—110—
서 론
대구(Gadus macrocephalus)는 겨울에 비교적 수심이 얕 은 연안으로 무리를 지어 이동하여 산란을 하는 것으로 알 려져 있다(Mishima, 1984; Hattori et al., 1993; Shimada and
Kimura, 1994). 태평양 대구는 번식군에 따라 회유범위가
매우 한정되어 있고 산란장으로의 회귀 성향이 강하여 각 번식군의 독립성이 매우 강한 것으로 알려져 있다. 진해만 대구도 mtDNA와 msDNA 분석결과에 근거하여 하나의 독 립된 집단을 형성하는 것으로 추정하였다(Gwak and Naka-yama, 2011). 이와 같이 산란장 회귀 성향을 갖고 있는 해
산어류에는 청어(Clupea pallasii) (Hoerston, 1982; Schwarzet al., 1993; Flostrand et al., 2009), European Plaice (Pleu-
ronectes platessa) (Hunter et al., 2003) 그리고 대서양 대구 (Gadus morhua) (Robichaud and Rose, 2001; Windle and Rose, 2005)가 알려져 있다. 특히, 청어와 대구의 경우 이와
같은 산란장 회귀 특성을 이용하여 산란장 별로 자원관리 를 하고 있으며, 산란장의 환경특성을 구명하는 것은 수산 자원의 효율적 관리를 위해서 매우 중요하다고 할 수 있다.캄차카 동부 연안에서는 대구 난자치어와 수온의 지리적 분포로부터 산란장과 산란 적수온을 추정하였다(Miyazaki,
1942). Uchida (1936)는 진해만에서 포란한 대구 친어가 다
수 어획되었고 산란기 이후 자치어가 채집되므로 진해만이 대구 산란장일 것으로 추정하였고, 진해만 대구는 자치어의 생존율을 높이고 저수온기에 빠른 성장을 위해 수온이 상 대적으로 높은 남쪽의 진해만을 산란장으로 택했을 것으로 추측하였다. 이와 같이 진해만에 산란회유한 대구 친어와 치어 채집에 대한 보고는 있으나 진해만 대구의 산란기 및 산란장 수온 그리고 초기 성장에 대해서는 구체적으로 밝진해만 대구의 부화일 및 초기 성장에 관한 연구
곽우석∙최병언*∙이소광1
국립경상대학교 해양과학대학 해양산업연구소, 1경상남도수산자원연구소
Spawning Time and Early Growth of Pacific Cod (Gadus macrocephalus) in Jinhae Bay, Korea
by Woo- Seok Gwak, Byung-Eon Choi* and So-Gwang Lee1(College of Marine Science, The Institute of Marine Industry, Gyeongsang National University, Tongyeong 650-160, Korea; 1Gyeongsangnam-do Fisheries Resources Research Institute, Tongyeong 650-974, Korea)ABSTRACT Hatching date and early growth of Pacific cod Gadus macrocephalus were estimated by examination of otolith microstructure of Pacific cod juveniles collected in Jinhae Bay of Korea from May to June, 2008. Water temperature during the main spawning time ranged from 7 to 9�
�C according to the geographic distribution of temperature measured from November to May between 2006 and 2009. The spawners were collected from December to February between 2006 and 2008, and the gonadosomatic index of spawners was larger in females than in males, showing a peak in January.
Total length of juveniles ranged from 37.5 to 94.9 mm (63.2±
±11.0 mm; mean±±SD). The number ofdaily growth increments in juvenile otoliths were on average 102
±±15 in May 17 samples, 119±±16 inMay 29 ones and 116±
±18 in June 3 ones. Hatching dates estimated from the number of daily growthincrements ranged from late December to mid-March, showing the peak between late January and mid-February. Daily growth in total length (TL, mm) can be adjusted to the Gompertz curve: TL
t==123.2 exp {
-- exp [-- 0.0165 (t-- 81.8)]} (r2==0.93, n==273).Key words : Pacific cod, juvenile, otolith, growth in length, spawning period
*교신저자: 최병언 Tel: 82-55-772-9150, Fax: 82-55-772-9049, E-mail: [email protected]
http://www.fishkorea.or.kr
혀지지 못하였다. 어류의 부화일과 초기 성장 추정에는 이 석의 미세구조가 널리 이용되는데 (Yoklavich and Bailey,
1990; Quinonez-Velazquez et al., 2000), 어류 이석의 미세성
장선을 관찰 분석하면 각 개체의 일일성장률, 산란시기, 회 유시기 등을 추정할 수 있다(Stevenson and Campana, 1992).또한 어류의 부화시기 추정을 위해서는 이석 일륜의 최초 형성시기와 미세성장선의 주기성이 먼저 증명되어야 한다.
태평양 대구 치어의 경우 부화일부터 일륜이 형성된다고 밝혀져(Narimatsu et al., 2007), 이 결과를 진해만 대구 치어 에 적용하면 부화시기를 추정할 수 있을 것으로 생각된다.
산란장 회귀 본능이 강한 대구의 효율적 자원 관리 및 보 존을 위하여 정확한 산란장과 산란시기 환경조건을 파악하 는 것이 중요하다. 본 연구에서는 진해만 대구의 산란기간 으로 추정되는 기간 동안 생식선숙도지수(GSI)를 조사하고,
5월과 6월에 진해만에서 채집된 대구치어의 이석 분석을
통해 산란시기 및 초기 성장을 추정하였다. 또한 3년간의 산란기 및 산란기 이후의 진해만 수온 조사 결과를 통해 수 온과 진해만 대구 산란의 연관성에 대하여 고찰하였다.재료 및 방법
1. 산란장 수온 및 대구 성어GSI 조사
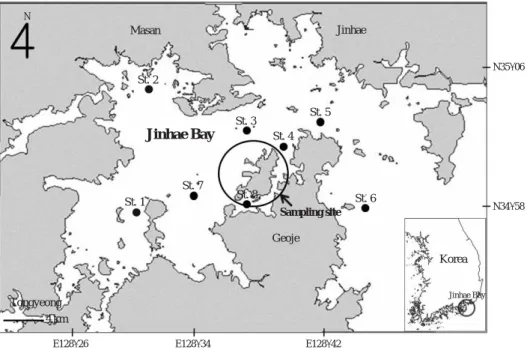
수온은 경상대학교 실습선 참바다호를 이용하여 2006년 부터 2009년까지 대구 산란회유가 시작되는 11월부터 그 다음해 5월까지 CTD (SBE 19-03)를 이용하여 매월 1회 8
개의 정점에서 측정하였다(Fig. 1). 산란기간 추정을 위한 생식소숙도지수(GSI)조사는 2006~2008년 2년간 산란회 유기간인 12월~2월 동안 매월 경상남도 진해만 해역에서 호망에 의해 어획된 것을 거제시 장목면 외포리 거제수협 위판장에서 구입하였다. 시료채집은 Uchida (1936)가 보고 한 진해만 대구 산란기를 초기, 중기, 말기로 나누었을 때 각 시기별 채집된 어체 크기가 다르다는 것에 근거하여 산 란기인 12~2월에 매월 1회씩 2년에 걸쳐 각 시기별 30개 체를 채집하여 실험실로 운반한 후 체장(0.1 cm), 체중(0.1
g)을 측정하였다. 또한 생식소중량지수를 구하기 위하여 생
식소 중량(0.1 g)을 계측하였다. GSI의 식은 다음과 같다:GSI=
=GW/BW×100여기서, GW는 생식소 중량, BW는 체중이다.
2. 이석분석
대구 치어는 2008년 5월 17일, 29일, 6월 3일에 각각 297 개체, 94개체, 299개체(총 690개체)를 경상남도 거제시 진 해만 내의 괭이섬과 칠천도 부근에서 정치망을 이용하여 3 회에 걸쳐 채집하였다(Fig. 1).
채집된 시료는 냉장 보관하여 실험실로 운반하였다. 실험 실에서 각 개체의 전장(0.01 mm)과 체중(0.01 g)을 측정하 였으며, 연령사정을 위하여 이석을 추출하였다. 두개골을 절 개하여 이석을 추출한 뒤, 증류수를 이용하여 이물질을 제 거하였다. 분석시료의 통일성을 기하기 위해 두개골 우측에 있는 이석(편평석)을 연령사정에 이용하였다. 추출한 이석
Fig. 1. Map showing the stations for water temperature measurement (●) and collection site (large open circle) of juvenile Pacific cod, Gadus macrocephalus in Jinhae Bay.
N35�06′
N34�58′
E128�26′ E128�34′ E128�42′
Jinhae Bay
Geoje
Korea
Jinhae Bay Sampling site
St. 1 St. 2
St. 3 St. 4
St. 5
St. 6 St. 7
St. 8
4 km
Jinhae Masan
N
Tongyeong
은 폴리에스테르 수지(Polycoat)와 촉진제 그리고 경화제 를 이용하여 상온에서 72시간 동안 고정하였다. 인공수지에 고정된 이석은 사포(Grid 80, 400, 800, 1,200, 2,000)와 Lap-
ping film (Grid 4000, 6000, 10,000, 15,000)을 이용하여 0.2~0.3 mm 두께로 연마하였다. 연마 과정 중 수시로 관찰
하며, 미세성장선이 잘 관찰될 때까지 연마하였다. 연마한 이석은 슬라이드글라스에 고정한 뒤, 현미경으로 관찰하면 서 재연마를 하였다. 이석 관찰은 입체현미경(OLYMPUSTOKYO, BX51)과 컴퓨터의 영상분석 시스템(AnalySIS LS)
을 이용하였다. 일륜은 영상분석프로그램을 이용하여 150~200% 확대하여 판독을 하였으며, 이석은 한 평면상으로 자
라지 않기 때문에 먼저 가장자리 성장선을 계수하고 연마 를 계속하여 핵까지 계수하였다. 분석을 통해서 일륜의 판 독이 어려울 때 재연마를 한 뒤 다시 관찰하였고 이번 연 구에는 이석 일륜 관찰이 가능한 273개를 이용하였다. 부화 일과 전장 사이의 관계는 Gompertz 성장모델로 비선형회귀 분석에 의하여 계산(Microsoft Excel solver routine) 하였다.결 과
1. 산란장 수온조사
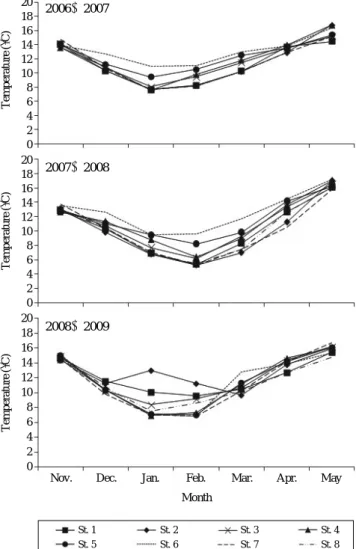
2006~2009년 11월부터 5월까지의 정점별 수온은 일반
적으로 1, 2월에 가장 낮았다(Fig. 2). 전체 정점에서 2006~2009년 저층의 평균수온은 1월 8.2� C, 2월 8.0� C로 나타났
으며, 2006~2008년에는 정점 5, 6과 2008~2009년에는 정 점 1, 2가 비교적 수온이 높았다. 조사결과 1, 2월이 대구 난 부화 적수온으로 나타났으며, 수온만으로 살펴보면 2006~2008년 정점 5, 6과 2008~2009년 정점 1, 2를 제외한 6개
의 정점이 산란하기에 가장 적당한 장소로 판단된다. 그리 고 비교적 높은 수온을 나타낸 정점은 산란 장소로 적합하 지 않은 것으로 생각된다. 2006~2009년 11월부터 3월까지 정점에서의 표층과 저층수온은 큰 차이가 없었지만 4월과5월의 표층과 저층수온은 차이가 있는 것으로 나타났다. 또
한 정점별 저층의 수온은 비슷한 수치를 기록하였다.2. 성어GSI 조사
대구 성어 GSI 조사를 위해 채집된 시료의 전장과 체중 은 Table 1과 같았다. 대구 암컷과 수컷 모두 산란기 후반 으로 갈수록 전장과 체중 모두 감소하였다. 생식소숙도지 수(GSI)는 암컷은 1월 평균 값이 28.3으로 가장 높았고 2
Fig. 2. Monthly variation of water temperature in Jinhae Bay from November to May, 2006~2009.
20 18 16 14 12 10 8 6 4 2 0 20 18 16 14 12 10 8 6 4 2 0
2006~2007
2007~2008
2008~2009 20
18 16 14 12 10 8 6 4 2 0
Nov. Dec. Jan. Feb. Mar. Apr. May
Month
St. 1 St. 2 St. 3 St. 4
St. 5 St. 6 St. 7 St. 8
Temperature(�C)Temperature(�C)Temperature(�C)
Table 1. Mean total length (TL) and body weight (BW) of adult Pacific cod, Gadus macrocephalus sampled for gonadosomatic index (GSI) analy- sis in Jinhae Bay
Female Male
Year Month
TL±SD (cm) BW±SD (g) TL±SD (cm) BW±SD (g)
2006 Dec. 67.4±6.6 3658.7±1187.9 67.1±6.9 3595.9±1221.1
2007 Jan. 64.6±8.1 2477.3±1793.1 64.6±8.6 2499.2±1846.9
Feb. 62.3±6.1 2807.7±877.8 58.0±11.3 2270.0±1400.0
2007 Dec. 66.3±5.0 3744.7±946.4 66.1±4.8 3693.5±868.0
2008 Jan. 67.1±5.7 4013.0±1199.5 65.3±4.1 3496.7±522.5
Feb. 62.6±3.5 3100.9±568.6 62.2±3.4 3061.0±852.5
월이 21.3으로 낮았으나 각 월 지수 값의 변이가 커서 월 간 차이는 유의하지 않았다. 수컷의 GSI 값은 암컷에 비하 여 낮았으나 월간에 큰 차이가 없었다(Fig. 3).
3. 치어 전장 분포
대구 치어는 5, 6월에만 690개체가 채집되었으며, 전장범 위는 37.5~94.9 mm (63.2±11.0 mm; mean±SD)였다. 5월
17일에는 297개체가 채집되었으며, 전장은 37.5~90.1 mm
이며, 5월 29일에는 94개체가 채집되었으며, 전장은45.3~92.5 mm이며, 6월 3일에는 393개체가 채집되었으며
전장범위는 44.1~94.9 mm로 5월과 비슷하지만 80 mm 이 상의 개체는 6월이 더 많았다. 평균전장은 5월 56.6±9.0mm, 6월 68.1±9.7 mm이다(Fig. 4).
4. 치어 이석의 미세구조
대구 치어 690개체의 이석을 연마한 후 관찰하여 일륜
판별이 가능한 273개체를 분석에 이용하였다. 이석 연마는 시상면과 횡단면에 대해서 수행되었으나, 시상면의 경우 이 석의 두께가 두꺼워서 핵 부분이 명확하게 관찰되지 않았 으며, 횡단면의 경우 핵 부분이 명확히 관찰되어 분석에 이
0 10 20 30 40
Female Male 0
10 20 30 40
Dec. 18
Dec. 23 Jan. 10 Feb. 10
Feb. 11 Jan. 11
Month
(a)
(b)
GSIGSI
Fig. 3. Monthly changes in gonadsomatic index (GSI) of female and male of Pacific cod Gadus macrocephalus during the spawning sea- son in Jinhae Bay. a: 2006~2007, b: 2007~2008.
Fig. 4. Length frequency distribution of juvenile Pacific cod, Gadus macrocephalus in Jinhae Bay from May to June 2008.
80
60
40
20
0 80
60
40
20
0
30 40 50 60 70 80 90 100
Total length (mm) 80
60
40
20
0
May 17 n==297
May 29 n==94
June 3 n==299
Number of individualsNumber of individualsNumber of individuals
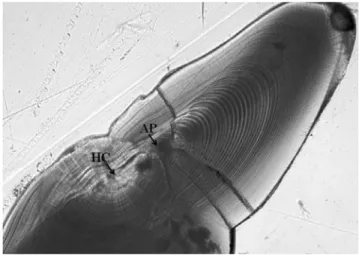
Fig. 5. Microphotograph of frontal plane of sagitta from a juvenile Pacific cod (62.5 mm TL) under the transmitted light. HC: hatch check;
AP: accessory primordium.
용하였다. 이석 횡단면의 경우 원형에서 한쪽으로 길게 늘 어진 모양을 하고 있었다. 전장이 큰 개체의 이석은 연마에 는 2배의 시간이 필요하지만 작은 개체의 이석보다 명확히 관찰할 수 있었다. 이석의 핵 주위에 이차성장 핵이 관찰되 었고, 부화 후부터 이차성장 핵이 나타나는 곳까지의 성장 선 수는 33~42개로 나타났다. 이차성장 핵이 형성된 이후 부터 이석은 한쪽 방향으로 늘어지며 성장선의 폭이 증가 하였다(Fig. 5).
5. 부화일과 체장성장
대구 치어 이석의 미세성장선수는 2008년 5월 17일 표 본에서는 77~150개(평균 102±15), 5월 29일에는 88~151 개(평균 119±16), 6월 3일에는 80~156개(평균 116±18) 의 범위였다. 이석의 미세성장선이 부화 이후부터 하루 주 기로 생성된 것으로 가정하고 추정한 대구 치어 부화일은
12월~3월이고 주로 1월~2월에 부화한 것으로 추정되었
다(Fig. 6).추정한 Gompertz성장식은 다음과 같다(Fig. 7):
TL
t==123.2 exp{-exp[-0.0165 (t-81.814)]}(r
2==0.93, n==273)TL
t는 연령 t (일)에 따른 전장(mm)이며, t는 부화 후 일 수이다.고 찰
어류의 산란에 있어서 산란장의 수온, 염분, 바닥의 부착 기질 또는 저질 등의 물리적인 조건은 매우 중요하며(Mori,
2000), 특히 침성란을 낳는 어류에 있어서 산란에 적당한
장소를 찾는 것이 중요하다고 할 수 있다. Cushing (1990) 은 어류의 산란장과 산란기는 난 또는 부화자어에게 최적 의 조건을 제공할 수 있는 공간과 시기에 의해 결정된다고 하였다. 진해만에 산란회유하는 대구도 겨울철 산란기간 동 안 수온, 산란장 저질 그리고 조류의 세기 등의 물리적, 시 간적으로 제한된 조건 하에서 산란하는 번식특성을 갖고 있 는 것으로 보고되었다(Uchida, 1936; Yamamoto, 1939). 그 러므로 대구의 산란장 및 산란기 환경특성을 구명하는 것 은 대구자원의 관리에 있어서 매우 중요하다. 진해만은 우 리나라 대구 산란장 중 가장 규모가 큰 것으로 알려져 있 으며, 산란기는 대략 12월말~1월말이고, 1월초~중순까지 가 산란성기이며 대구 산란회유시 진해만 표층수온이 12~13� C, 이때 저층수온은 5~9� C로 보고하였다(Uchida, 1936).
또한 Yoseda (1992)는 일본 Noto Island 주변해역에서 대구 산란회유시 수온은 11~13�
C, 산란성기시 저층 수온은 9~
11� C로 보고하였다. Kawamura and Okubo (1950)는 무츠만
대구에 대하여 산란회유시 수온은 11.5�C, 산란성기는 12월
하순이고 이때 수온은 8~9�C로 보고하였다. 이번 연구에
서 진해만의 1월 저층수온은 6~13�C 범위였는데, 2007년
정점 6 그리고 2009년 정점 1과 2에서 10�C와 13� C가 측
정된 것을 제외하면 모든 정점의 1월 수온이 7~9�C에 해
당하므로 84년 전 진해만의 저층수온 범위와 거의 차이가 없었다. 이와 같이 대구의 각 산란장 별 산란 성기는 다르 Fig. 6. Hatching date of juvenile Pacific cod, Gadus macrocephalusestimated from the number of growth increments in otoliths collected in 2008.
45
30
15
0 45
30
15
0
20 31 10 20 31 10 20 29 10 20
Dec. Jan. Feb. Mar.
45
30
15
0
Number of individualsNumber of individualsNumber of individuals
May 17 n==112
May 29 n==57
June 3 n==104
Fig. 7. Growth in total length by the Gompertz model of juvenile Pacific cod Gadus macrocephalus in Jinhae Bay.
100
80
60
40
20
0
75 90 105 120 135 150 165
Age in days after hatching
Total length(mm)
TLt==123.2 exp{-exp[-0.0165 (t-81.814)]}
r2==0.93
지만 산란 성기의 수온이 비슷하여, 각 산란장의 수온에 따 라 산란기가 결정되는 것으로 볼 수 있다.
Yamamoto (1939)는 진해만 대구의 수온에 따른 부화기간
을 9.9�C에서 9.7일과 10.3일로 보고하였고, Uchida (1936)
는 진해만 내에서 가장 낮은 수온인 4~5�C에서 부화까지 18~20일, 6~9� C에서는 12~14일이 소요된다고 보고하였
다. 치어 이석의 미세성장선은 부화 이후부터 하루주기로 생성되므로(Narimatsu et al., 2007) 관찰한 일륜수로부터 부 화된 시기를 역 추정한 결과 12월 20일부터 3월 16일 사이 로 나타났다. 추정한 273개체 중 13%만이 12, 3월에 부화 되었고 나머지는 1, 2월에 부화한 것으로 나타났다. 대구 치 어가 채집된 2008년의 진해만 저층수온 조사결과가 1월 평 균 7.3�C (6.9~8.8� C), 2월 평균 5.6� C (5.3~6.3� C)인 것과
수온에 따른 수정란 부화일 수를 고려할 때, 이번 연구에서 분석된 치어의 경우, 주로 1월과 2월에 산란되었으며, 특히1월 중순~말에 집중적으로 산란된 것으로 추정된다. GSI
평균값도 12, 1, 2월 모두 높게 나타났지만 1월이 제일 높 은 값을 나타내어 치어의 이석 일륜에 근거한 산란기 추정 결과와 잘 일치하였다. 이와 같은 결과는 산란기와 산란기 수온에 대한 Uchida (1936)의 결과와도 잘 일치한다. 결과적 으로 진해만 대구 성어의 GSI와 치어의 이석 일륜 추정 결 과에 근거하여 진해만 대구 산란기는 12~3월이고, 주 산란 기는 1월로 추정된다. 한편 진해만에서 3월 초 전장 약 20mm의 치어, 5월 초~중순 수온 15� C 전후일 때 전장 40~
80 mm, 5월 하순 수온 15~17� C일 때 전장 70~90 mm의
치어가 채집된다고 하였다(Uchida, 1936). 또한 만 내 수온 상승과 치어가 성장함에 따라 6월 중부터 치어가 진해만에 서는 채집되지 않고 만 밖으로 이동한다고 하였는데 이번 연구에서도 대구 치어가 채집된 시기와 채집 시기에 따른 치어의 성장경향은 일치하였으나 5월 하순 이후 채집되는 치어의 전장 범위에서 차이가 있어서 5월 29일과 6월 3일 의 경우 전장 40~70 mm의 개체가 다수 채집되었다. 진해 만 대구 자원관리에 있어서 치어의 출현 시기와 성장은 중 요한데 이번 연구에서는 5월과 6월 초에 채집된 한정된 시 료를 이용하였으므로 이에 대해서는 추가적인 연구가 필요 할 것으로 판단된다.Narimatsu et al. (2007)은 대구 이석의 일륜은 부화 이후
부터 하루 주기로 형성된다고 하였다. 진해만 대구 치어의 이석은 부화 후 38일 정도 지나면 이차성장 핵이 형성되며, 그 이후에는 이석의 각 부위별로 성장속도가 달라지고 성 장선 폭도 넓어진다. 이와 같이 부화 후 이차성장 핵이 형 성되는 기간이 33~42일로 변이가 크지 않은 것은 채집지 역에 서식하는 대부분의 개체가 유사한 환경 조건에서 성 장하였기 때문인 것으로 판단된다. 그러나 Narimatsu et al.(2007)은 태평양 대구의 이차성장 핵의 형성은 51~70번째
사이에서 형성된다고 하여 이번 연구 결과와 차이가 있었다. 이차성장 핵의 형성은 변태(Toole et al., 1993; Modin et
al., 1996), 먹이전환(Marks and Conover, 1993; Secor et al., 1995), 서식지 변화(Sogard, 1991; Plaza et al. 2001) 과정 중
에 나타난다고 하였다. 또한 이차성장 핵이 관찰되는 어류의 이석에서 이차성장 핵은 변태 이후 형성되는 것이 보고되어(Nishimura and Yamada, 1984; Lee and Byun, 1996), 이차성
장 핵까지의 일륜수는 변태까지의 기간을 나타내는 지표로 이용될 수 있다고 하였다(Campana, 1984). 대서양 대구를 제 외한 대구 속 중에서(Hussy et al., 2003), 이차성장 핵은 저 서생활단계 또는 착저 이전에 나타난다고 보고되었다(Qui-nonez-Velazquez, 1999; Arneri and Morales-Nin, 2000). 그러
나 태평양 대구는 대략 전장 25 mm의 자어 단계에 변태를 하고(Matarese et al., 1989; Takatsu et al., 1995), 저층바닥에 착저하는 대략 전장 45 mm일 때 치어 단계에 이른다고 하 였다(Takatsu et al., 1995; Morioka and Kuwata, 2002). 이차 성장핵의 형성 도중에 대구는 아직 가슴지느러미 골화를 완 료하지 않았으며, 먹이의 급격한 변화는 성장기간 도중에 확인할 수 없었다(Matarese et al., 1989; Takatsu et al., 1995).Narimatsu et al. (2007)에 의하면 대구의 가슴지느러미 골화
는 이차성장핵의 형성과 연관될 가능성이 있다고 하였다.그리고 어류의 치어기에는 지느러미 등의 다른 골격 형성 에 주로 집중되었던 탄산칼슘 성분이 변태를 마친 후 체장 과 이석 성장으로 전이되며, 이 시기에 체형과 행동양식이 변화됨에 따라 이석에 이차성장핵이 생성되고 환경과 먹이 섭식에 적응하여 이석 성장이 빨라지고 일일성장선 폭도 넓어지는 것으로 보고하였다(문과 이, 1999). 그러므로 인 공적으로 대구를 길러 치어 이석을 분석한 Narimatsu et al.
(2007)의 결과와 자연에서 채집한 대구 이석의 이차성장핵
형성시기의 차이는 먹이섭식이나 환경적인 영향에 의한 것 으로 추측되며, 이석의 미세구조 분석 시 서식환경에 따른 성장차이를 고려해야 할 것으로 생각된다.요 약
진해만 대구의 산란기 수온, 부화일과 초기성장을 추정하 였다. 2006년부터 2009년 사이 11월에서 5월까지 매월 8개 의 정점의 수온을 측정하여 분석한 결과 주 산란기의 수온 은 7~9�
C였다. 2006~2008년의 주산란기인 12월에서 2월
사이 산란어의 생식소숙도지수(GSI)는 수컷에 비하여 암컷 이 유의하게 컸으며, 1월에 최대값을 보였다. 2008년 5~6 월에 채집된 치어의 전장은 37.5~94.9 mm (63.2±11.0mm; mean±SD) 범위였으며, 이석의 부화 이후 일륜수의
평균(±SD)은 5월 17일 표본에서는 102±15, 5월 29일에 는 119±16, 6월 3일에는 116±18였다. 이석의 일륜수로부 터 추정한 부화일은 12월 하순에서 3월 중순이었으며, 주부화 시기는 1월 하순에서 2월 중순이었다. 부화 이후 일 령 (t)에 대한 전장 (TL, mm)의 성장은 Gompertz 성장식,
TL
t==123.2 exp {-exp [-0.0165 (t-81.814)]} (r2==0.93, n==273)로 나타낼 수 있었다.
사 사
진해만 수온 조사 시 많은 도움을 주신 경상대학교 실습 선 참바다호 관계자 분들께 감사드립니다. 그리고 현장조사 시 많은 도움을 준 경상대학교 해양생명과학과 어류학 및 해양생물유전학연구실 학생들에게 진심으로 감사드립니다.
인 용 문 헌
문형태∙이태원. 1999. 이석의 미세구조를 이용한 가덕도 천해역 문치가자미(Limanda yokohamae) 유어의 연령과 성장. 한 국어류학회지, 11: 46-51.