Cilostazol Promotes the Migration of Brain Microvascular Endothelial Cells
Sae-Won Lee1,2*, Jung Hwa Park1,2 and Hwa Kyoung Shin1,2,3*
1Korean Medical Science Research Center for Healthy-Aging, Pusan National University, Yangsan, Gyeongnam 626-870, Korea
2Department of Korean Medical Science, School of Korean Medicine, Pusan National University, Yangsan, Gyeongnam 626-870, Korea
3Division of Meridian and Structural Medicine, School of Korean Medicine, Pusan National University, Yangsan, Gyeongnam 626-870, Korea
Received September 1, 2016 /Revised November 2, 2016 /Accepted November 3, 2016
Cilostazol is known to be a selective inhibitor of phosphodiesterase III and is generally used to treat stroke. Our previous findings showed that cilostazol enhanced capillary density through angiogenesis after focal cerebral ischemia. Angiogenesis is an important physiological process for promoting re- vascularization to overcome tissue ischemia. It is a multistep process consisting of endothelial cell pro- liferation, migration, and tubular structure formation. Here, we examined the modulatory effect of cil- ostazol at each step of the angiogenic mechanism by using human brain microvascular endothelial cells (HBMECs). We found that cilostazol increased the migration of HBMECs in a dose-dependent manner. However, it did not enhance HBMEC proliferation and capillary-like tube formation. We used a cDNA microarray to analyze the mechanisms of cilostazol in cell migration. We picked five candidate genes that were potentially related to cell migration, and we confirmed the gene expression levels by real-time PCR. The genes phosphoserine aminotransferase 1 (PSAT1) and CCAAT/enhancer binding protein β (C/EBPβ) were up-regulated. The genes tissue factor pathway inhibitor 2 (TFPI2), retinoic acid receptor responder 1 (RARRES1), and RARRES3 were down-regulated. Our observations suggest that cilostazol can promote angiogenesis by promoting endothelial migration. Understanding the cilostazol-modulated regulatory mechanisms in brain endothelial cells may help stimulate blood vessel formation for the treatment of ischemic diseases.
Key words : Brain microvascular endothelial cell, expression microarray, ischemic disease, motility, therapeutic angiogenesis
*Corresponding authors
*Tel : +82-51-510-8476, Fax : +82-51-510-8437
*E-mail : [email protected] (Hwa Kyoung Shin)
*Tel : +82-51-510-8434, Fax : +82-51-510-8437
*E-mail : [email protected] (Sae-Won Lee)
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2016 Vol. 26. No. 12. 1367~1375 DOI : http://dx.doi.org/10.5352/JLS.2016.26.12.1367
Introduction
Angiogenesis is the formation of new blood vessels from pre-existing vessels, and is a multistep process consisting of endothelial cell proliferation, migration, and tubular struc- ture formation [1, 4]. When quiescent endothelial cells are activated by various stimuli, activated endothelial cells de- grade extracellular matrix, proliferate, and migrate to the site of stimuli. Angiogenesis is completed by the formation and organization of a new lumen, namely, in a tube structure.
Angiogenesis is involved in embryonic vasculature for- mation and is a central process in cancer progression, which means that tumor growth can be effectively inhibited by an-
ti-angiogenic therapy [1, 5]. Conversely, angiogenesis is also an important physiological process for promoting revascula- rization to overcome tissue ischemia. Vascular endothelial growth factor (VEGF) restores ischemic renal injury through angiogenesis [20]. Pioglitazone promotes blood flow recov- ery and capillary density through VEGF upregulation [2].
Administration of plant extract in cases of ischemic stroke reduced infarct damage and enhanced angiogenesis via the angiogenic factors VEGF and angiopoietin-1 and 2 [6]. In addition, the transplantation of bone marrow mononuclear cells into ischemic myocardium augmented collateral vessel formation and improved functional outcome [11]. Implanta- tion of peripheral mononuclear cells into ischemic hindlimb prevented limb amputation by inducing neovascularization [35]. Therefore, therapeutic angiogenesis holds promise for treatment of ischemic diseases.
Cilostazol, a selective type III phosphodiesterase inhibitor, is known to be an anti-platelet agent with vasodilatory activ- ity, and to increase intracellular concentrations of cyclic ad- enosine monophosphate (cAMP) [11]. In addition to these vasodilator and anti-platelet effects, cilostazol also possesses anti-inflammatory and antioxidant properties [15, 26]. Cilo-
stazol showed protective effects against cerebral ischemic in- jury via inhibition of apoptotic and oxidative cell death [15].
Interestingly, cilostazol possesses neovascularization effects.
Cilostazol increased the production of hepatocyte growth factor in vascular smooth muscle cells, which enhanced an- giogenesis in hind limb ischemia [29]. In addition, increased vasculature post-cilostazol administration was observed in mdx dystrophic skeletal muscle [10]. Cilostazol promoted endothelial nitric oxide production in human aortic endothe- lial cells, as well as endothelial tube formation [9]. Cilostazol reduced infarct volume and preserved motor and cognitive function in a rat stroke model. These effects were mediated by promoting angiogenesis through pericyte proliferation [24]. Cilostazol has been reported to promote neovasculari- zation and the expression of angiogenic factors in the hippo- campus in a mouse forebrain ischemia model [30]. Cilostazol also enhanced integrin-dependent homing of endothelial progenitor cells to sites of ischemia in a forebrain ischemia model [14]. Herein, we investigated which angiogenic mech- anisms are regulated by cilostazol by using human brain mi- crovascular endothelial cells.
Materials and Methods
Cell culture and drugs
Human brain microvascular endothelial cells (HBMECs;
passages 5-7, ACBRI) were cultured in complete medium [M199 medium containing 20% fetal bovine serum (FBS;
Gibco), basic fibroblast growth factor (bFGF; 3 ng/ml;
Millipore), heparin (10 U/ml) and 1% penicillin/streptomy- cin (both from Gibco) [17]]. Cilostazol (donation from Otsuka Pharmaceutical Co. Ltd) was dissolved in dimethyl sulfoxide (10 mM stock solution).
In vitro tube formation assay
Formation of capillary-like tubular structure was analyzed by tube formation assay. Briefly, growth factor-reduced Matrigel (200 μl, BD Biosciences) was placed into 24-well culture plates and polymerized for 30 min at 37°C. HBMECs (1×104 cells/well) were seeded onto Matrigel-polymer and incubated in M199 containing 1% FBS with or without cil- ostazol (30 μM). Every hour up to 26 hr, the tubular structures were photographed with an Olympus TH4-200 microscope.
BrdU incorporation assay
Cell proliferation was measured with a 5-bromo-2'-deoxy-
uridine (BrdU) incorporation assay. HBMECs (3,000 cells/
well) were seeded in 96-well culture plates and cultured in a complete medium for 24 hr. The cells were then treated with cilostazol in basal medium (M199 medium containing 1% FBS) for 72 hr. Cells were labeled with BrdU and in- cubated with anti-BrdU antibody conjugated with perox- idase according to the instructions for the Cell Proliferation ELISA, BrdU (calorimetric) (Roche). After removing anti- body conjugate and washing, the substrate solution was added for 20 min and the reaction was stopped by adding a stop solution. The absorbance at 450 nm was measured (SpectraMax 190 microplate reader, Molecular device).
Cell migration assay
HBMECs were seeded on 60-mm culture dishes and cul- tured up to 95% confluence. After scratching with a razor blade 5-mm in width, the cultures were washed with se- rum-free medium twice and further incubated in M199 me- dia containing 1% FBS and 1 mM thymidine. Appropriate concentrations of cilostazol were added to HBMECs, and mi- gration was allowed for 16 hr. Cells were rinsed with se- rum-free medium, fixed with absolute methanol for 2 min, and stained by Giemsa’s staining solution (Kanto). Cells were photographed with an Olympus TH4-200 microscope.
Migration activity was quantitated by counting the number of cells that moved beyond the reference line (the injury line by the razor blade).
RNA preparation, labeling, and microarray analysis Total RNA from HBMECs was extracted using Trizol (Invitrogen Life Technologies, Carlsbad, USA) according to the manufacturers’ protocol. For quality control, RNA purity and integrity were evaluated using denaturing gel electro- phoresis, as well as analysis of the OD 260/280 ratio and using an Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, USA). Total RNA was amplified and purified us- ing the Ambion Illumina RNA amplification kit (Ambion, Austin, USA) to yield biotinylated cRNA according to the manufacturer’s instructions. Labeled cRNA samples (750 ng) were hybridized to each Illumina Human HT-12 expression (V4) bead array for 16-18 hr at 58°C according to the manu- facturer's instructions (Illumina, Inc., San Diego, USA).
Detection of array signal was carried out using Amersham fluorolink streptavidin-Cy3 (GE Healthcare Bio-Sciences, Little Chalfont, UK) following the bead array manual.
Arrays were scanned with an Illumina Bead Array Reader
Table 1. Sequences of primers for real-time PCR
Gene name Primer sequence Size (bp) Gene Bank ID
PSAT1 (Forward) 5’-TGCCCAGAAGAATGTTGGCT-3’
(Reverse) 5’-TCCAGAACCAAGCCCATGAC-3’ 177 NM_058179.3
C/EBPβ (Forward) 5’-CGACGAGTACAAGATCCGGC-3’
(Reverse) 5’-TGCTTGAACAAGTTCCGCAG-3’ 186 NM_005194.3
TFPI2 (Forward) 5’-GCCAACAGGAAATAACGCGG-3’
(Reverse) 5’-AGAAATTGTTGGCGTTGCCC-3’ 149 NM_006528.3
RARRES1 (Forward) 5’-GCGCTACAACCCAGAGTCTT-3’
(Reverse) 5’-TCGATGAGCCGTGTACAAGTT-3’ 126 NM_206963.1
RARRES3 (Forward) 5’-TCTGGCTCCTCCAAGTGAGT-3’
(Reverse) 5’-CCAACCATCTCCTTCGCAGA-3’ 198 NM_004585.2
confocal scanner according to the manufacturer's instructions.
Array data export, processing, and analysis were performed using Illumina BeadStudio v3.1.3 (Gene Expression Module v3.3.8). All data analysis and visualization of differentially expressed genes was conducted using GeneSpring 7.3 soft- ware (Agilent Technologies., Inc.).
Verification of gene expression with real-time PCR The Moloney Murine Leukemia Virus-reverse tran- scriptase (Promega, Madison, WI, USA) was used to produce cDNA from 2 μg of total RNA according to the manu- facturer’s recommendations. Real-time PCR was performed using a Rotor-Gene Q real-time PCR system (Qiagen, Düsseldorf, Hilden, Germany) with SYBR Green PCR Master Mix (Qiagen, Düsseldorf, Hilden, Germany) using the pri- mers listed in Table 1. The results were normalized to 18S rRNA gene expression. All experiments were performed in triplicate and repeated at least three times, and threshold cycles (Ct) were used to quantify the mRNA expression of the target genes.
Western blot analysis
Total protein from HBMECs was isolated as standard techniques, separated by 12% sodium dodecyl sulfate-poly- acrylamide gel electrophoresis (SDS-PAGE), and transferred onto a nitrocellulose membrane (Amersham Biosciences, Piscataway, NJ). Primary antibody against PSAT1 (Thermo Fisher scientific Inc., Waltham, USA) was applied to nitro- cellulose membrane, followed by secondary antibody con- jugated with horseradish peroxidase. Anti-α-tubulin anti- body (Sigma, St. Louis, USA) was used as an internal control.
The intensity of chemiluminescence was measured using an ImageQuant LAS 4000 apparatus (GE Healthcare Life Sciences, Uppsala, Sweden). Band intensity was quantified
using ImageJ (NIH).
Statics
All data are expressed as the mean ± the standard devia- tions (SD). The statistical differences between the groups were compared using the unpaired t-test or the one-way analysis of variance (ANOVA). P-values≤0.05 were considered to indicate statistically significant results.
Results
Cilostazol increased brain endothelial migration Cilostazol has been known to enhance neovascularization after focal ischemia [30]. We observed the effect of cilostazol at each step of the angiogenic mechanism using human brain microvascular endothelial cells (HBMECs). We first con- ducted a wound migration assay to analyze endothelial mo- tility in response to cilostazol (Fig. 1). Treatment with cil- ostazol significantly increased HBMEC migration compared to the control basal medium group in a dose-dependent manner (Fig. 1B).
Endothelial proliferation and capillary-like tube formation was not affected by cilostazol
We next analyzed the effect of cilostazol on HBMEC pro- liferation (Fig. 2A). In contrast to the migration assay, cil- ostazol did not enhance HBMEC proliferation compared to basal medium alone. HBMECs cultured in complete medium were used as a positive control, and they showed enhanced proliferation (Fig. 2A). We then tested if cilostazol could stimulate vessel-like tube formation in HBMECs (Fig. 2B).
Tube formation capacity was not different between cil- ostazol-treated and basal medium-treated cells.
A
B
Fig. 1. Cilostazol promoted HBMEC migration. (A) A HBMEC migration assay was performed 16 hr after cilostazol treatment.
After Giemsa’s staining, cells were photographed (Magnification, x400). (B) A quantitative graph of three independent experi- ments with duplicates is shown. **p<0.01 and ***p<0.001 compared to HBMECs cultured in control basal medium.
A
B
Fig. 2. Cilostazol did not affect the proliferation and tube formation of brain endothelial cells. (A) HBMEC proliferation was as- sessed by a BrdU incorporation assay at 72 hr after cilostazol treatment (n=3,
**p<0.01). Complete medium: M199 me- dium containing 20% FBS and growth factors; basal medium: M199 medium containing 1% FBS. (B) Cilostazol (30 μm) did not affect the formation of capil- lary-like tube networks. Magnification:
×400.
Table 2. Selected genes by us involving known migration-related genes
Gene name Description Synonyms Differential expressiona
PSAT1 Homo sapiens phosphoserine
aminotransferase 1 (PSAT1) MGC1460; PSA; EPIP; PSAT Up-regulated
C/EBPβ Homo sapiens CCAAT/enhancer binding proteinβ (C/EBPβ)
CRP2; TCF5; LAP; IL6DBP;
C/EBP-beta; NF-IL6; MGC32080 Up-regulated TFPI2 Homo sapiens tissue factor pathway
inhibitor 2 (TFPI2) TFPI-2; PP5; FLJ21164 Down-regulated
RARRES1 Homo sapiens retinoic acid receptor
responder (tazarotene induced) 1 (RARRES1) TIG1 Down-regulated
RARRES3 Homo sapiens retinoic acid receptor
responder (tazarotene induced) 3 (RARRES3) MGC8906; HRASLS4; TIG3; RIG1 Down-regulated
aBetween cilostazol (30 μM)-treated HBMECs and control HBMECs as described.
Gene regulation by cilostazol in brain endothelial cells
A microarray analysis was performed for control HBMECs and cilostazol-treated HBMECs to investigate the genes in- fluenced by cilostazol. Compared with control HBMEC cells, there were 107 genes in the expression profile changed (>1.5-fold DEGs) following cilostazol treatment, with 15 genes up-regulated and 92 genes down-regulated. We searched the roles of these genes in previous reports and selected 5 candidate genes that were known to be possibly involved in cell migration (Table 2). We confirmed the candi- date gene expression of 2 up-regulated genes (PSAT1, C/
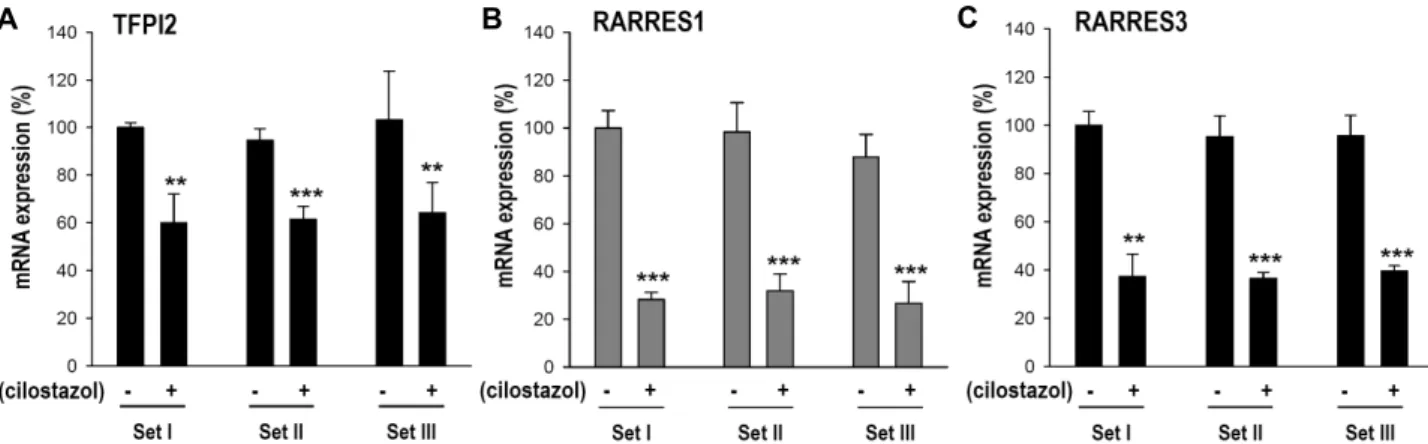
EBPβ) and 3 down-regulated genes (TFPI2, RARRES1 and RARRES3) by real-time PCR (Fig. 3 and Fig. 4). In addition, PSAT1 expression was confirmed by western blot analysis, which was upregulated by cilostazol treatment (Fig. 3C).
Discussion
Endothelial migration is one of the most intensively stud- ied processes in angiogenesis. The migration process is regu- lated by multiple signaling pathways that can act in a differ- ent manner on different blood vessels, and this signaling selectivity is thought to contribute to the modulation of the simultaneous formation of separate blood vessels [31, 33].
We found that cilostazol treatment stimulated brain endo- thelial motility (Fig. 1), and several genes known to be asso- ciated with cell migration were modulated by cilostazol:
PSAT1 and C/EBPβ were up-regulated and TFPI2, RARRES1 and RARRES3 were down-regulated (Table 2, Fig. 3, Fig. 4).
Phosphoserine aminotransferase 1 (PSAT1) is an enzyme involved in serine biosynthesis. PSAT1 has some important metabolic functions such as PSAT1 deficiency results in in-
tractable seizures and acquired microcephaly [8], and re- duced expression in diabetic mice [36]. However, there was no previous information regarding PSAT1 expression and function in endothelial migration. Increased expression of PSAT1 has been reported to be associated with cancer meta- stasis [19, 27, 34]. PSAT1 overexpression promoted esoph- ageal squamous cell carcinoma metastasis, and PSAT1 stim- ulates the expression of snail, which has been reported as a key regulator of cell migration and epithelial-mesenchymal transition [13, 19]. In addition, PSAT1 inhibition reduced the invasion and metastasis of cancer cells [34]. From these pre- vious reports, we assumed that cilostazol-mediated PSAT1 up-regulation may have a role in endothelial migration.
Expression of CCAAT/enhancer binding protein β (C/
EBPβ) increases in cilostazol-treated HBMECs (Fig. 3). C/
EBPβ is a member of a transcription factor family that takes part in the cell migration process [3, 12, 18]. Induction of C/EBPβ in muscle stem cells enhanced their migration and contributed to muscle repair in dystrophic muscle [12].
C/EBPβ is an important mediator in endothelial migration during pathological angiogenesis [18]. In addition, hep- atocyte growth factor-mediated motility upregulation of hu- man melanocyte depended on C/EBPβ [3]. Therefore, the upregulation of C/EBPb by cilostazol in HBMECs may play an important role in endothelial migration.
Among the down-regulated genes by cilostazol (Fig. 4), tissue factor pathway inhibitor 2 (TFPI2) has been well char- acterized to be involved in tumor angiogenesis and pro- gression. Reduced TFPI2 expression correlated with angio- genesis in pancreatic carcinomas [37], TFPI2 knock-down promoted the migration of glioma cells [7], and TFPI2 over- expression inhibited the migration of trophoblast cells [38].
In addition, the overexpression of TFPI2 decreased matrix
A
B
C
Fig. 3. Quantitative real-time PCR confirmation of the up-regulated candidate genes by cilostazol treatment in HBMECs. Real-time PCR analysis of (A) phosphoserine aminotransferase 1 (PSAT1) mRNA and (B) CCAAT/enhancer binding protein β (C/EBPβ) mRNA. RNAs from cilostazol (30 μM)-treated HBMECs and control HBMECs were used. Three independent experiments with triplicates were performed. **p<0.01, ***p<0.001. (C) Cilostazol upregulated the PSAT1 protein level. Total protein (20 μg) was subjected to 12% SDS-PAGE, followed by western blotting using anti-PSAT1. Quantification graphs (n=3, ***p<0.001).
A B C
Fig. 4. Quantitative real-time PCR confirmation of the down-regulated candidate genes by cilostazol treatment in HBMECs. Real-time PCR analysis of (A) tissue factor pathway inhibitor 2 (TFPI2), (B) retinoic acid receptor responder 1 (RARRES1) and (C) retinoic acid receptor responder 3 (RARRES3) mRNA. RNAs from cilostazol (30 μM)-treated HBMECs and control HBMECs were used. Three independent experiments with triplicates were performed. **p<0.01, ***p<0.001.
metalloproteinases (MMPs), which play critical roles in the degradation of extracellular matrix, and in cell migration during metastasis [16, 25]. Another down-regulated gene by cilostazol were retinoic acid receptor responder 1 (RARRES1)
and RARRES3 (Fig. 4). RARRES1 was initially identified as a novel retinoic acid receptor regulated gene [22], and has been reported as a potential tumor suppressor gene down- regulated in tumors [23, 28]. Inhibition of RARRES1 in-
creased prostate cancer cell invasion [23]. RARRES3 has been reported to associate with metastasis, suppressing the meta- stasis of colorectal and breast cancers [21, 32]. Although there is no published data of RARRES1 and RARRES3 in endothelial migration, reports about how reduction of RARRES1 or RARRES3 increased tumor metastasis suggest that reduction of RARRES1 or RARRES3 by cilostazol may be one of the regulatory mechanisms for HBMEC motility stimulation by cilostazol.
Taken together, the expression of the PSAT1, C/EBPβ, TFPI2, RARRES1, and RARRES3 genes modulated by cil- ostazol treatment in HBMECs may result in increased brain endothelial cell migration ability, thus promoting the angio- genesis process. Thus, the clarification of the cilostazol- modulated molecular mechanisms involving these genes will be helpful for angiogenesis-based therapeutics for the treatment of ischemic diseases.
Acknowledgement
This work was supported by a 2-Year Research Grant of Pusan National University.
References
1. Betz, C., Lenard, A., Belting, H. G. and Affolter, M. 2016.
Cell behaviors and dynamics during angiogenesis. Develop- ment 143, 2249-2260.
2. Biscetti, F., Straface, G., Arena, V., Stigliano, E., Pecorini, G., Rizzo, P., De Angelis, G., Iuliano, L., Ghirlanda, G. and Flex, A. 2009. Pioglitazone enhances collateral blood flow in is- chemic hindlimb of diabetic mice through an Akt-depend- ent VEGF-mediated mechanism, regardless of PPARgamma stimulation. Cardiovasc. Diabetol. 8, 49.
3. Damm, S., Koefinger, P., Stefan, M., Wels, C., Mehes, G., Richtig, E., Kerl, H., Otte, M. and Schaider, H. 2010. HGF- promoted motility in primary human melanocytes depends on CD44v6 regulated via NF-kappa B, Egr-1, and C/EBP- beta. J. Invest. Dermatol. 130, 1893-1903.
4. Dimmeler, S. and Zeiher, A. M. 2000. Endothelial cell apop- tosis in angiogenesis and vessel regression. Circ. Res. 87, 434-439.
5. Folkman, J. 1995. Seminars in Medicine of the Beth Israel Hospital, Boston. Clinical applications of research on angio- genesis. N. Engl. J. Med. 333, 1757-1763.
6. Gandin, C., Widmann, C., Lazdunski, M. and Heurteaux, C. 2016. MLC901 favors angiogenesis and associated recov- ery after ischemic stroke in mice. Cerebrovasc. Dis. 42, 139-154.
7. Gessler, F., Voss, V., Seifert, V., Gerlach, R. and Kogel, D.
2011. Knockdown of TFPI-2 promotes migration and in- vasion of glioma cells. Neurosci. Lett. 497, 49-54.
8. Hart, C. E., Race, V., Achouri, Y., Wiame, E., Sharrard, M., Olpin, S. E., Watkinson, J., Bonham, J. R., Jaeken, J., Matthijs, G. and Van Schaftingen, E. 2007. Phosphoserine amino- transferase deficiency: a novel disorder of the serine biosyn- thesis pathway. Am. J. Hum. Genet. 80, 931-937.
9. Hashimoto, A., Miyakoda, G., Hirose, Y. and Mori, T. 2006.
Activation of endothelial nitric oxide synthase by cilostazol via a cAMP/protein kinase A- and phosphatidylinositol 3-kinase/Akt-dependent mechanism. Atherosclerosis 189, 350-357.
10. Hermes Tde, A., Macedo, A. B., Fogaca, A. R., Moraes, L.
H., de Faria, F. M., Kido, L. A., Cagnon, V. H. and Minatel, E. 2016. Beneficial cilostazol therapeutic effects in mdx dys- trophic skeletal muscle. Clin. Exp. Pharmacol. Physiol. 43, 259- 267.
11. Kamihata, H., Matsubara, H., Nishiue, T., Fujiyama, S., Tsutsumi, Y., Ozono, R., Masaki, H., Mori, Y., Iba, O., Tateishi, E., Kosaki, A., Shintani, S., Murohara, T., Imaizumi, T. and Iwasaka, T. 2001. Implantation of bone marrow mon- onuclear cells into ischemic myocardium enhances collateral perfusion and regional function via side supply of angio- blasts, angiogenic ligands, and cytokines. Circulation 104, 1046-1052.
12. Lala-Tabbert, N., Fu, D. and Wiper-Bergeron, N. 2016.
Induction of CCAAT/enhancer-binding protein beta ex- pression with the phosphodiesterase inhibitor isobutylme- thylxanthine improves myoblast engraftment into dystro- phic muscle. Stem Cells Transl. Med. 5, 500-510.
13. Lamouille, S., Xu, J. and Derynck, R. 2014. Molecular mecha- nisms of epithelial-mesenchymal transition. Nat. Rev. Mol.
Cell Biol. 15, 178-196.
14. Lee, D. H., Lee, H. R., Shin, H. K., Park, S. Y., Hong, K.
W., Kim, E. K., Bae, S. S., Lee, W. S., Rhim, B. Y. and Kim, C. D. 2011. Cilostazol enhances integrin-dependent homing of progenitor cells by activation of cAMP-dependent protein kinase in synergy with Epac1. J. Neurosci. Res. 89, 650-660.
15. Lee, J. H., Park, S. Y., Shin, H. K., Kim, C. D., Lee, W. S.
and Hong, K. W. 2008. Protective effects of cilostazol against transient focal cerebral ischemia and chronic cerebral hypo- perfusion injury. CNS Neurosci. Ther. 14, 143-152.
16. Lee, S. W., Jung, K. H., Jeong, C. H., Seo, J. H., Yoon, D.
K., Suh, J. K., Kim, K. W. and Kim, W. J. 2011. Inhibition of endothelial cell migration through the downregulation of MMP-9 by A-kinase anchoring protein 12. Mol. Med. Rep.
4, 145-149.
17. Lee, S. W., Won, J. Y., Kim, W. J., Lee, J., Kim, K. H., Youn, S. W., Kim, J. Y., Lee, E. J., Kim, Y. J., Kim, K. W. and Kim, H. S. 2013. Snail as a potential target molecule in cardiac fibrosis: paracrine action of endothelial cells on fibroblasts through snail and CTGF axis. Mol. Ther. 21, 1767-1777.
18. Lee, W. J., Lan, K. H., Chou, C. T., Yi, Y. C., Chen, W. C., Pan, H. C., Peng, Y. C., Wang, K. B., Chen, Y. C., Chao, T. H., Tien, H. R., Sheu, W. H. and Sheu, M. L. 2013. Tpl2 inhibitors thwart endothelial cell function in angiogenesis
and peritoneal dissemination. Neoplasia 15, 1036-1048.
19. Liu, B., Jia, Y., Cao, Y., Wu, S., Jiang, H., Sun, X., Ma, J., Yin, X., Mao, A. and Shang, M. 2016. Overexpression of Phosphoserine Aminotransferase 1 (PSAT1) predicts poor prognosis and associates with tumor progression in human esophageal squamous cell carcinoma. Cell Physiol. Biochem.
39, 395-406.
20. Logue, O. C., McGowan, J. W., George, E. M. and Bidwell, G. L. 3rd. 2016. Therapeutic angiogenesis by vascular endo- thelial growth factor supplementation for treatment of renal disease. Curr. Opin. Nephrol. Hypertens. 25, 404-409.
21. Morales, M., Arenas, E. J., Urosevic, J., Guiu, M., Fernandez, E., Planet, E., Fenwick, R. B., Fernandez-Ruiz, S., Salvatella, X., Reverter, D., Carracedo, A., Massague, J. and Gomis, R.
R. 2014. RARRES3 suppresses breast cancer lung metastasis by regulating adhesion and differentiation. EMBO Mol. Med.
6, 865-881.
22. Nagpal, S., Patel, S., Asano, A. T., Johnson, A. T., Duvic, M. and Chandraratna, R. A. 1996. Tazarotene-induced gene 1 (TIG1), a novel retinoic acid receptor-responsive gene in skin. J. Invest. Dermatol. 106, 269-274.
23. Oldridge, E. E., Walker, H. F., Stower, M. J., Simms, M. S., Mann, V. M., Collins, A. T., Pellacani, D. and Maitland, N.
J. 2013. Retinoic acid represses invasion and stem cell phe- notype by induction of the metastasis suppressors RARRES1 and LXN. Oncogenesis 2, e45.
24. Omote, Y., Deguchi, K., Kono, S., Liu, N., Liu, W., Kurata, T., Yamashita, T., Ikeda, Y. and Abe, K. 2014. Neurovascular protection of cilostazol in stroke-prone spontaneous hyper- tensive rats associated with angiogenesis and pericyte proliferation. J. Neurosci. Res. 92, 369-374.
25. Pan, J., Ma, D., Sun, F., Liang, W., Liu, R., Shen, W., Wang, H., Ji, Y., Hu, R., Liu, R., Luo, X. and Shi, H. 2013. Over-ex- pression of TFPI-2 promotes atherosclerotic plaque stability by inhibiting MMPs in apoE-/- mice. Int. J. Cardiol. 168, 1691-1697.
26. Park, W. S., Jung, W. K., Lee, D. Y., Moon, C., Yea, S. S., Park, S. G., Seo, S. K., Park, C., Choi, Y. H., Kim, G. Y., Choi, J. S. and Choi, I. W. 2010. Cilostazol protects mice against endotoxin shock and attenuates LPS-induced cyto- kine expression in RAW 264.7 macrophages via MAPK in- hibition and NF-kappaB inactivation: not involved in cAMP mechanisms. Int. Immunopharmacol. 10, 1077-1085.
27. Pollari, S., Kakonen, S. M., Edgren, H., Wolf, M., Kohonen, P., Sara, H., Guise, T., Nees, M. and Kallioniemi, O. 2011.
Enhanced serine production by bone metastatic breast can- cer cells stimulates osteoclastogenesis. Breast Cancer Res.
Treat. 125, 421-430.
28. Sahab, Z. J., Hall, M. D., Zhang, L., Cheema, A. K. and Byers, S. W. 2010. Tumor suppressor RARRES1 regulates DLG2, PP2A, VCP, EB1, and Ankrd26. J. Cancer 1, 14-22.
29. Sanada, F., Kanbara, Y., Taniyama, Y., Otsu, R., Carracedo,
M., Ikeda-Iwabu, Y., Muratsu, J., Sugimoto, K., Yamamoto, K., Rakugi, H. and Morishita, R. 2016. Induction of angio- genesis by a type III phosphodiesterase inhibitor, cilostazol, through activation of peroxisome proliferator-activated re- ceptor-gamma and cAMP pathways in vascular cells.
Arterioscler. Thromb. Vasc. Biol. 36, 545-552.
30. Shin, H. K., Lee, H. R., Lee, D. H., Hong, K. W., Lee, J. H., Park, S. Y., Lee, S. J., Lee, J. S., Lee, W. S., Rhim, B. Y. and Kim, C. D. 2010. Cilostazol enhances neovascularization in the mouse hippocampus after transient forebrain ischemia.
J. Neurosci. Res. 88, 2228-2238.
31. Vanhollebeke, B., Stone, O. A., Bostaille, N., Cho, C., Zhou, Y., Maquet, E., Gauquier, A., Cabochette, P., Fukuhara, S., Mochizuki, N., Nathans, J. and Stainier, D. Y. 2015. Tip cell- specific requirement for an atypical Gpr124- and Reck-de- pendent Wnt/beta-catenin pathway during brain angio- genesis. Elife 4, e06489.
32. Wang, Z., Wang, L., Hu, J., Fan, R., Zhou, J., Wang, L. and Zhong, J. 2015. RARRES3 suppressed metastasis through suppression of MTDH to regulate epithelial-mesenchymal transition in colorectal cancer. Am. J. Cancer Res. 5, 1988- 1999.
33. Wiley, D. M., Kim, J. D., Hao, J., Hong, C. C., Bautch, V.
L. and Jin, S. W. 2011. Distinct signalling pathways regulate sprouting angiogenesis from the dorsal aorta and the axial vein. Nat. Cell Biol. 13, 686-692.
34. Yan, S., Jiang, H., Fang, S., Yin, F., Wang, Z., Jia, Y., Sun, X., Wu, S., Jiang, T. and Mao, A. 2015. MicroRNA-340 Inhibits esophageal cancer cell growth and invasion by tar- geting phosphoserine aminotransferase 1. Cell Physiol.
Biochem. 37, 375-386.
35. Youn, S. W., Lee, S. W., Lee, J., Jeong, H. K., Suh, J. W., Yoon, C. H., Kang, H. J., Kim, H. Z., Koh, G. Y., Oh, B.
H., Park, Y. B. and Kim, H. S. 2011. COMP-Ang1 stimulates HIF-1alpha-mediated SDF-1 overexpression and recovers is- chemic injury through BM-derived progenitor cell recruit- ment. Blood 117, 4376-4386.
36. Yu, J., Xiao, F., Guo, Y., Deng, J., Liu, B., Zhang, Q., Li, K., Wang, C., Chen, S. and Guo, F. 2015. Hepatic phosphos- erine aminotransferase 1 regulates insulin sensitivity in mice via tribbles homolog 3. Diabetes 64, 1591-1602.
37. Zhai, L. L., Wu, Y., Huang, D. W. and Tang, Z. G. 2015.
Increased matrix metalloproteinase-2 expression and re- duced tissue factor pathway inhibitor-2 expression correlate with angiogenesis and early postoperative recurrence of pancreatic carcinoma. Am. J. Transl. Res. 7, 2412-2422.
38. Zhou, Q., Xiong, Y., Chen, Y., Du, Y., Zhang, J., Mu, J., Guo, Q., Wang, H., Ma, D. and Li, X. 2012. Effects of tissue factor pathway inhibitor-2 expression on biological behavior of BeWo and JEG-3 cell lines. Clin. Appl. Thromb. Hemost. 18, 526-533.
초록:Cilostazol에 의한 뇌혈관내피세포의 세포이동 증진 효과연구
이세원1,2*․박정화1,2․신화경1,2,3*
(1부산대학교 건강노화 한의과학 연구센터, 2부산대학교 한의학전문대학원 한의과학과, 3부산대학교 한의학전문대
학원 경락구조의학부)
Cilostazol은 phosphodiesterase III의 선택적 저해제로 알려져 있으며, 뇌졸중 치료에 일반적으로 사용되고 있 다. Cilostazol을 처리한 경우, 국소 뇌허혈이 발생한 후에 혈관신생을 통해서 혈관형성이 향상된다는 것을 본 연 구자들이 발표하였다. 혈관신생은 조직의 허혈상태를 극복하기 위해서 혈관재생을 촉진하는 중요한 과정으로써, 혈관내피세포의 증식, 이동, 모세관구조 형성의 다단계 과정으로 구성되어 있다. 이에 본 연구에서는 인간 뇌혈관 내피세포를 이용하여 cilostazol이 혈관신생의 각 단계들에 어떤 영향을 미치는지 조사하였다. Cilostazol은 농도 의존적으로 뇌혈관내피세포의 이동성을 촉진하였으나, 뇌혈관내피세포의 증식과 모세관구조 형성에는 영향을 미 치지 않았다. Cilostazol이 세포이동을 조절하는 기전을 분석하기 위해서 cDNA microarray를 수행하였고, 세포이 동에 관련성이 있는 5종의 후보 유전자들을 선택하여 real-time PCR을 통해 해당 유전자의 발현을 검증하였다.
Cilostazol에 의해서 발현양이 조절되는 유전자들로써, phosphoserine aminotransferase 1 (PSAT1)와 CCAAT/en- hancer binding protein β (C/EBP β)은 발현이 증가하였고, tissue factor pathway inhibitor 2 (TFPI2), retinoic acid receptor responder 1 (RARRES1), RARRES3는 발현이 감소하였다. 이상의 결과를 통해서 cilostazol이 혈관 내피세포의 이동을 촉진하여 혈관신생을 향상시킬 수 있음을 제안할 수 있으며, 뇌혈관내피세포에 대한 cilostazol 의 조절기전에 대해서 더욱 상세히 규명을 한다면 혈관형성을 통하여 허혈성 질환을 치료할 수 있는 유용한 정보 가 될 것으로 기대한다.