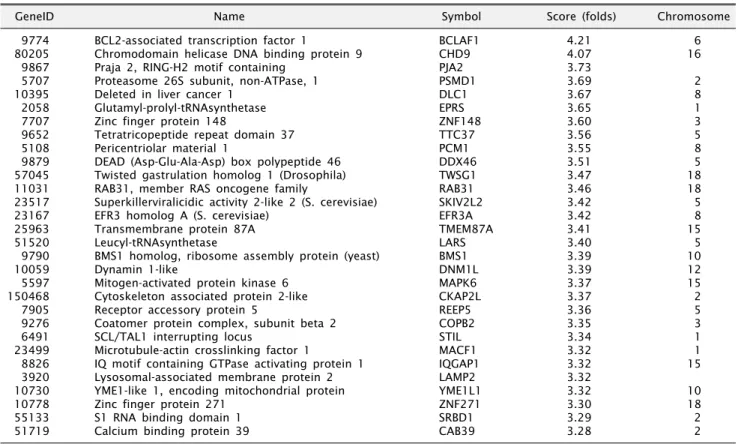
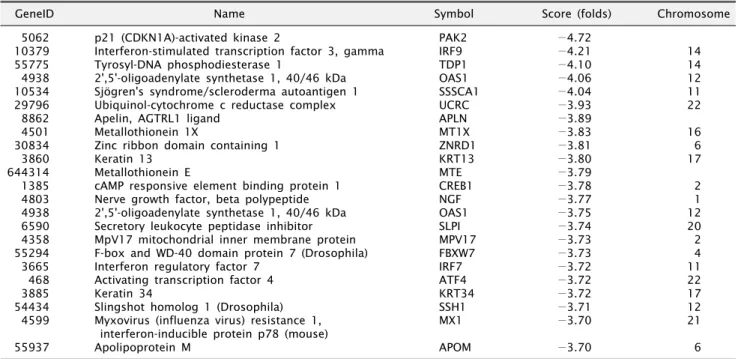
관련 문서
In vitro cell migration in APE or JAG1 siRNA-treated M059K cell line Fig,11 Down-regulation of JAG1 induces S phase arrest in
:Acidulated phosphate fluoride treatment and formation of caries-like lesions in enamel: effect of application time.. Schmidt HF : The fluoride varnish
In contrast, cells expressing wild-type form or constitutively active form of RapC (GFP-RapC and GFP-RapC G13V ) showed decreased cell area and cell adhesion, whereas rapC
Biochemical markers of bone turnover can be classified according to the process that underlie in markers of bone formation, products of the osteoblast
(1) PCR monitoring of bphC gene of Ralstonia eutropha H850 in soil Amount of bphC gene amplification of H850 cells was not generally proportional to the cell density
4 > Effects of Taro on COX-2 expression and iNOS expression(hot water) in human thyroid cancer cells. The cells were pretreated for 48hours with either
of mineralized cancellous bone allograft(Puros) and anorganic bovine bone matrix(Bio-oss) for sinus augmentation: Histomorphometry at 26 to 32 weeks after
Comparison of cell attachment on the bone grafting materials synthesized as the defined mixing ratio of Hydroxyapatite (HA) and β-tricalcium phosphate (β-TCP) by DAPI staining