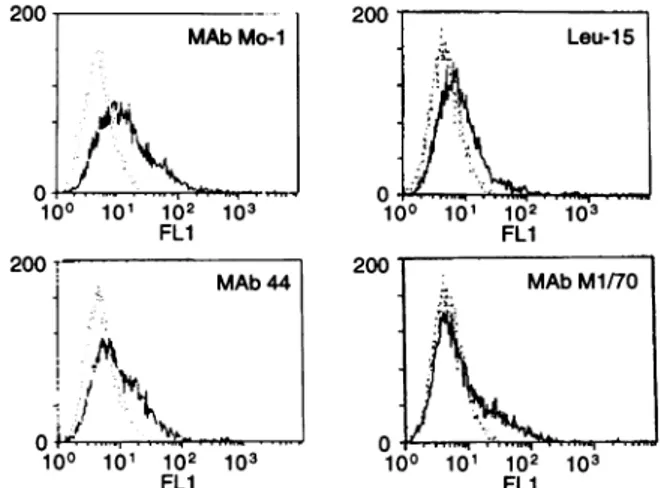
The iC3b receptor expression was observed by
immunofluo-rescenceflowcytometryfromC.albicanspretreatedwith
anti-iC3b receptor antibody purified from culture supernatant of ATCC MI/70 antibody by affinitychromatography (Fig.2).
Effect of anti-iC3breceptorantibodyonadhesion of C. albicans
toHDMEC
Preliminarystudiesweredonetocomparetherateof adhesion
between HDMEC and C. albicansaccordingtothe number of C. albicans and elapsed time. The binding of 5x105 CFU,
1 x 106CFU and 2x106CFUof C. albicanstoHDMECwere
25-1 +3-8%, 29-9+222% and 283 +1-5%, respectively. The
adherences 30min, 1 hrand 2 hrafterculturewere8 1+19%,
241+37% and 25-4+4-9%. Theadherence increased signifi-cantly after 1 hr of culture. Furtherstudies were carried out
under culture conditions with 106 CFU of C. albicans being
cultured for 1hr.
The adhesion of C. albicans without any prior treatment to HDMEC was 240+336%. Adherences were 25-4+554%,
27 9+1 8%, 26-3+0-3% and 16 5+2-2%whenC. albicanswas treated before inoculation with rat IgG2b,mAb ECCD-2,
anti-CD18,andM1/70 mAb, respectively (Fig. 3).
Effect ofanti-iC3breceptorantibodyonmortalityin micewith experimentallyinduceddisseminatedcandidiasis
Preliminarystudieswereperformedtodeterminethemortality
ratesofBALB/cmiceaccordingtothe numberof C. albicans.
All miceinjected with 5x106CFU of C. albicansculturedin
RPMI-1640 mediaexpired within 1 day.At 1 x106 CFUand
5x105 CFU, the average survival durations were 27+0-6
days and 43+0-6 days, respectively. Inoculation was done with 1 x106 CFU of C. albicans to determine mortality and
effects of the anti-iC3b receptor antibodies. Group I was inoculated withonlyC. albicans. GroupIIwaspretreatedwith
200 2C MAb Mo-I .., I..: 10° 101 102 103 FLi Leu-15 )0 101 102 103 FLI MAbM1/7O 100 101 102 103 FL1
Figure2. Expressionof iC3b receptoronthe surface of C. albicans. Candida albicans was incubated with anti-iC3b receptor mAb anti-Mol, Leu5,44andMI/70andanalysed byfluorescence-activated cell sorter. The expression of iC3b receptors can be observed on the surface of C. albicans.
C 1997Blackwell ScienceLtd, Immunology, 92,104-110
0 14" o-T- "4 100 101 102 103 FLi )o --0 MAb44 .
!"IXPM
"...
V /v 4 M AD. t%-bEffects
No Ab Irrelevant mAb Anti-CD18 Anti-iC3bR -d -I 0 10 20 %binding 30Figure 3. The effects ofanti-iC3breceptorantibodiesonthe adhesion
of C. albicans to HDMEC. Candida albicans was pretreated with irrelevant antibody ECCD-2, mAbanti-CD18 against the 1-chain of iC3breceptor,andM1/70against the a-chain of iC3breceptor,added inconcentrations of 20jg/ml at370 for 45 min, and incubated with HDMEC monolayer at 37° for 1hr. Results show the mean percentage+SD ofquadruplicate microtitre wells.
Ml/70mAb 1 hr prior to inoculation with C. albicans and
Group III was pretreated with mAb ECCD-2 1hr prior to
inoculation with C. albicansas control. Immunofluorescence studies revealed strong fluorescence on the surface of C. albicans collected from the serum of mice 6 days after the initialinoculation of M1/70 mAb. Fluorescence wasobserved on the surface of C. albicans by immunofluorescent flow
cytometry from serum collected 2, 4 and 6 days after inoculation.
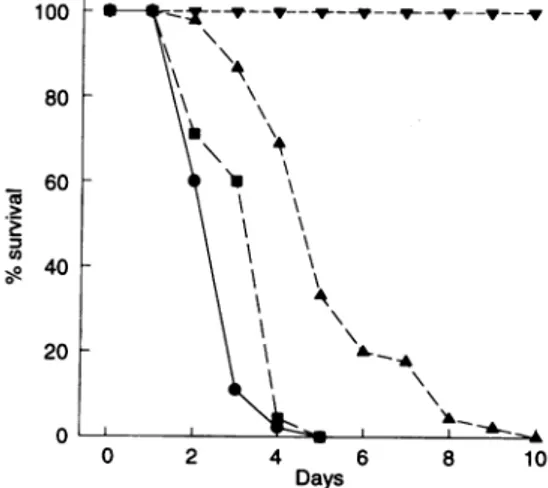
GroupIVwasinjectedwithonly anti-iC3b antibodiesand all 45 micewerealive 30days afterinjection. However, all 45 mice in bothGroupI andGroupIII expired between 2 and5
100 80 - 60 ._,-40 20 0
Table1. Effectof anti-iC3breceptorantibodyonduration of survival
inmice withexperimentally induced disseminated candidiasis
No.of Survival duration
Inoculationgroup animals (days)
C. albicansonly* 45 2-7+0 7
C. albicans+mAb MI/70t 45 5-3+1 8
C. albicans+control
mAbJ
45 34+009mAbM1/70only§ 45 >30
*GroupIwasinoculatedwithonly C.albicans.
tGroup II was inoculated with C. albicans and intravenously
injected with anti-iC3b receptor antibody M1/70 at 2 mg/kg of
concentration.
tGroup III was intravenously injected with mAb ECCD-2 at 2mg/kg of concentration and inoculated withC. albicans.
§Group IV was injected with only anti-iC3b receptor antibody
M1/70.
days after inoculation. All mice in Group IIexpired between 2 and 10 days after inoculation. However,the mortality rate inGroupII at 4days after inoculationwas 311%,which was
significantly lower than the mortality rates at this time in
Groups I and III (P=00001, Fig. 4). The average survival durationsforGroupsI, IIandIIIwere2 7 + 0 7days, 5 3+18 days,and 34+09 days, respectively, (Table 1). These differ-encesweresignificant.White blood cell counts andmorphology
of mouseneutrophilswere notdifferent between the irrelevant
mAb-treated group and theM1/70-treatedgroup.
Effect of anti-iC3b receptorantibodyonrenalinvasion of C. albicans in mice withexperimentally induceddisseminated
candidiasis
To evaluate the effects of anti-iC3b receptor antibody in disseminatedcandidiasis,micewereinjectedintravenouslywith 5 x105 CFU ofC. albicanscultured in RPMI-1640. Themice
were killedatrandomand the kidneyswere removed,
homo-genized and cultured on a Sabouraud dextrose agar plate. 10r 0) a) 'D C) J 0 E 0) cl 0) 0 -j 0 2 4 Days 8 10 9 Group \ GroupII 8 . 7 6 5 2 4 Days post-infection 6
Figure 4. Effect of anti-iC3breceptor antibodyon mortality of mice
with experimentally induced disseminated candidiasis. Group Iwas
inoculated with only C. albicans (0). Group IIwasinoculated with C. albicansandintravenouslyinjected with anti-iC3breceptorantibody M1/70 at 2 mg/kg (A). Group III wasintravenously injected with mAb ECCD-2at2mg/kg and inoculated withC.albicans(R).Group IV was injected with only anti-iC3b receptor antibody M1/70 (V). Themortalityandsurvivalduration of the fourgroups werecompared.
Figure5. Effect ofanti-iC3breceptorantibody onrenal invasion of
mice with experimentally induced disseminated candidiasis. GroupI
wasinoculated withonly C. albicans. Group IIwasinoculated with
C. albicansandintravenously injectedwithanti-iC3breceptorantibody M1/70at2mg/kg. Renal specimenswereharvestedevery2days for 6 days to evaluate the invasion of kidneys. A quantitative culture assaywasperformed three times foreachmouse. TheaverageCFU of C. albicans/gofkidneywerecompared.
(a)
(b]..
;7
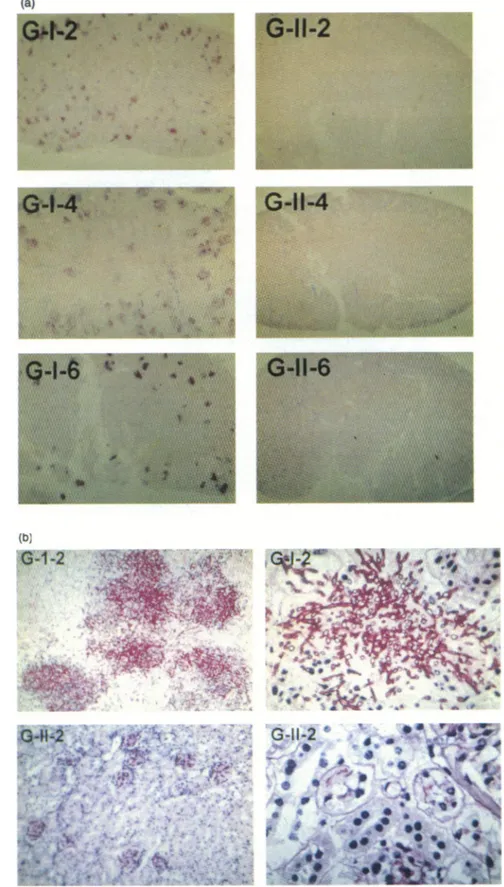
_4 l_s X. ~_ _ .Figure6. (a)Histopathological findings ofkidney of mice with
expernmentally
induceddisseminated candidiasis and treated withanti-iC~b receptor antibody (PAS stainx40). (b) Higher magnification (x 400). Group Iwasinoculated with only C.albicans.
Group IIwasinoculated withC. albicans andintravenouslyinjectedwithanti-iC~breceptorantibody
Ml/70
at2mg/kg. Renalspecimens were harvested every2 days for 6 days toIevaluate the invasion ofkidneys. Specimens were paraffin-embedded and
stained withperiodicSchiffreagent. Thisstainingrevealedmanyfungalsporesandhyphaeintheglomerulusatthe seconddayof infection inGroupI,whereasGroupIIshowedonlyfewfungalsporesandhyphae. G-1-2, Group I-day 2;G-I14,Group I-day 4;
G-I-6, GroupI-day 6;G-1I-2,Group II-day 2; G-11-4, Group
Il-day
4; G-II-6, GroupIl-day
6.in disseminated
On the days 2, 4 and 6 of infection, Group I had
3 0+0-2x 109CFU, 3-3+1-8x109CFU, 32+O1 x107CFU
of C. albicans, respectively, in 1g ofkidney. By comparison, Group II had [*6+1 3x106 CFU, 28+l1-2x 105 CFU,
1-9+1-2x 105 CFU of C. albicans, respectively, in 1 g of
kidney. Thesedifferencesinrenalinvolvementwerestatistically
significant (P=0-0001, Fig. 5).
Periodic acid-Schiff staining of the extracted renal
speci-mens revealed many fungal spores and hyphae in the renal
medulla on day 2 ofinfection inGroup I. Group II showed
onlya fewfungal spores and hyphae on day 2 of infection.
GroupIshowedheavy infiltrationofinflammatorycellsatthe sixthdayofinfection(Fig. 6).
DISCUSSION
Theimportance of the adherence of Candidain the
pathogen-esis ofdisseminated disease is increasingly recognized. There
have beenreportsthatpathogenicityofcandidal strains in vivo
correlate directly with yeast adhesion in vitro.' However,
studiesonfungus-to-hostcell adhesionmechanisms have been
relatively sparse. It is possible to categorize these adhesion
mechanisms by considering the components ofeach
ligand-receptor.'1 The first category includes protein-protein inter-actions such asthe iC3b receptor on C. albicans. The second
category includes interactions in which a surface Candida
proteinwithlectinactivity recognizessugarresiduesassociated
with the host-cell plasma membrane. The third category
involves acarbohydrate ligand, mannan, onthe Candida cell surface that is recognizedby an unknown host-cell receptor.
Complement fragments adheremore readily to Candida than
toendothelial cells in vitro.4'2CR3onneutrophilsis involved inrecognition of iC3bonthesurface ofmicro-organisms and
subsequent phagocytosis. TheiC3breceptoronthesurfaceof C. albicans hasrecentlybeen isolated andcharacterized.'3The
iC3b receptor in Candida shares antigenic and structural homologywith CR3 onneutrophils.46
As we, and others, have observed the expression of iC3b
receptoris usually below 50%dependingon thetypeof mAb used, condition of culture medium, species or strains of
Candidaused, etc.56'8"12The low expression in ourstudy was
due to the RPMI medium with low glucose concentration.
When theglucoseconcentrationwasincreased, the expression of iC3b receptor was up-regulated (unpublished results).
However, the differenceinthe expression could be attributed
to thetypeofmAbused.
The relationship between hyperglycaemia and candidal infection has been well documented.14 However, the exact mechanism by which sensitivity to candidal infection is
increased isunknown.There havebeenreportsthat expression ofiC3breceptoronCandida andadhesiontoculturedHUVEC correlate with glucose concentrations.56 Moreover, adhesion to cultured HUVEC is suppressed by 50-80% with mAb directedagainst theac-chain of the iC3breceptor.8
As there are marked differences between the endothelial cells ofvarious
vessels,9"10"15
andasC. albicans infectsthe skin andmucousmembranes easily,weperformedadhesion experi-mentsusing culturedHDMEC isolated from skin. Ourresults show that adherence of C. albicans correlates with reactiontime and number of organisms. But there was a marked
decrease when anti-iC3b receptor mAb were added. These
findingsaresimilar toprevious studies usingHUVEC.8
Intercellular adhesionmolecule-l (ICAM-1),whichactsas
a ligand for neutrophil CR3, has no role in the adhesion of iC3breceptoron C. albicans.8"6 Recently, the host ligandfor
the iC3b receptor on C. albicans has been characterized as
iC3b."' C3 is synthesized from endothelial cells'8 and it is
plausibletoassumethatC3 isdegraded byproteolyticenzymes
in the serum orcell with the resultant iC3battaching to the endothelial surface and acting as a ligand for the iC3b
reetr7,18,19
receptor.7""
In the present study, binding of C. albicans to HDMEC
was partially inhibited by mAb M1/70. This would be
explainedbythefindingof 50000, 100 000, 130 000 and 165 000 MWproteins inextractsofC. albicansreacting with anti-iC3b
receptor antibodies in immunoblotting and
immunoprecipit-ation studies.6'7 Hence, thecombination of thisvariability of the iC3b receptorepitopesand specificity of each mAbmight
have resulted in the partial inhibition as seeninour study. We confirmed that the iC3b receptor on C. albicans is essential for adhesiontoculturedHDMEC. Thusweassumed that prevention of the yeast-to-host cell adhesion could block thedissemination of Candidato surrounding tissue. haemato-genously disseminated candidiasis was induced in mice by intravenous injection with C. albicans. The mortality rate of the group injected with anti-iC3b receptor mAbwas signifi-cantly lower than that of the normal control. The 50% survival
rateincreasedsignificantly. Theamountofyeastand the extent
of invasion of renal tissue was significantly less. The affinity
constantforthe iC3breceptoris 2-1+0 5x106L/M for human neutrophils20 and virtually identical to that of C. albicans.5 However, Candida possess 2x106 iC3b receptors per yeast
which is nearly 100 times higher than neutrophils.5 The increased survivalrateinmicetreated withanti-iC3breceptor
antibodies indicatesthat thismodality mightbe beneficialfor future development of a new therapy for disseminated
candidiasis.
TheiC3breceptor on Candida ishomologous in srtucture to human CR3, however, it is not the same molecule. There isno doubt that antibodies directly against the Candida iC3b
receptorwouldbelikelytobindbetterto Candida andtohave betterinhibitory effects. However, at this time, wecould not
obtain the purifiedcandidal iC3b receptor anditsmAb. It is desirable that similar experiments using the above antibody
becarriedoutasanextstep toevaluatethe clinical usefulness
ofanti-iC3breceptorantibodyinthetreatmentof haematogen-ouscandidiasis.
The anti-iC3b receptormAb may have deleterious effects
on host neutrophils and aggravate the course ofdisease, but
ourin vivo results indicate that theyhave atherapeutic effect
in haematogenously disseminated candidiasis without signifi-cant alterations in host neutrophils. Attachment to the iC3b
receptor on C. albicans blocks adhesion to endothelium and subsequent systemic invasion. Adherence to the endothelium iscorrelatedwithvirulence. However, it istooearlytodiscuss
theinteractions betweenintravenously injected C. albicans and
the anti-iC3b receptor mAb based solely on the current experiment. We plan to use radioisotope-labelled anti-iC3b antibodiesforfuturestudies andtoevaluatethesafety profiles
ofanti-iC3b mAb inhumans.
ACKNOWLEDGMENTS
Theauthorsthank Dr Masatoshi Takeichi (Kyoto University, Kyoto,
Japan) for providing ECCD-2 mAb. We are also grateful for the
experttechnicalassistanceof Nam Soo Jang. This paper was supported
in part by the Non-Directed Research Fund, Korea Research
Foundation., Korea.
REFERENCES
1. BEACHY E.H. (1981)Bacterial adherence:adhesion-receptor
inter-actions mediatingtheattachment ofbacteria tomucosal surfaces.
JInfectDis143, 325.
2. FRETER R. & JoNES G.W. (1983)Models forstudyingthe role of
bacterialattachment invirulence andpathogenesis.RevInfectDis 5,s647.
3. SCHELD W.M., STRUNK R.W., BALIAN G. & CALDERONE R.A.
(1985) Microbial adhesiontofibronectin in vitrocorrelates with
production of endocarditis in rabbits. Proc Soc Exp Biol Med
180,474.
4. EDWARDS J.E. JR., GAITHER T.A., O'SHEA J.J. et al. (1986)
Expression of specific binding sites on Candida with functional andantigeniccharacteristics ofhuman complementreceptors. J
Immunol137, 3577.
5. GILMORE B.J., RETINAS E.M., LORENZ J.S. & HosTErrER M.K.
(1988)AniC3breceptor onCandida albicans:structure,function, andcorrelates forpathogenicity.JInfectDis157, 38.
6. HosTmrrER M.K., LORENZ J., PREUs L.& KENDRICKK.E.(1990) The iC3b receptor on Candida albicans: subcellular localization and modulation ofreceptor expression by glucose. JInfectDis 161, 761.
7. EIGENTLER A., SCHULZ T.F., LARCHER C. et al. (1989)
C3bi-binding protein on Candida albicans: temperature-dependent expression andrelationshiptohuman complementreceptor type
3.InfectImmun57, 616.
8. GUSTAFSONK.S.,VERCELLOTr'iG.M., BENDELC.M.&HosTETER M.K. (1991)Molecularmimicry in Candidaalbicans: role ofan
integrin analogue in adhesion of theyeasttohuman endothelium.
JClin Invest87,1896.
9. LEEK.H.,LAWLEYT.J.,Xu Y.&SWERLICK R.A.(1992)VCAM-1,
ELAM-1, andICAM-1-independent adhesionof melanoma cells toculturedhumandermal microvascular endothelial cells.J Invest Dermatol 98, 79.
10. SWERLICKR.A., LEE K.H., LiS.J., CAUGHMANNS.W. & LAWLEY T.J. (1992) Regulation ofvascular cell adhesion molecule 1 on humandermalmicrovascularendothelial cells. J Immunol 149, 698. 11. CALDERONE R.A. &BRAUN P.C.(1991) Adherence and receptor
relationships ofCandida albicans. Microbiol Rev 55, 1.
12. HEIDENREICH F. & DIERICH M.P. (1985) Candida albicans and
Candida stellatoidea in contrast to other Candida species, bind iC3b and C3d but not C3b. Infect Immun 50, 598.
13. ALAEI S., LARCHER C., EBENBICHLER C., PRODINGER W.M., JANATOVA J. & DIERICH M.P. (1993) Isolation and biochemical
characterization of the iC3b receptorof Candida albicans. Infect Immun61,1395.
14. HORN R., WONG B., KIEHN T.E. & ArMSTRONG D. (1985) Fungemia ina cancerhospital: changing frequency,earlier onset
andresultsoftherapy. RevInfectDis7,646.
15. SWERLICK R.A LEE K.H., WICK T.M & LAWLEY T.J. (1992)
Humandermalmicrovascularendothelial butnothumanumbilical vein endothelial cellsexpress CD36 in vivo and vitro. JImmunol
148, 78.
16. LoS.K.,VANSEVENTERG.A.,LEVINS.M. & WRIGHT S.D. (1989) Two leukocyte receptors (CD1 la/CD18 and CD1 lb/CD18) mediate transient adhesiontoendothelium by bindingtodifferent ligands.JImmunol143, 3325.
17. BENDELC.M. & HosTETTER M.K. (1993) Distinct mechanisms of epithelial adhesion for Candida albicansand Candida tropicalis.
JClinInvest92, 1840.
18. BROOIMANS R.A., VAN DERARK A.A., BUURMAN W.A.,VAN Es
L.A.& DAHA M.R.(1990)Differential regulation of complement factorHandC3productioninhuman umbilicalveinendothelial cellsby IFN-,y andIL-1. JImmunol144, 3835.
19. WRIGHT S.D., REDDY P.A., JONG M.C.T. & ERICKSON B.W.
(1987)C3bireceptor (complement receptor type3) recognizesa
region ofcomplementprotein C3 containing the sequence Arg-Gly-Asp.ProcNatl Acad Sci USA84,1965.
20. GORDON D.L., JOHNSON G.M. & HOSTETTER M.K. (1987) Characteristics ofiC3b binding to human polymorphonuclear leukocytes.Immunology 60, 553.