1
서 론
유방암의 발생, 성장, 및 전이과정에는 세포의 생존 또는 죽음에 관련하는 여러 가지 인자가 복합적으로 작용한 다.(1) 이 과정에는 종양세포의 발생 과정에서 숙주의 면역 체계를 회피하기 위한 기전, 성장과정에서 필요한 혈관 형 성인자 및 성장 인자의 분비, 지속적 세포 분열을 위한 신호 전달체계의 변화 등이 포함된다.(2)
스테로이드 성 호르몬인 에스트로겐은 다양한 조직의 발 생과정뿐만 아니라 여러 생리현상의 항상성 유지에 깊은 연관을 가지고 있다. 반면에 에스트로겐은 유방암을 포함 한 여러 가지 여성 종양의 발생 및 진행에도 중요한 역할을 한다고 알려져 있다.(3) 세포에 대한 에스트로겐의 영향은 주로 에스트로겐 수용체 α, 또는 β수용체와의 결합을 통 해 이루어지며, 이들 에스트로겐 수용체 α, 또는 β수용체 는 기본적으로 ligand에 의해 활성화되는 전사 조절 인자라 볼 수 있다.(4-6) 유방조직의 발생 및 유방암에 대한 에스트 로겐의 영향은 이들 조직세포에 존재하는 에스트로겐 수용 체, 특히 ER-α에 의해 일어나며 에스트로겐과 에스트로겐 수용체의 결합은 세포의 증식을 유도하는 여러 신호전달 경로를 활성화시킨다.(7,8) 최근의 연구에 의하면 에스트로 겐은 세포사멸을 억제하는 대표적 유전자인 bcl-2의 promoter에 작용하여 발현을 촉진함으로써 세포사멸을 차 단한다는 사실이 밝혀졌다.(9) 그러나 에스트로겐의 다양한 작용기전 및 기능에 비추어 볼 때 bcl-2 유전자의 발현만으 로는 종양의 발생과정에 관여하는 에스트로겐의 역할을 충 분히 설명하기에는 부족한 점이 많다. 최근에 에스트로겐 수용체와 관련을 가지며 여러 유전자의 전사 조절 인자로 Further studies using breast cancer tissues will be needed.
(J Korean Surg Soc 2005;68:1-8)
Key Words: Breast cancer, Estradiol, E2IG5 중심 단어: 유방암, Estradiol, E2IG5
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
1Singal Kang-Nam Hospital, Departments of Surgery and
2Pathology, Kyung Hee University Hospital, Seoul, Korea
책임저자:홍성화, 서울특별시 동대문구 회기동 1번지 ꂕ 130-702, 경희대학교 의과대학 외과학교실 Tel: 02-958-8264, Fax: 02-966-9366
E-mail: [email protected]
접수일:2004년 6월 5일, 게재승인일:2004년 10월 18일
Estradiol에 의해 유도되는 E2IG5 단백의 유방암 세포 주에 대한 작용
1신갈 강남병원, 경희대학교 의과대학 외과학교실 및 2병리학교실
송정윤1․장연수․고석환․홍성화․오수명․윤 충․박용구2․박재훈2
Effect of Estradiol-induced E2IG5 on Breast Cancer Cell Line
Jeong Yoon Song, M.D.1, Yeon Soo Chang, M.D., Suck Hwan Koh, M.D., Sung Wha Hong, M.D., Soo Myung Oh, M.D., Choong Yoon, M.D., Yong Koo Park, M.D.2 and Jae Hoon Park, M.D.2
Purpose: Estrogens control the development and cell proliferation of various tissues including the normal mam- mary epithelial cells, where they induce the expression of the immediate and delayed hormone-responsive genes. The proliferative effects of estrogen have been attributed to its ability to increase the expression of the key cell cycle regulatory genes responsible for cell cycle progression. How- ever, the regulation of cell proliferation is only one aspect of estrogen function. It has also been well documented that estrogen plays a critical role in the etiology and progression of human breast and gynecological cancers. This tumori- genic effect of estrogen might be associated with its anti- apoptotic activities such as of Bcl-2 induction. The aim of this study was to clarify the role of E2IG5, which is an estrogen-induced downstream effector molecule, in breast cancer cell lines.
Results: This study shows that E2IG5 is a pro-apoptotic protein that is localized to the mitochondrial membrane via two distinct transmembrane domains. When over-expressed, it induces a mitochondrial permeability transition with the re- sultant of release cytochrome c and caspase activation.
However, three out of four breast cancer cell lines lost their estrogen dependence of E2IG5 expression, which suggests the possible involvement of E2IG5 in the development of breast cancer.
Conclusion: These results suggest that breast cancer cells may loose their pro-apoptotic signals and selectively use the proliferative mechanism of estrogen, which drives the normal mammary epithelial cells to transform into cancer cells.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ 작용하는 많은 새로운 단백질들이 발견되었다.(10) 이 중
에스트로겐의 세포 분열과 관련된 부분은 세포 주기 조절 인자의 발현을 통하여 이루어진다고 생각되나 이는 에스트 로겐의 다양한 기능 중 일부에 불과하다고 볼 수 있다.(11) 특히 에스트로겐의 기능 중에서 세포사멸에 관여하는 신호 전달 체계에 관한 연구는 아직 미미한 수준이다.
에스트로겐은 에스트로겐 수용체와의 결합을 통하여 다 양한 유전자의 발현을 조절하는데 2000년 Charpentier등은 에스트로겐에 의해 유도되는 수많은 유전자를 Serial Analy- sis of Gene Expression (SAGE) 방법을 이용하여 밝혀내고 보고한 바 있다.(4) 이 유전자 중에는 estrogen-induced gene 5 (E2IG5, GeneBank Accession No. AF191020)라는 유전자가 포함되어 있다. 본 연구는 에스트로겐이 유방암과 밀접한 관련이 있다는 사실에 착안하여 에스트로겐이 유방암의 발 생, 진행과정에 미치는 영향을 이해하기 위하여 에스트로 겐에 의해 유도되는 유전자인 E2IG5가 유방암 세포주에 미 치는 영향, 특히 세포사멸에 미치는 영향을 중심으로 연구 하였다.
방 법
1) 세포배양
유방암 세포주 Hs 578T, SKBR3, SCC-1395, 및 MDA- MB-435 등은 한국세포주은행(Seoul, Korea)에서 구입하였 으며 10% FBS가 첨가된 RPMI 1640배지를 사용하여 37oC, 5% CO2 배양기에서 배양하였다. 약 70%의 밀도를 보일 때 계대 배양을 시행하였다.
2) RNA추출 및 cDNA제작
유방암 세포주에서 guanidine isothiocyanate-chloroform- isoamyl alcohol을 사용한 single-step method로 RNA를 추출 하였다.(12) 추출한 RNA 5μg을 AMV Reverse Transcriptase (Gibco BRL)를 사용하여 cDNA를 제작하여 염기서열 분석 및 연쇄중합효소 반응에 사용하였다.
3) 유전자 동정 및 분석
E2IG5 유전자 염기서열에 관한 정보는 GeneBank (Acces- sion No. AF191020)에서 얻었으며 sense primer (5'-atgggga gcctcagcggtct-3'), anti-sense primer (5'-ctctgttttggccttcatagct-3') 와 신장조직에서 합성된 human cDNA library (Clontech)를 template로 하여 연쇄중합효소법(PCR)을 이용하여 증폭하 였다. 증폭된 유전자는 Topo TA/His-V5벡터에 넣은 후 T7 primer를 이용하여 염기서열을 확인하였다. 변이형 유전자 는 splicing overlap extension method를 이용하여 제작하였 다.(13)
4) 세포사 측정
세포사의 측정은 trypan blue exclusion method를 이용하거 나, 또는 DAPI 염색을 시행한 후 형광현미경 하에서 분절되 거나 농염화된 염색질을 가지고 있는 핵의 수를 헤아려 판 단하였다.
5) 유전자 주입
유방암 세포주에 대한 유전자 주입은 배양 dish에 60%의 밀도를 보일 때 Transfast Reagent (Promega)를 이용하여 일 시적인 유전자 과 발현을 유도하였다.
6) Caspase 효소 활성 분석
Caspase-3 및 caspase-9의 활성분석은 106개의 세포를 50 μl의 세포용해 용액(50 mM HEPES, pH 7.4, 100 mM NaCl, 0.1% CHAPS, 1 mM DTT, 0.1 mM EDTA)에서 용해시킨 후 5분간 얼음에 방치하였다. 이후 10분간 10,000 g에서 원심 분리한 후 상층액을 회수하였다. 상층액에 Ac-DEVD-pNA 또는 Ac-LEHD-pNA를 첨가한 후 spectrophotometer를 이용 하여 405 nm에서 흡광도를 측정하였다.(14)
7) 항체 합성
Anti-E2IG5 polyclonal antibody (L16)은 KLH가 결합되어 있는 peptide DLKRINGFCTKPQESP 5μg을 토끼에 주사한 후 2주 후 추가 접종하였다. 4주 후 혈청을 분리한 후 Pro- tein A를 이용하여 항체를 분리하였다.
8) 면역조직화학검사
유방암 세포주를 4% paraformaldehyde에 고정한 후 0.1%
Triton X-100으로 세포막에 작은 구명을 뚫었다. 이후 3%
bovine serum albumin으로 항체의 비특이적 결합을 차단한 후 anti-AL40, anti-V5 또는 anti-His 일차항체(Invitrogen, USA)를 붙인 후 다시 FITC, Texas-Red가 붙어있는 이차항 체를 결합시켰다. 관찰은 형광현미경으로 screening 후 con- focal microscope (Bio-Rad)를 사용하여 관찰하였다.
9) 반 정량적 RT-PCR
cDNA 1μl를 template로 하여 GAPDH primer (sense; 5'- ATGTCTCAGAGCAACCGGGAG-3', antisense; 5'-TTTCCG ACTGAAGAGTGAGC-3')각 10 pmoles, 10x PCR amplifi- cation buffer 10μl, 1.25mM dNTP 16μl, H2O 30μl를 넣어 94oC에서 45초간 denaturation, 58oC에서 30초간 annealing, 72oC에서 45초간 polymerization을 36 cycle 시행하였다. 시 료를 2% agarose gel에서 20분간 전기영동한 후 ethidium bromide에 10분간 염색하여 Gel-doc system (Bio-Rad, CA)으 로 정량화했다. 이 결과를 바탕으로 band의 밝기에 따라 template cDNA의 양을 증감하여 상대적으로 cDNA의 양을
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ 동일하게 조절했다. 이후 bax, bak, p53, bid, bcl-2, bcl-XL
primer를 사용하여 각각 35, 36, 38 cycles의 PCR반응을 시행 하여 Gel-doc system (Bio-Rad, CA)으로 정량한 값을 GAP DH에 대한 값으로 나누어 각 유전자의 발현 정도를 정량화 하였다.
10) 면역블롯
3×106개의 세포를 회수한 후 protease inhibitors (Pheny- lmethylsulfonyl fluoride 1 mM, Aprotinin 0.23 unit/ml, Leu- peptin 10μM, Benzamidine 1 mM)를 포함한 protein extrac-
tion buffer (NaCl 0.15M, Tris 10 mM (pH 7.4), EDTA 5 mM, 1% NP-40)에 넣은 후 10분간 얼음 위에 보관하였다. 14,000 xg, 4oC에서 10분간 원심분리한 후 상등액을 취하여 Lowry method로 단백질 양을 정량하였다. 동량(50μg/ lane)의 단백 질을 10% acrylamide gel상에서 전기영동하여 semi-dry transblotter를 이용하여 nitrocellulose (NC) membrane에 blot- ting하였다. NC membrane을 PBS에 1회 세척한 후 Blocking buffer (2% BSA in PBS)에 넣어 4oC에서 overnight하였다.
Anti-AL60 항체와 상온에서 1시간 반응시킨 후 Washing buffer (PBS with 0.2% Tween 20)로 5분씩 3회 세척한 후 이 차 항체(anti-goat IgG conjugated with peroxidase, Vec- tor)(dilution 1:1500)와 상온에서 40분간 반응시킨 후
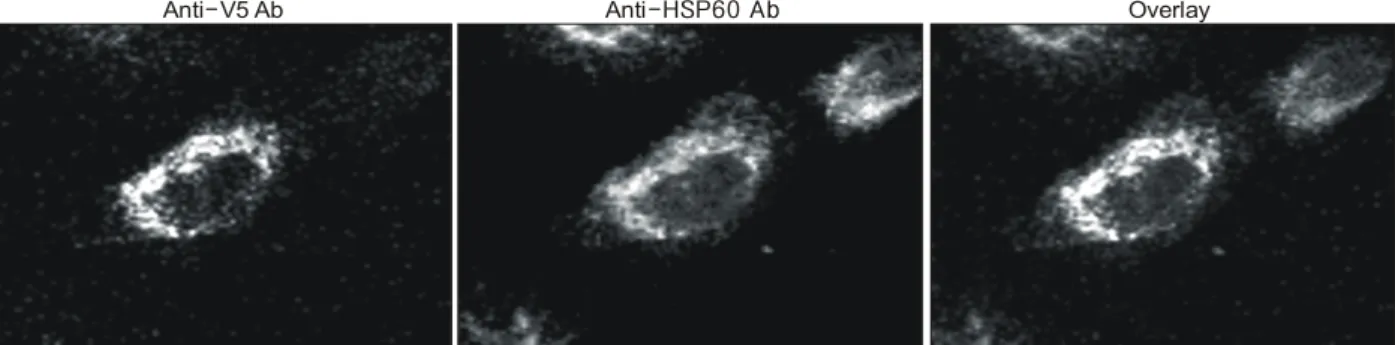
Fig. 3. E2IG5 is a mitochondrial protein. HS 578T breast cancer cells were transiently transfected with pcDNA-E2IG5. At 24 h after transfection, the cells were double stained with mouse anti-V5 antibody conjugated with FITC (E2IG5) and rabbit anti-HSP60 antibody conjugated with Texas-Red (HSP60). The stained images were combined to compare the staining pattern of both proteins (Overlay) and their coincidence was indicated by yellow.
Anti V5 Ab- Anti-HSP60 Ab Overlay
Fig. 1. Estrogen dependency of E2IG5 mRNA expression in Breast cancer cell lines. cDNA was synthesized from the total RNAs extracted from the indicated breast cancer cell lines treated with 10μM estrogen and subjected to RT-PCR analysis. 32 and 26 cycles of amplification were performed for E2IG5 and GAPDH, respectively. Fold increases of E2IG5 expression (mean±S.D.) were presented using den- sitometry. E2IG5 expression level at 0 h was arbitrarily set to 1.
Fig. 2. Estrogen dependency of E2IG5 protein expression in Breast cancer cell lines. 30μg of cell lysates extracted from estrogen-treated breast cancer cell lines was subjected to western blot analysis using polyclonal anti-E2IG5 or β -tubulin antibodies. Fold increases of E2IG5 protein ex- pression was presented. E2IG5 expression level at 0 h was arbitrarily defined as 1.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Washing buffer (PBS with 0.2% Tween 20)로 5분씩 3회 세척
하였다. Membrane을 ECL (Amersham Pharmacia)을 이용하 여 x-ray film에 감광하였다. 감광된 x-ray film을 Densi- tometer (Bio-Rad)를 이용하여 정량화하였다.
결 과
1) 에스트로겐이 유방암 세포주의 E2IG5발현에 미치 는 영향
합성 에스트로겐을 각 세포주에 1μM의 농도로 처리한 후 1, 2, 6, 12, 24 시간 후에 회수하여 RNA와 단백질을 추출 하였다. 반 정량적 RT-PCR과 면역 블롯을 이용하여 E2IG5 의 발현을 비교하였다. 결과 Hs 578T, SKBR3, SCC-1395 세 포주에서는 에스트로겐의 자극에도 불구하고 의미 있는 E2IG5 mRNA의 변화는 관찰되지 않았다. 반면에 MDA- MB-435 세포주는 에스트로겐의 자극에 대하여 E2IG5 mRNA의 발현이 증가됨을 관찰하였다(Fig. 1). 단백질 수준 에서의 발현상의 변화도 mRNA의 변화와 유사한 결과를 얻 었다. Hs 578T, SKBR3, SCC-1395 세포주는 에스트로겐의 자극에 대하여 단백질 수준에서의 변화가 관찰되지 않았으 나 MDA-MB-435 세포주는 에스트로겐의 자극에 대하여 E2IG5 단백의 증가가 관찰되었다(Fig. 2).
2) E2IG5의 유방암 세포 주 내의 분포
E2IG5의 세포 내 발현 분포를 확인하기 위하여 pcDNA- E2IG5를 Transfast Reagent를 이용하여 세포 내로 주입하였 다. 주입 24시간 후에 세포를 회수하여 벡터에 부착되어 있 는 항 V5에 대한 항체를 이용하여 면역조직화학염색을 시 행하였다. 결과 E2IG5는 세포질 내에 과립상으로 분포하여 미토콘드리아에 분포하는 소견을 보였다. 이를 확인하기 위하여 미토콘드리아 기질 단백질인 HSP60에 대한 항체로 면역조직화학적 염색을 이행한 후 V5에는 FITC, HSP60에 는 Texas-Red가 부착된 이차항체를 이용하여 염색한 후 confocal microscope를 이용하여 관찰하였다.(15) FITC와 Texas-Red를 중첩시켰을 때 두 형광 물질이 일치하는 결과 를 얻었다. 이 결과를 통하여 E2IG5는 미토콘드리아에 발 현되는 단백질임을 확인하였다(Fig. 3).
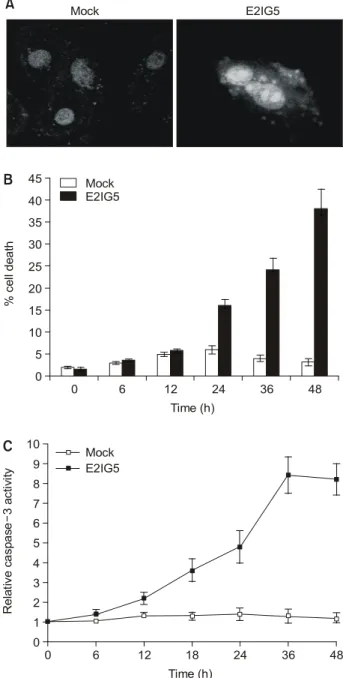
3) E2IG5의 과 발현이 유방암 세포에 미치는 영향 E2IG5가 유방암 세포에 미치는 생물학적 기능을 밝히기 위하여 E2IG5를 Hs 578T 유방암 세포주에 과 발현시킨 후 48시간 후에 세포를 회수한 후 DAPI염색을 시행하여 핵의 형태학적 변화를 관찰하였다. 결과로 분절되거나 농염화 된 염색질을 갖는 핵이 관찰되어 형태학적으로 세포사멸을 시사하는 소견으로 판단하였다(Fig. 4A). 농염화된 염색질 이나 분절된 핵을 갖는 세포의 수를 헤아린 결과 24시간 후 부터 mock vector를 사용한 세포에 비해 의미 있는 차이를
Fig. 4. E2IG5 induces apoptotic cell death with caspase activation.
HS 578T breast cancer cells were transfected with 1μg of mock vector or pcDNA-E2IG5. (A) After 48 h of trans- fection, cells were harvested, stained with DAPI, and visualized using confocal microscope. (B) Cells with frag- mented nuclei or chromatin condensation in transfected cells were counted at indicated time points. From which, percent cell death was obtained. (C) Cells were harvested after indicated times of transfection and caspase assays were performed. Caspase activities of Mock-transfected cells were arbitrarily set to 1. Fold-increase of caspase activity was presented.
Mock E2IG5
A
0 6 12 18 24 36 48
10
Relative caspase3 activity-
Time (h) 0
E2IG5 9
8 7 5 3 6 4
2 1
C Mock
0 6 12 24 36 48
% cell death
Time (h) 0
5 10 15 20 25 30 35 40 45
E2IG5
B Mock
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Fig. 6. E2IG5 induction sensitizes MDA-MB-435 breast cancer cell line to methotrexate. Breast cancer cell lines were pre-treated with 1μM estrogen for 24 h and then incubated with with 10μM methotrexate for indicated times. Cells with fragmented nuclei or chromatin condensation in transfected cells were counted at indicated time points. From which, percent cell death was obtained.
Fig. 5. E2IG5 induces mitochondrial PT in breast cancer cells. HS 578T breast cancer cells were transiently transfected with 1μg of mock vector or pcDNA-E2IG5 (A) in the presence of 20μM CsA (B) or 100μM BA (C). After 24 h of transfection, cells were stained with 400 nM potential-sensitive Mitotracker Red CMXRos and MTP was determined by flow cytometry.
100 101 102 103 104
Counts
0 40 80 120 160 200
A
100 101 102 103 104
Counts
0 40 80 120 160 200
B
100 101 102 103 104
Counts
0 40 80 120 160 200
C
HGTD P-
Mock
HGTD P-
Mock
HGTD P + CsA-
HGTD P- Mock
HGTD P + BA-
Fluorescence intensity
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ 보이기 시작하였다(Fig. 4B). E2IG5가 세포사멸을 유도한다
는 사실을 규명하기 위하여 E2IG5를 과 발현시킨 후 24시 간부터 caspase 효소의 활성도를 측정하였다. 그림에서 볼 수 있듯이 caspase-3의 활성도가 대조군에 비하여 의미 있게 증가되는 것이 관찰되었다(Fig. 4C). 이러한 결과로서 E2IG5는 유방암 세포주에서 세포사멸을 유도하는 유전자 임을 알 수 있다.
4) E2IG5의 세포손상 기전
위의 연구결과에서 E2IG5는 미토콘드리아에 발현되며 세포사멸을 유도하는 유전자임이 확인되었다. 따라서 E2IG5에 의한 세포사멸 과정에서 미토콘드리아를 통한 경 로가 중요한 역할을 하리라 판단되었다. 이를 바탕으로 E2IG5의 과 발현이 미토콘드리아에 미치는 영향을 분석하 였다. 먼저 Hs 578T 유방암 세포주에 E2IG5를 과 발현시킨 후 미토콘드리아 막 전위를 측정하였다. Fig. 5에서 보듯이 E2IG5 과 발현 24시간 후 미토콘드리아 막 전위가 감소되 는 것이 관찰되었다. 그러나 이러한 미토콘드리아 막 전위 의 감소는 다른 경로에 의한 손상 결과 이차적으로 발생될 수 있다. 따라서 이를 구분하기 위하여 미토콘드리아 막 전 위를 소실시키는 permeability transition 의 억제 물질인 cy- closporin A (CsA)와 bonkrekic acid (BA)를 처리하여 막 전위 및 세포사멸 정도를 측정하였다. 결과 CsA 및 BA에 의해 미토콘드리아 막 전위의 감소가 차단되었으며 유방암 세포 주의 세포사멸이 감소됨을 확인하였다(Fig. 5B, C). 이상의 결과는 E2IG5에 의한 세포사멸 과정에서 미토콘드리아가 중요한 경로에 있음을 시사한다.
5) E2IG5의 발현이 Methotrexate에 의한 유방암 세포 사멸에 미치는 영향
이상의 결과로 E2IG5는 유방암 세포주에서 세포사멸을 유도하는 유전자이며 세포주에 따라 에스트로겐에 의해 유 도되는 세포주와 유도되지 않는 세포주가 있음이 확인되었 다. 다음으로 에스트로겐에 의해 E2IG5가 유도된 유방암 세포주와 그렇지 않은 세포주에서 항암제로 사용되는 methotrexate에 대한 감수성의 변화를 분석하였다. 에스트로 겐에 의해 E2IG5의 발현이 유도되지 않는 Hs 578T, SKBR3, SCC-1395 유방암 세포주와 발현이 유도되는 MDA-MB-435 세포주를 대상으로 1μM 에스트로겐에 24시간 노출시킨 후 methotrexate 1μg/ml을 첨가하여 각 시간별로 세포사멸의 정도를 측정하였다. E2IG5가 유도된 MDA-MB-435 세포주 는 methotrexate를 단독 투여한 경우보다 세포사멸이 의미 있게 증가하였으나 E2IG5가 유도되지 않은 Hs 578T, SKBR3, SCC-1395 세포주는 특이한 변화를 관찰할 수 없었 다(Fig. 6). 이상의 결과로 E2IG5의 발현은 유방암 세포주에 서 항암제에 대한 감수성을 증가시킨다는 사실을 확인하였 다.
고 찰
1999년 Shi 등(AF201944)이 human dendritic cell에서 E2IG5를 처음 동정한 후 그 기능에 대해서는 밝혀져 있지 않았으나(16) 본 연구에서 4개의 유방암 세포 주를 대상으 로 과 발현을 통하여 E2IG5의 기능을 분석한 결과 세포사 멸을 유도하는 유전자로 밝혀졌다. 더욱이 이 단백질이 분 포하는 세포 내 소 기관이 미토콘드리아라는 점과 세포사 멸 유도과정에서 permeability transition을 유도한다는 점, 또 한 미토콘드리아와 밀접한 연관성을 갖는 cytochrome c 유 리가 관찰된다는 사실은 본 연구 결과를 뒷받침해 주는 일 치된 결과로 생각할 수 있다.(17-20)
저자는 E2IG5의 발현에 있어 에스트로겐 의존성을 밝히 고자 4개의 유방암 세포 주를 사용하여 에스트로겐 자극 후 RNA 및 단백질 수준에서 E2IG5의 발현을 검증한 결과 일반적으로 알려진 바와는 달리 4개의 유방암 세포 주중에 서 3개의 세포 주는 에스트로겐에 반응을 하지 않았다. 이 는 E2IG5가 세포사멸을 유도하는 유전자라는 사실에 근거 한다면 유방암 세포 주의 세포사멸 효과가 감소할 수 있다 고 추정할 수 있다. 그러나 어떤 기전을 통하여 E2IG5의 발 현이 차단되는지는 추후 연구가 필요하나 RNA와 단백질 수준에서 모두 반응을 보이지 않는 것으로 미루어 post- translational degradation보다는 전사과정에서의 변화로 생각 된다.
에스트로겐은 세포사멸을 유도하는 하위 유전자의 발현 을 조절하는 반면에 세포사멸을 억제하는 유전자의 발현도 조절한다.(21,22) 세포 증식에 관여하는 많은 유전자들은 기 능상 세포 사멸과 밀접한 연관성을 가지고 있다.(23,24) 최 근까지 진행된 많은 연구에서 에스트로겐은 유방암을 포함 한 많은 암에서 세포사멸을 억제한다고 알려져 있다. 항 에 스트로겐은 유방암 세포 주인 MCF-7에서 세포사멸을 유도 하였으며, 또한 에스트로겐을 제거한 nude mouse에서 MCF-7으로 유도된 종괴가 빨리 사라진다는 연구 결과가 있다.(25,26) 이러한 사실들로 미루어, 에스트로겐은 세포사 멸 억제 유전자뿐만 아니라 세포사멸 유도 유전자의 발현 을 유도한다. 그러나 세포사멸을 유도하는 유전자의 유도 는 promoter 등 유전자 발현 조절 기전의 이상으로 발현이 되지 않고 세포사멸을 억제하는 유전자 또는 세포를 증식 시키는 유전자의 발현만 전달됨으로써 궁극적으로 유방암 을 포함한 종양의 발생을 유도한다고 생각된다. 그러나 이 러한 가설의 검증에는 많은 추가적인 연구가 수행되어야 한다. 첫째, 유방암 세포 주에서 에스트로겐에 의한 E2IG5 의 발현이 억제되는 기전을 밝혀야 한다. 본 연구에서 볼 수 있듯이 E2IG5의 유전적 변이는 관찰되지 않은 점으로 미루어 유전적 변이 자체보다는 조절 과정의 변화가 더 가 능성이 높다고 생각된다. 둘째, 유방암 세포주가 아닌 환자
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ 조직에서 E2IG5의 발현을 비교, 분석하여야 한다. 즉 정상
유방조직 및 동일 환자의 유방암 조직을 대상으로 E2IG5의 발현을 비교, 분석하여 유방암의 발생, 진행 과정에서 E2IG5의 의미를 밝혀야 한다.
본 연구의 마지막 결과에서 E2IG5의 발현은 유방암 세포 주의 methotrexate에 대한 세포사멸의 민감도를 증가시켰다.
이는 추후 유방암에 대한 유전자 치료과정에서 E2IG5를 이 용한 treatment modality 개발 가능성을 시사한다고 볼 수 있 다. 그러나 모든 항암제에서 이러한 현상이 관찰된 것은 아 니므로 많은 지속적 연구가 필요한 부분이다.
본 연구 결과 E2IG5는 특정 실험 하 유방암 세포주에 세 포사멸을 유도한다는 사실이 밝혀졌으며 유방암 세포주에 서 E2IG5의 발현이 다른 종양세포에서의 발현보다 감소되 어 있었다. 이러한 사실은 유방암의 발생과정에서 E2IG5의 발현이 억제됨으로써 에스트로겐에 의한 신호과정에서 세 포사멸을 조절하는 신호는 차단되고 종양세포의 증식을 촉 진하는 신호만 선택적으로 전달되어 유방암 발생에 선택적 우위점을 가지게 된다고 볼 수 있다. 추후 각 단계별 유방암 조직을 대상으로 E2IG5의 발현을 분석한다면 유방암의 발 생, 진행과정에서 E2IG5의 역할을 좀더 구체적으로 밝힐 수 있다고 생각한다.
REFERENCES
1) Murphy L, Cherlet T, Lewis A, Banu Y, Watson P. New in- sights into estrogen receptor function in human breast cancer.
Ann Med 2003;35:614-31.
2) Sledge GW Jr, Miller KD. Exploiting the hallmarks of cancer:
the future conquest of breast cancer. Eur J Cancer 2003;39:
1668-75.
3) Pike M, Spicer D, Dahmoush L, Press M. Estrogens, pro- gesterones, normal breast cell proliferation, and breast cancer risk. Epidemiol Rev 1993;15:17-35.
4) Charpentier AH, Bednarek AK, Daniel RL, Hawkins KA, Laflin KJ, Gaddis S, et al. Effects of estrogen on global gene expression: Identification of novel targets of estrogen action.
Cancer Res 2000;60:5977-83.
5) Pace P, Taylor J, Suntharalingam S, Coombes RC, Ali S.
Human estrogen receptor beta binds DNA in a manner similar to and dimerizes with estrogen receptor alpha. J Biol Chem 1997;272:25832-8.
6) Shanmugam M, Krett NL, Maizels ET, Cutler RE Jr, Peters CA, Smith LM, et al. Regulation of protein kinase C delta by estrogen in the MCF-7 human breast cancer cell line. Mol Cell Endocrinol 1999;148:109-18.
7) van der Burg B, van Selm-Miltenburg AJ, de Laat SW, van Zoelen EJ. Direct effects of estrogen on c-fos and c-myc protooncogene expression and cellular proliferation in human breast cancer cells. Mol Cell Endocrinol 1989;64:223-8.
8) Dubik D, Dembinski TC, Shiu RP. Stimulation of c-myc on- cogene expression associated with estrogen-induced prolif- eration of human breast cancer cells. Cancer Res 1987;47:
6517-21.
9) Perillo B, Sasso A, Abbondanza C, Palumbo G. 17beta- estradiol inhibits apoptosis in MCF-7 cells, inducing bcl-2 expression via two estrogen-responsive elements present in the coding sequence. Mol Cell Biol 2000;20:2890-901.
10) McKenna NJ, Xu J, Nawaz Z, Tsai SY, Tsai MJ, O'Malley BW. Nuclear receptor coactivators: multiple enzymes, multiple complexes, multiple functions. J Steroid Biochem Mol Biol 1999;69:3-12.
11) Prall OW, Sarcevic B, Musgrove EA, Watts CK, Sutherland RL. Estrogen-induced activation of Cdk4 and Cdk2 during G1-S phase progression is accompanied by increased cyclin D1 expression and decreased cyclin-dependent kinase inhibitor association with cyclin E-Cdk2. J Biol Chem 1997;272:10882- 94.
12) Chomczynski P, Sacchi N. Single-step method of RNA isola- tion by guanidium thiocyanate-chloroform extraction. Anal Biochem 1987;162:156-9.
13) Ray R, Chen G, Vande Velde C, Cizeau J, Park JH, Reed JC, et al. BNIP3 heterodimerizes with Bcl-2/Bcl-X (L) and induces cell death independent of a Bcl-2 homology 3 (BH3) domain at both mitochondrial and nonmitochondrial sites. J Biol Chem 2000;275:1439-48.
14) Nancy A., Yuri L. Caspase: Enemies within. Science, 1998;
281:1312-6.
15) Soltys BJ, Gupta RS. Interrelationships of endoplasmic retic- ulum, mitochondria, intermediate filaments, and microtubules -a quadruple fluorescence labeling study. Biochem Cell Biol 1992;70:1174-86.
16) Shi J, Peng Y, Li N, Gu W, Han Z, Fu G, et al. Novel genes expressed in human dentritic cell. GeneBank Accession No.
AF201944.
17) Petit PX, Lecoeur H, Zorn E, Dauguet C, Mignotte B, Gougeon ML. Alterations in mitochondrial structure and func- tion are early events of dexamethasone-induced thymocyte apoptosis. J Cell Biol 1995;130:157-67.
18) Zoratti M, Szabo I. The mitochondrial permeability transition.
Biochim Biophys Acta 1995;1241:139-76.
19) Susin SA, Lorenzo HK, Zamzami N, Marzo I, Snow BE, Brothers GM, et al. Molecular characterization of mito- chondrial apoptosis-inducing factor. Nature 1999;397:441-6.
20) Li P, Nijhawan D, Budihardjo I, Srinivasula SM, Ahmad M, Alnemri ES, et al. Cytochrome c and dATP-dependent for- mation of Apaf-1/caspase-9 complex initiates an apoptotic protease cascade. Cell 1997;91:479-89.
21) Berthois Y, Dong XF, Martin PM. Regulation of epidermal growth factor-receptor by estrogen and antiestrogen in the human breast cancer cell line MCF-7. Biochem Biophys Res Commun 1989;159:126-31.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ 22) Weisz A, Bresciani F. Estrogen regulation of proto-oncogenes
coding for nuclear proteins. Crit Rev Oncog 1993;4:361-88.
23) Raff MC. Social controls on cell survival and cell death. Na- ture 1992 356:397-400.
24) White E. Life, death, and the pursuit of apoptosis. Genes Dev 1996;10:1-15.
25) Bardon S, Vignon F, Montcourrier P, Rochefort H. Steroid
receptor-mediated cytotoxicity of an antiestrogen and an antiprogestin in breast cancer cells. Cancer Res 1987;47:
1441-8.
26) Kyprianou N, English HF, Davidson NE, Isaacs JT. Pro- grammed cell death during regression of the MCF-7 human breast cancer following estrogen ablation. Cancer Res 1991;
51:162-6.