─
─ 224 ──
감성돔
, Acanthopagrus schlegeli
의 담수순화 과정에서 나타나는 생리학적 반응민병화∙김범광∙허준욱∙방인철*∙변순규**∙최철영***∙장영진 부경대학교 양식학과, *순천향대학교 생명과학부,
**국립수산과학원 양식환경 연구소, ***미국 국립보건원
Physiological Responses during Freshwater Acclimation of Seawater-Cultured Black Porgy
(Acanthopagrus schlegeli)
Byung Hwa Min, Bum Kwang Kim, Jun Wook Hur, In Chul Bang*, Sun Kyu Byun**, Cheol Young Choi*** and Young Jin Chang Department of Aquaculture, Pukyong National University, Busan 608-737, Korea
*Department of Biological Resources, Soonchunhyang University, Asan 336-745, Korea
**Aquaculture Environment Institute, National Fisheries Research & Development Institute, Tong-yong 650-943, Korea
***Laboratory of Gene Regulation and Development, National Institute of Child Health and Human Development, National Institutes of Health (NIH), Bethesda, MD 20892, USA
Physiological responses (cortisol, glucose, Na±, Cl-, osmolality, aspartate aminotransferase (AST), alanine aminotransferase (ALT), hematological factors, and survival rate) were investi- gated during freshwater acclimation of seawater-cultured black porgy (Acanthopagrus schle- geli). As control, in seawater, there were no significant differences in the investigated items during the experimental period. In step-salinity change, plasma cortisol level was significantly increased from 13.0±1.4 ng/mL at the beginning to 29.5±14.8 ng/mL at 5 days, and recovered to the basal level (19.7±1.6 ng/mL) at 10 days. Glucose showed a tendency to co-increase with cortisol. Plasma Na±and Cl-concentrations decreased from 176.7±3.8 and 152.7±1.5 mEq/L to 111.8±38.0 and 73.5±26.5 mEq/L, respectively, at 5 days. But these recovered as completely as the basal levels at the end of the experiment (20 days). Also, changes of osmolality levels showed a similar tendency with Na±and Cl-levels. Although AST was markedly elevated at 12 hours, its level soon recovered to the basal level. No significant differences in Hct, RBC and Hb were found during the experimental period. Survival rate was 86.7% at 5 days and 83.3% at 20 days.
The results presented in this paper suggest that it may be possible to culture black porgy in freshwater.
Key words : Black porgy, Acanthopagrus schlegeli, freshwater acclimation, step-salinity change, physiological response
서 론
감성돔, Acanthopagrus schlegeli은 수컷의 생식소가 먼저 나타나는 인접적 자웅동체(consecutive hermaph-
rodite) 어류로서 연안의 암반주위에 서식하여 낚시어종
으로 인기를 끌고 있으며, 경제성이 높은 양식대상 어종 인 것으로 알려져 있다. 또한 감성돔은 숭어, Mugil cephalus, 농어, Lateolabrax japonicus와 함께 염분 내성 이 강한 광염성 어종으로서(Kimura and Tanaka, 1991), 최근 본 종의 이러한 특성인 강한 염분내성을 이용하여 담수순화 양식을 위한 연구가 진행되어져 오고 있다 (Chang et al., 2002). 감성돔의 담수순화 양식이 가능해 진다면 내수면 어류양식의 활성화를 기대할 수 있을 뿐 만 아니라, 적조로 인한 피해 감소, 질병의 역치료 등 여 러 가지 잇점을 내다 볼 수 있다.
이러한 해산어류의 담수순화 양식에서는 담수순화 방 법, 어체의 건강상태, 크기 및 사육 환경 등이 중요한 요 인으로 작용한다. 이러한 요인들 중 담수순화 방법 즉, 사육수의 인위적인 염분변화는 어체의 생리반응에 큰 영향을 미치는 요인이 되며, 어류가 감지하는 가장 큰 스트레스 요소 중의 하나로서 어류의 혈중 coltisol 농도 를 상승시키는 역할을 한다(Chang and Hur, 1999). 어 류가 지속적으로 스트레스에 노출되게 되면, 어병에 대 한 저항성이 감소하게 되고, 어류의 성장 및 번식능력이 떨어지는 것으로 알려져 있다(Wedemeyer and Yasu- take, 1977; Pickering et al., 1987). 어류가 스트레스를 감지하게 되면, 시상하부-뇌하수체-간신선축으로의 정 보전달을 통하여 cortisol이 혈중으로 분비된다(Specker et al., 1989). 시상하부와 뇌하수체는cortisol을 조절하는 기관으로, 특히 뇌하수체 전엽의 간신세포로부터 분비되 는 부신피질 자극호르몬(adrenocorticotropic, ACTH)은 cortisol 분비와 합성을 조절하는 호르몬으로 알려져 있 다(Sumpter et al., 1994). 또한 어류에서의 cortisol은 어 체의 항상성 유지를 위해 글루코스 신생합성(gluconeo- genesis)을 통하여 glucose의 혈중 분비를 증가시킨다 (Chang and Hur, 1999).
현재까지 감성돔의 담수순화에 관한 연구로는 급격한 염분변화에 따른 혈장 cortisol, 이온변화 등 스트레스와 연관된 혈액생리학적인 측면의 실험결과가 보고되어 있 을 뿐(Chang et al., 2002), 아직 국내외적으로 담수순화 방법과 담수사육에 대한 체계적인 연구결과가 얻어지지 않은 실정이다.
본 연구에서는 해수에서 사육된 감성돔을 단계적인 염분변화를 통하여 담수로 순화시키는 과정에서, 혈장
cortisol 및glucose 농도 등 감성돔 어체내에서의 생리학 적 반응을 조사하여 어류가 감지하는 스트레스의 수준 을 제시하고, 이를 기초로 감성돔 담수순화 양식을 위한 기초자료를 얻고자 하였다.
재료 및 방법
1. 실험어와 실험조건
해수에서 사육하던 평균전장 12.1±0.7 cm, 평균체중 30.6±6.5 g의 감성돔 총 240마리를 실험에 사용하였다. 실험은 순환여과 사육시스템으로 구성된8개의 FRP 원 형수조(수용적 220 L)에 각각 30마리씩 2반복으로 수용 하였다.
120마리의 실험어가 수용된 실험구(4개 수조)의 염분 을 시간당 2‰씩 낮추어 실험개시 12시간째에 10‰의 염분농도가 되도록 조절한 후, 10‰의 농도를72시간 동 안 유지한 다음, 다시 단계적으로 염분 농도를 낮추어 12시간째에 0‰의 완전 담수가 되도록 설정하였다. 대조 구인 4개 수조의 실험어 120 마리는 35‰의 완전 해수 에서 사육하였다(Fig. 1). 실험기간중 수온은 18~23�C, 용존 산소량은 5.8~6.6 mg/L였으며, 자연 광주기 상태 로 실험을 실시하였다. 실험기간중, 상업용 돔 사료를 어 체당2%씩 공급하였다.
2. 혈액의 채취와 분석
각 실험구의 실험어 6 마리로부터 heparin을 처리한 주사기(1 mL-23 G)를 사용하여 마취없이 미부혈관에서 30초 이내에 혈액을 채취하였다. 실험어로부터 혈액을 채취하기 이전에 공급한 먹이가 어체의 혈액성상에 미 치는 영향을 최소화하기 위하여 채혈24시간 전부터 절 식시켰다. 혈액은 0 (35‰), 12, 84 (10‰), 96시간째, 5일, 10일 및20일째(0‰)에 각각 채혈하였다(Fig. 1).
채취한 혈액은 상온에서 10분간 방치한 후, 5,600×g 로 5분간 원심분리한 후, 얻어진 혈장을 -72�C로 냉동 시켜 보관하면서 실험에 사용하였다. 혈장 cortisol 농도 는 Donaldson (1981)의 방사면역측정법(RIA)에 따라 cortisol RIA kit (DPC, Los Angeles, U.S.A.)로 항원과 표 지항원이 항체에 경쟁적으로 반응하도록 유도한 다음, Hewlett Packard Gamma Counter (Cobra II 5010, Pack- ard Co., U.S.A.)로 측정하였다. 혈장 glucose, Na±, Cl- 농도, AST (aspartate aminotransferase) 및 ALT (alanine aminotransferase)는 Chemistry Autoanalyzer (Advid 1650, JEOL Co., Japan)를 사용하여 측정하였으며, 혈장
삼투질 농도는 micro-osmometer (3MO plus, Advanced Instruments Inc., USA)를 사용하여 측정하였다. 채혈 직후에 자동혈액분석기(H5M, SEAC co., Italy)를 사용하 여, 적혈구 용적(hematocrit, Hct), 적혈구 수(red blood cell, RBC) 및 혈색소 농도(hemoglobin, Hb)를 측정하였 다.
3. 생존율 조사
실험기간중 실험구와 대조구에서 매일 폐사개체를 파 악하여 폐사율을 구하고, 이로부터 생존율을 역산하여 구하였다.
4. 통계처리
각 실험결과로부터 얻어진 자료값 사이의 유의차 유 무는 SPSS-통계 패키지(version 9.0)에 의한 ANOVA 및 Duncan’s multiple range test로 검정하였다.
결 과
1. 혈장의 cortisol 및 glucose 농도
실험기간 중, 어체내에서의 cortisol과 glucose의 농도 변화는 Fig. 2와 같다. Cortisol 농도는 실험개시시 13.0
±1.4 ng/mL에서 실험개시 5일째 29.5±14.8 ng/mL로 유의하게 증가하였다. 그 후, 10일째부터 감소하기 시작 하여 실험종료시(20일째)에는 7.5±2.3 ng/mL로 실험개 시시 수준으로 회복되었다. 대조구에서는 실험개시시부 터 종료시까지 9.1±1.9~11.6±3.3 ng/mL로 유의한 변 화를 보이지 않았다. Glucose 농도는 실험개시시 52.3±
7.2 mg/dL에서 실험개시 5일째까지는 변화를 보이지 않 았지만, 실험개시 10일째에74.0±1.4 mg/dL로 유의하게 증가하였다(P⁄0.05). 그 후, glucose 농도는 감소하기 시 작하여 실험종료시에는 66.5±0.7 mg/dL로 실험개시시 수준으로 회복되었다. 대조구에서는 실험개시시부터 종 료시까지 51.5±0.7~63.5±6.4 mg/dL로 유의한 변화를 보이지 않았다.
2. 혈장의Na± 및 Cl- 농도 및 삼투질 농도
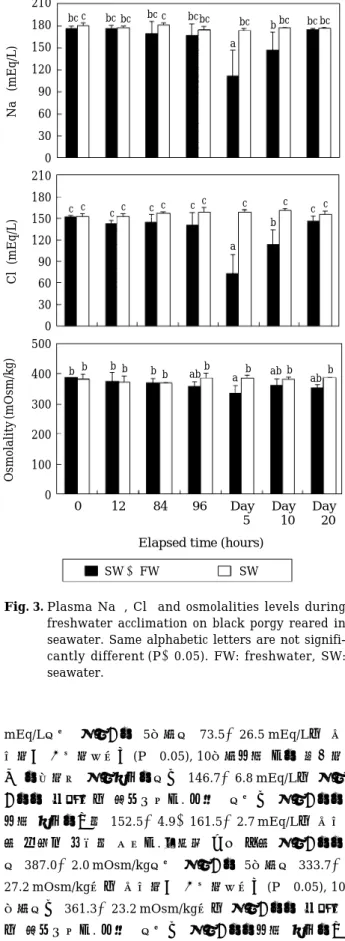
실험기간 중, Na±, Cl-및 삼투질 농도의 변화가 관찰 되었다(Fig. 3). Na±의 농도는 실험개시시 176.7±3.8 mEq/L에서 실험개시 5일째 111.8±38.0 mEq/L로 유의 하게 감소하였으나(P⁄0.05), 10일째부터 증가하기 시작 하여 실험종료시에는 175.3±1.5 mEq/L로 실험개시시 수준으로 회복되었다. 대조구에서는 실험개시시부터 종 료시까지 173.7±4.1~180.7±3.8 mEq/L로 유의한 변화 를 보이지 않았다. Cl-의 농도는 실험개시시 152.7±1.5 Fig. 1. Salinity changes designed for the experiment.
Arrows indicate the blood sampling times.
Fig. 2. Plasma cortisol and glucose levels during fresh- water acclimation on black porgy reared in sea- water. Same alphabetic letters are not significant- ly different (P¤0.05). FW: freshwater, SW: sea- water.
0 12 24 36 48 60 72 84 96 D-5 D-10 D-20 Time (hours)
25 20 15 10 5 0
Salinity(‰) 30
20
10 0
ab
ab ab
ab ab
ab ab
abc abc
abc abc
ab ab ab
ab ab ab
bc
bc ab ab
ab ab a
a a
c
Cortisol(ng/ml)Glucose(mg/dl)
100
80
60
40
20
0
SW→FW SW
0 12 84 96 Day Day Day
-5 -10 -20
Elapsed time (hours)
mEq/L에서 실험개시 5일째에 73.5±26.5 mEq/L로 유 의하게 감소하였으나(P⁄0.05), 10일째부터 다시 증가하 기 시작하여 실험종료시에는 146.7±6.8 mEq/L로 실험 개시시 수준으로 회복되었다. 대조구에서는 실험개시시 부터 종료시까지 152.5±4.9~161.5±2.7 mEq/L로 유의 한 변화를 보이지 않았다. 삼투질 농도 또한 실험개시시 에 387.0±2.0 mOsm/kg에서 실험개시 5일째에 333.7± 27.2 mOsm/kg으로 유의하게 감소하였으나(P⁄0.05), 10 일째에는 361.3±23.2 mOsm/kg으로 실험개시시 수준으 로 회복되었다. 대조구에서는 실험개시시부터 종료시까 지 383.0±15.6~388.7±35.0 mOsm/kg으로 유의한 변
화를 보이지 않았다.
3. 혈장의AST 및ALT
AST 및 ALT의 변화는 Table 1에 나타내었다. AST는 실험개시시에 35.7±8.1 IU/L에서 실험개시 12시간째에 50.0±2.8 IU/L로 유의하게 증가하였지만(P⁄0.05), 12 시간 이후로는 오히려 실험개시시보다 유의하게 낮게
나타났다(P⁄0.05). 대조구에서는 실험개시시부터 종료
시까지30.5±6.4~41.5±10.6 IU/L로 유의한 변화를 보 이지 않았다. ALT는 실험구와 대조구 공히 실험개시시 부터 종료시까지 각각 1.5±0.7~3.8±0.5 IU/L와 1.5± 0.2~2.1±0.6 IU/L로 유의한 변화를 보이지 않았다.
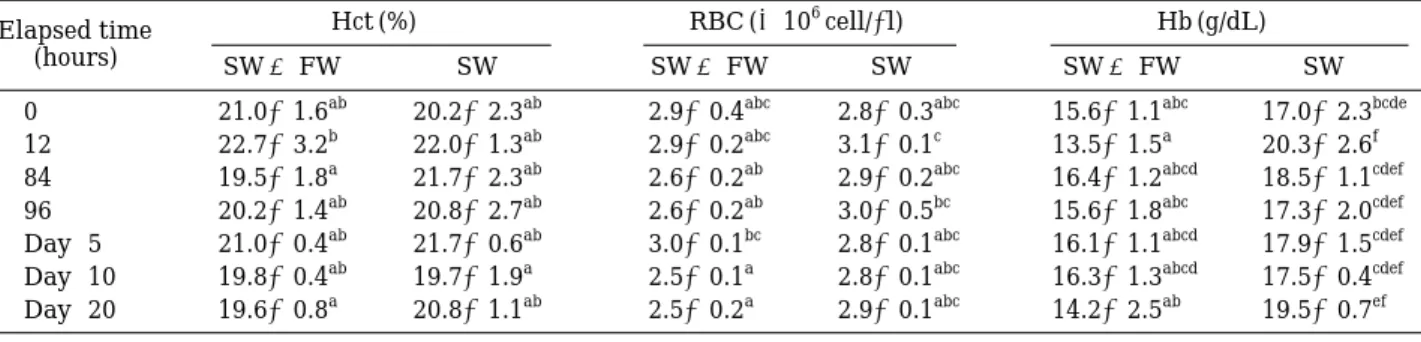
4. 전혈의Hct, RBC 및Hb
Hct, RBC 및 Hb의 변화는 Table 2에 나타내었다. Hct 는 실험구와 대조구 모두 실험개시시부터 종료시까지 각각 19.6±0.8~22.7±3.2%와 19.7±1.9~22.0±1.3%
로 유의한 변화를 보이지 않았다. RBC는 실험구와 대조 구에서 실험개시시부터 종료시까지 각각 2.5±0.1~3.0
±0.1×106cell/µL와 2.8±0.1~3.1±0.1×106cell/µL로 유의한 변화를 보이지 않았다. Hb 또한 실험구와 대조구 에서 실험개시시부터 종료시까지 각각 13.5±1.5~16.4
±1.2 g/dL와 17.0±2.3~19.5±0.7 g/dL로 유의한 변화 를 보이지 않았다.
5. 생존율
실험기간중 실험구에서의 감성돔의 생존율은 0‰ 염 분농도였던 실험개시 4일째에 90%로 감소하기 시작하 여, 5일째에 86.7%, 20일째에는 83.3%로 나타났다. 대조 Fig. 3. Plasma Na±, Cl- and osmolalities levels during
freshwater acclimation on black porgy reared in seawater. Same alphabetic letters are not signifi- cantly different (P¤0.05). FW: freshwater, SW:
seawater.
Table 1. Plasma AST and ALT levels during freshwater acclimation on black porgy reared in seawater Elapsed time AST (IU/L) ALT (IU/L)
(hours) SW→ FW SW SW→ FW SW
0 35.7±8.1b 32.3±4.7ab 2.3±0.6ab 1.8±0.3a 12 50.0±2.8c 31.7±6.5ab 3.8±0.5b 1.9±0.3a 84 21.7±4.5a 39.2±3.3bc 2.3±1.5a 1.7±0.5a 96 22.0±2.0a 37.0±9.9bc 2.4±0.5ab 1.9±0.3a Day-5 20.5±0.7a 30.4±5.2ab 2.8±1.0ab 2.0±0.7a Day-10 20.0±6.6a 39.5±6.4bc 2.0±1.4a 1.6±0.5a Day-20 21.5±9.2a 33.5±4.9ab 1.5±0.7a 1.5±0.2a The values are mean±SD. Means within each item followed by the same alphabetic letter are not significantly different (P¤0.05).
AST: aspartate aminotransferase, ALT: alanine aminotransferase, FW: freshwater, SW: seawater.
210 180 150 120 90 60 30 0 Na± (mEq/L)Cl- (mEq/L)Osmolality(mOsm/kg)
210 180 150 120 90 60 30 0 500 400 300 200 100 0
0 12 84 96 Day Day Day
-5 -10 -20
Elapsed time (hours)
SW→FW SW
bcc bc bc bc bcbc bc bc bc bc
c c c c c c c c c c
c c
a
a c
a b
b
b b b b b b abb b abb b
ab
구에서는 실험종료시인 20일째에 96.7%로 감소하였다 (Fig. 4).
고 찰
해산어류인 감성돔의 담수순화 과정중 사육수의 염분 변화는 어류가 감지하는 스트레스 중 가장 큰 요인으로 작용하여 이러한 영향이 어류의 혈장 cortisol 및 cate-
cholamine 등의 농도를 상승시키는 결과를 유도한다
(Mazeaud and Mazeaud, 1981; Schreck, 1981; Pickering and Pottinger, 1995). Cortisol은 대사 에너지, 물-이온 평형, 산소 uptake, 면역 능력을 조절하는 기능을 갖고 있으며(Wendelaar Bonga, 1997), 이러한 조절기능으로 인하여 어류가 스트레스 요인에 대하여 적응할 수 있게 되는 것으로 알려져 있다(Barton and Iwama, 1991). 또 한, 스트레스로 인한 혈장 cortisol의 증가는 어체에 유 해한 영향을 미치게 되어, 성장억제 및 성성숙을 지연시 킨다는 보고도 있다(Pickering et al., 1987). 일반적으로 어류가 여러 스트레스 요인들에 노출되게 되면, 정보전
달에 의하여 시상하부-뇌하수체-간신선축에 영향을 미 치게 되고, 결국 혈중으로 cortisol이 분비된다(Wende- laar Bonga, 1997). Cortisol 합성에 미치는 스트레스 요 인의 영향은porgy, Sparus aurata (Mosconi et al., 1998;
Rottland et al., 2001; Sunyer et al., 1995), red porgy, Pargus pargus (Rotlland and Tort, 1997) 및 rainbow trout, Oncorhynchus mykiss (Contreras-Sanchez et al., 1998; Ruane et al., 1999) 등에서 보고된 바와 같이 담수 및 해수어종 모두에서 나타나고 있다.
본 연구에서도 해수에서 담수로의 단계적인 염분변화 는 실험어인 감성돔에 스트레스 요인으로 작용하여 담 수순화 5일째에 cortisol 농도의 수준이 29.5±14.8 ng/
mL로서 안정시때의 13.0±14.8 ng/mL 보다 2배 정도 증가되었다. 이러한 결과로 보아, 10‰ 해수에서 담수로 의 전환 후 24시간 이내에 강한 스트레스를 받고 있음 을 알 수 있다. Chang et al. (2002)은 감성돔을 중간 순 화과정을 거치지 않고 곧장 해수에서 담수로 옮겼을 때 cortisol의 농도가 3시간째 10배로 증가하였다고 보고하 였다. 이러한 결과로 보아, 해수에서 담수로의 급격한 염 분변화는 단계적인 염분변화보다 어류가 느끼는 스트레 스의 반응이 빨리 나타나며, 또한 강한 스트레스를 받고 있는 것임 알 수 있다. 그러나, 이미 담수에 적응된 감성 돔을 해수로 즉시 옮겼을 때 cortisol의 농도변화는 유 의한 차이를 보이지는 않았지만, 다소 증가하는 경향이 있다고 보고하였다(Chang et al., 2002). Chang et al.
(2002)이 보고한 cortisol 농도의 증가는 두가지 측면에 서 설명할 수 있다. 첫째, 급격한 염분변화에 따른 어류 의 스트레스 반응일 가능성, 둘째, 어류의 혈장 cortisol 이 아가미 염류세포에서 Na±, K±-ATPase 활성 및 Na±, K±-ATPase α-subunit의 발현을 촉진시켜 해수적 응과 염분내성을 증가시켰다는 보고(Pickford et al., 1970)에서와 같이, 감성돔을 담수에서 해수로 옮길 때의
(hours) SW→ FW SW SW→ FW SW SW→ FW SW
0 21.0±1.6ab 20.2±2.3ab 2.9±0.4abc 2.8±0.3abc 15.6±1.1abc 17.0±2.3bcde
12 22.7±3.2b 22.0±1.3ab 2.9±0.2abc 3.1±0.1c 13.5±1.5a 20.3±2.6f
84 19.5±1.8a 21.7±2.3ab 2.6±0.2ab 2.9±0.2abc 16.4±1.2abcd 18.5±1.1cdef 96 20.2±1.4ab 20.8±2.7ab 2.6±0.2ab 3.0±0.5bc 15.6±1.8abc 17.3±2.0cdef Day-5 21.0±0.4ab 21.7±0.6ab 3.0±0.1bc 2.8±0.1abc 16.1±1.1abcd 17.9±1.5cdef Day-10 19.8±0.4ab 19.7±1.9a 2.5±0.1a 2.8±0.1abc 16.3±1.3abcd 17.5±0.4cdef Day-20 19.6±0.8a 20.8±1.1ab 2.5±0.2a 2.9±0.1abc 14.2±2.5ab 19.5±0.7ef The values are mean±SD. Means within each item followed by the same alphabetic letter are not significantly different (P¤0.05). Hct:
hematocrit, RBC: red blood cell, Hb: hemoglobin, FW: freshwater, SW: seawater.
Fig. 4. Changes of survival rate of black porgy during experimental period.
120 100 80 60 40 20 0
40 30 30 25 20 15 10 5 0
Survival(%) Salinity(‰)
0 2 4 6 8 10 12 14 16 18 20
Rearing time (days)
SWÆFW SW Salinity
cortisol의 증가는 아가미 염류세포의 활성을 향상시려 는 작용일 가능성, 이러한 두가지 측면으로 해석될 수 있다. 본 연구에서 해수에서 담수로의 순환시, 실험개시 5일째에 생존율이 90%로 감소하는 결과를 보였으며, 이 시기는 혈장 cortisol 농도가 최고값을 나타내는 시점과 일치하였다. Chang and Hur (1999)는 틸라피아의 경우, 담수에서 해수로 전환하였을때 혈장 cortisol의 급격한 증가와 함께 매우 낮은 생존율을 보고한 바 있다. 따라 서 혈장 cortisol의 급격한 상승은 생존율 감소와 밀접 한 연관성이 있음을 알 수 있다. 그러나, 실험개시5일째 이후로는 생존율의 변화가 거의 없는 것으로 나타난 것 으로 보아, 이는 감성돔이 담수에 잘 적응하고 있는 것 으로 판단되어, 본 연구에서는 측정하지 않았다.
Barton and Iwama (1991)는 glucose 농도는 스트레스 에 의해 증가되며, cortisol 농도가 증가함에 따라glucose 농도가 증가하는 현상은 스트레스에 의한 호르몬 반응 에 따른2차적인 반응이라고 보고하였다. 이러한 결과는 숭어, Mugil cephalus (Chang and Hur, 1999), pejerrey, Odontesthes bonariensis (Tsuzuki et al., 2001)등 많은 어류에서 보고되었으며, 본 연구에서도 cortisol과 glu- cose의 동반상승이라는 동일한 결과가 나타났다. 한편, Chang et al. (2002)은 감성돔을 해수에서 담수로 즉시 옮긴 후 3시간째 cortisol 농도는 유의하게 증가한 반면, 실험개시 24시간 후까지 glucose 농도는 변화가 없는 것 으로 보고하였다. 이와 같은 결과에 대하여, Chang et al.
(2002)은 cortisol에 의해 glucose 신생합성이 일어나기
이전에 glucose가 항상성 유지를 위해 에너지원으로 급
격히 사용되었을 가능성, 그리고 또 다른 가능성으로 해 수에서 담수로 옮긴 후 3시간이 경과되기 이전에 glu- cose가 cortisol과 함께 동반 상승하였다가, cortisol보다 먼저 스트레스 이전의 농도로 회복되었을 가능성에 대 하여 언급한 바 있다.
일반적으로 어류는 환경수의 삼투질 농도가 체내보다 높으면 삼투압 조절에 의하여 이온은 유입되며 물은 빠 져나가게 되므로, 어류는 체내의 일정한 삼투질 농도를 유지하기 위하여 이온은 방출하며 물은 흡수하는 저삼 투압조절(hypo-osmoregulation) 능력과, 반대로 환경수 의 삼투질 농도가 체내보다 낮으면 이온은 빠져나가며 물은 유입되므로 어류는 체내의 항상성을 유지하기 위 하여 이온은 체내로 흡수되고 물은 체외로 방출하는 고 삼투압조절(hyper-osmoregulation) 능력을 갖고 있다. 그러나, 본 연구에서와 같이, 해수에서 담수로 염분변화 시 어류는 체액보다 고장액에서 저장액으로 노출되게 되므로 어체는 저삼투압조절에서 고삼투압조절로의 전 환이 빠른 시간내에 제대로 조절되지 않아 체내의 Na±
및 Cl-이 환경수로 빠져나갔을 가능성이 있다. 이와 같 은 가능성은Sampaio and Bianchini (2002)가 보고한 광 염성 어종인 가자미, Paralichthys orbignyanus에서 환 경수의 염분 농도가 낮아짐에 따라 혈장의 Na±및 Cl- 의 수준이 낮아졌다는 결과로서 뒷받침된다. 본 연구에 서는 해수에서 담수로 전환된지 24시간째(실험 5일째) 에 Na±및 Cl-의 농도는 감소하였지만, 해수에서 담수 로 도달할 때까지 유의한 차이가 없는 점으로 보아, 감 성돔은 삼투질조절 능력이 뛰어난 어종일 가능성도 제 기된다.
또한 본 실험에서 측정된 혈액의 AST, ALT는 모두 아민기 전이효소의 일종으로 간, 비장 등의 세포에 분포 하고 있는데, 어체가 정상적인 경우에는 혈중의 활성농 도가 낮지만, 조직에 괴사가 일어나거나, 병적 증상이 나 타날 때는 세포 외로 방출됨으로써 혈중 농도가 높아지 게 된다( 등, 1986). 본 연구의 경우, 10‰ 해수에서 는AST의 증가가 나타났으나, Chang et al. (unpublished data)의 연구결과, 35‰에서 사육한 감성돔을 5‰로 바 로 옮기더라도AST에는 변화가 없는 점으로 보아, 환경 수의 암모니아와 같은 환경 요인으로 의한 일시적인 증 가로도 볼 수 있다. 그러나, 감성돔을 해수에서 담수로 바로 옮겼을 때AST의 수준이 급격하게 증가하는 것으 로 나타났다(Chang et al., unpublished data). 따라서, 담 수순화 방법중 단계적인 염분변화의 방법은 급격한 염 분변화를 최소화할 수 있어, 간이나 비장 등의 기관을 구성하고 있는 세포의 생리적 부담을 줄일 수 있을 것 으로 판단된다.
Hct, RBC 및Hb 등의 혈액 인자는 생체내에서의 산소 운반 능력을 판단하는 척도이다. Davis and Parker (1990)는 해산어류의 스트레스는 일반적으로 혈중 Hct, RBC 및 Hb 등의 농도를 증가시킨다고 보고하였다. 그러 나, 본 연구에서는 유의한 차이를 나타내지 않는 점으로 보아 해수에서 담수로의 단계적인 염분변화의 경우, 감 성돔은 산소수급에 문제가 있을 정도의 스트레스는 받 고 있지 않는 것으로 판단된다.
본 연구의 결과를 종합해 볼 때, 해수에서 담수로의 단계적인 염분변화에 따른 감성돔의 순화의 경우, 어체 의 스트레스 반응은 유발하였으나 단계적인 염분변화 없이 직접 해수에서 담수로 급격하게 염분을 변화시킨 감성돔에 비하여 cortisol, glucose 및 삼투질농도 등의 변화가 낮게 나타났으며(Chang et al., 2002), 혈액성상의 유의한 변화 또한 인정되지 않았다. 따라서 감성돔의 담 수순화시 단계적인 염분변화 방법은 급격한 염분 변화 의 방법보다 어체의 생리상태에 크게 영향을 미치지 않 는 것으로 사료된다. 앞으로의 연구는 본 연구에서 제시
과 내분비학적 방법을 통하여 감성돔의 장기간 사육시 성장, 생존율 및 건강도 등도 함께 검토해 보아야 할 것 으로 생각된다.
적 요
해산어류의 담수순화 양식을 위한 기초자료를 얻고자, 단계적인 염분변화의 방법을 이용하여 감성돔을 해수에 서 담수로 순화시켰을 때 여러 생리학적 반응을 조사하 였다.
Cortisol 농도는 실험개시시에 13.0±1.4 ng/mL였던 것이 실험개시 5일째 29.5±14.8 ng/mL로 유의하게 증 가하였지만 실험개시 10일째에 19.7±1.6 ng/mL로 실험 개시시 수준으로 회복되었다. Glucose 농도는 cortisol과 동반 상승하는 경향이 나타났다.
Na±및 Cl-는 실험개시시에 각각 176.7±3.8 mEq/L, 152.7±1.5 mEq/L였던 것이 실험개시 5일째 111.8± 38.0 mEq/L, 73.5±26.5 mEq/L로 유의하게 감소하였으 나, 실험종료시에는 실험개시시 수준으로 회복되었다. 삼 투질 농도 또한 이와 비슷한 경향을 나타내었다. AST는 실험개시 후 12시간째에 증가하였지만 그 이후 실험개 시시 수준으로 감소하였다. Hct, RBC 및 Hb는 실험기간 동안 유의한 변화가 관찰되지 않았다.
생존율은 실험구에서 실험개시 4일째에 90%로 감소 하기 시작하여 실험개시 5일째에는 86.7%, 실험개시 20
일째에는 83.3%로 하락하였으나, 대조구에서는 실험종
료시인20일째에96.7%의 생존율을 보였다.
이상의 결과를 종합해 볼 때 본 연구는 단계적인 염 분변화를 통한 감성돔 담수순화 양식의 가능성이 있음 을 알 수 있었다.
사 사
이 논문은 해양수산부 해양수산개발원 수산특정연구 개발사업(20000108)에 의해 지원되었으며 이에 감사 드 립니다.
인 용 문 헌
Barton, B.A. and G.K. Iwama. 1991. Physiological changes
Chang, Y.J., B.H. Min, H.J. Chang and J.W. Hur. 2002.
Comparison of blood physiology in black porgy (Acan- thopagrus schlegeli) cultured in converted freshwater from seawater and seawater from freshwater. J. Korean Fish. Soc., 35 : 595~600 (in Korean).
Chang, Y.J. and J.W. Hur. 1999. Physiological responses of grey mullet (Mugil cephalus) and Nile tilapia (Oreochro- mis niloticus) by rapid changes in salinity of rearing water. J. Korean Fish. Soc., 32 : 310~316 (in Korean).
Contreras-Sanchez, W.M., C.B. Schreck, M.S. Fitzpatrick and C.B. Pereira. 1998. Effects of stress on the repro- ductive performance of rainbow trout (Oncorhynchus mykiss). Biol. Reprod., 58 : 439~447.
Davis, K.B and N.C. Parker. 1990. Physiological stress in striped bass: Effect of acclimation temperature. Aqua- culture, 91 : 349~358.
Donaldson, E.M. 1981. The pituitary-interrenal axis as an indicator of stress in fish. Academic Press, London, 11pp.
Kimura, R. and M. Tanaka. 1991. Prolactin production during larval and early juvenile periods of euryhaline marine fish, black sea bream Acanthopagrus schlegeli.
Nippon suisan gakkaishi/bull. Jap. Soc. Sci. Fish., 57 : 1833~1837.
Mazeaud, M.M. and F. Mazeaud. 1981. Adrenergic res- ponses to stress in fish. In: Pickering, A.D. Ed. , Stress in Fish. Academic Press, London, pp. 49~75.
Mosconi, G., A. Gallinelli, A.M. Polzonetti-Magni and F.
Facchinetti. 1998. Acetyl salmon endorphin-like and interrenal stress response in male gilthead sea-bream, Sparus aurata. Neuroendocrinology, 68 : 129~134.
Pickering, A.D. and T.G. Pottinger. 1995. Biochemical effects of stress. In: Hochachka, P.W., T.P. Mommsen.
Eds., Biochemistry and Molecular Biology of Fishes, vol. 5. Elsevier, Amsterdam, pp. 349~379.
Pickering, A.D., T.G. Pottinger, J. Carragher and J.P.
Sumpter. 1987. The effects of acute and chronic stress on the levels of reproductive hormones in plasma of mature male brown trout, Salmo trutta L. Gen. Comp.
Endocrinol., 68 : 249~259.
Pickford, G.E., P.K.T. Pang, E. weinstein, J. Torreti, E.
Hendler and F.H. Epstein. 1970. The response of hypo- physectomized cyprinodont, Fundulus heteroclitus, to replacement therapy with cortisol: effects on blood serum and sodium-potassium activated adenosine tri- phosphatase in the gills, kidney and intestinal mucosa.
Gen. Comp. Endocrinol., 14 : 524~534.
Rotlland, J. and L. Tort. 1997. Cortisol and glucose res- ponses after acute stress by net handling in the sparid red porgy previously subjected to crowding stress. J.
Fish Biol., 51 : 21~28.
Rottland, J., P.H. Balm, J. Perez-Sanchez, S.E. Wendelaar -Bonga and L. Tort. 2001. Pituitary and interrenal function in gilthead seabream (Sparus aurata L., Tele- ostei) after handling and confinement stress. Gen.
Comp. Endocrinol., 121 : 333~342.
Ruane, N.M., S.E. Wenderlaar-Bonga and P.H. Balm.
1999. Differences between rainbow trout and brown trout in the regulation of the pituitary-interrenal axis and physiological performance during confinement.
Gen. Comp. Endocrinol., 115 : 210~219.
Sampaio, LA. and A. Bianchini. 2002. Salinity effects on osmoregulation and growth of the euryhaline flounder Paralichthys orbignyanus. J. Exp. Mar. Biol. Ecol., 269 : 187~196.
Schreck, C.B. 1981. Stress and compensation in teleostean fishes: responses to social and physical factors. In: Pick- ering, A.D. Ed., Stress in Fish. Academic Press, London, pp. 295~321.
Specker, C.B., C.S. Bradford, M.S. Fitzpatrick and R. Pati- no. 1989. Regulation of the interrenal of fishes: Non- classical control mechanism. Fish Physiol. Biochem., 7 :
259~265.
Sumpter, J.P., T.G. Pottinger, M. Rand-Weaver and P.M.
Campbell. 1994. The wide-ranging effects of stress on fish. In: Davey, K.G., R.E. Peter, S.S. Tobe (Eds.), Per- spectives in Comparative Endocrinology. National Res- earch Council of Canada, Ottawa, pp. 535~538.
Sunyer, J.O., E. Gomez, V. Navarro, J. Quesada and L.
Tort. 1995. Physiological response and depression of humoral components of the immune system in gilthead sea-bream (Sparus aurata) following daily acute stress.
Can. J. Fish. Aquat. Sci., 52 : 2339~2346.
Tsuzuki, M.Y., K. Ogawa, C.A. Strussmann, M. Maita and F. Takashima. 2001. Physiological responses during stress and subsequent recovery at different salinities in adult pejerrey Odontesthes bonariensis. Aquaculture, 200 : 349~362.
Wedemeyer, G.A. and W.T Yasutake. 1977. Clinical meth- ods for the assessment of the effects of environmental stress on fish health. U.S. Fish and Wildlife Service Technical Paper, 89 : 18.
Wendelaar Bonga, S.E. 1997. The stress response in fish.
Physiol., Rev. 77 : 591~625.
, 崎久 , 崎 . 1986. 鑑.
, 京, 361pp.
Received : August 29, 2003 Accepted : November 24, 2003