88 책임저자:권현주, 614-714, 부산시 부산진구 가야동 산24번
지, 동의대학교 자연과학대학 생명응용학과 Tel: 051-890-1519, Fax: 051-890-1532 E-mail: [email protected]
접수일:2013년 2월 15일, 1차 수정일:2013년 2월 19일, 2차 수정일:2013년 2월 20일, 게재승인일:2013년 2월 22일
Correspondence to:Hyun Ju Kwon
Department of Life Science and Biotechnology, College of Natural Science, Dong-Eui University, San 24, Gaya-dong, Busanjin-gu, Busan 614-714, Korea
Tel: +82-51-890-1519, Fax: +82-51-890-1532 E-mail: [email protected]
노로통(
Fructus liquidambaris
) 메탄올 추출물에 의해 유도된 HT29 대장암 세포의 G1기 세포 주기 정지 기전동의대학교 1블루바이오소재개발센터, 2자연과학대학 생명응용학과, 동의대학교 3한의과대학 생화학교실,
4대학원 바이오물질제어학과(BK21 Program)
윤희정1ㆍ장연경2ㆍ최영현1,3,4ㆍ김병우1,2,4ㆍ권현주1,2,4
Methanol Extract of Fructus liquidambaris Induces G1 Cell Cycle Arret in Human Colon Adenocarcinoma HT29 Cells
Hee Jung Yun1, Yeon Kyeong Jang2, Yung Hyun Choi1,3,4, Byung Woo Kim1,2,4 and Hyun Ju Kwon1,2,4
1Blue-Bio Industry RIC, 2Department of Life Science and Biotechnology, College of Natural Science,
3Department of Biochemistry, College of Oriental Medicine, Dong-Eui University, 4Biomaterial Control (BK21 Program), Graduate School, Dong-Eui University, Busan 614-714, Korea
Fructus Liquidambaris, fructus of Liquidambar formosana Hance fructus, is traditional herbal medicine used in oriental region. There is no report for the anticancer activity of Fructus Liquidambaris yet. In this study, we investigated the anti-cancer effect of methanol extract of Fructus Liquidambaris (MEFL) in human colon adenocarcinoma HT29 cell. Treatment of MEFL to HT29 cells resulted in the growth inhibition and the G1 cell cycle arrest in a dose-dependent manner. In western blot analysis, the level of Cdk2, Cyclin D/E, phospho-pRb and E2F-1, which are G1-cell cycle-related proteins, were decreased in HT29 cells treated with MEFL. And the expression of phospho-p53 (Ser 15), phospho-Chk2 (Thr 68), p21 and p27 were increased, but Cdc25A was down-regulated in a dose-dependent manner. These results revealed that MEFL induces G1 cell cycle arrest through the induction of p21 and p27 by activated p53, and the reduction of Cdc25A by activated Chk2. This study is the first report about anticancer activity of MEFL, additional research is necessary. (Cancer Prev Res 18, 88-95, 2013)
Key Words: Fructus liquidambaris, HT29 cells, G1 cell cycle arrest, p21, Cdc25A
서 론
암은 세포 주기 조절의 교란에 의해 세포가 비정상적 으로 계속 증식되는 것으로,1) 우리 나라 사망 원인 1위의 질환이다. 보건복지부의 국가 암 통계 결과에 의한 암발 생률 추이는 1999년 인구 10만 명당 219.9명에서 2009년 299.4명으로 증가하여 연평균 3.4%의 증가율을 보였고,
갑상선암, 위암, 대장암, 폐암, 간암의 순으로 많이 발생 한 것으로 나타났다. 이 중 대장암은 24,986건 발생, 전체 암환자의 13.0%를 차지하여 우리나라에서 발생한 암의 3위이고, 남성의 경우 연 6.7%, 여성의 경우 연 5.1%의 비율로 매년 발생률이 증가하고 있다. 또한 전 세계적으 로도 매년 백만 명의 대장암 환자가 진단되며 전체 암의 세 번째, 사망원인의 네 번째를 차지하고 있다.2) 이렇게 증가하는 대장암의 발병원인은 정확히 알려져 있지는
않으나 흡연, 과체중과 비만도, 육류 소비량, 과잉 알콜 섭취 등의 식이 요인과 유전적 요인, 육체적 운동 및 활 동량 등과의 관련성들이 보고되어 있다.3~5) 대장암은 혈 관을 중심으로 많은 수의 세포들이 서로 덩어리진 형태 로 성장하는 대표적인 고형 암으로 결장과 직장에 생기 는 악성 종양을 말한다. 따라서 결장과 직장에 암종이 존재할 때는 국소적인 절제수술만으로 치료가 가능하 나, 다른 장기로 전이되었거나 수술 후 재발의 방지 및 생존 기간의 연장을 위해 5-fluorouracil, leucovorin, irinote- can, oxaliplatin 같은 화학요법제가 병용 치료되고 있다.6) 이러한 화학 요법제의 병용 치료로 대장암 환자의 생존 기간이 향상되었으나, 화학 요법제는 환자의 상태 및 처 리 용량과 용법에 따라 다르나 구토, 설사, 탈모 등의 부 작용이 나타난다.7) 이러한 화학 요법제의 부작용을 줄이 고 환자의 생존 기간을 연장시키기 위해 새로운 암치료 제의 개발을 위한 노력이 꾸준히 이루어지고 있고, 최근 에는 비교적 유효성과 안전성이 보장되어 있는 천연물 로부터 항암 활성 물질을 찾는 연구가 활발히 진행되고 있다.8~10)
항암 화학 요법제들은 약물에 따라 다양한 기작을 통 해 암세포의 세포주기를 정지시키거나 세포 자멸사를 유도함으로써 암세포의 성장을 억제시키거나 사멸시킨
다.11,12) 세포 주기는 다양한 분자들에 의해 조절되어 완
전한 genome을 정확하게 딸세포로 전달시킨다. 그러나 이러한 세포 주기 조절계에 이상이 생기면 암세포가 될 가능성이 크다. 세포 주기는 Cyclin과 Cyclin dependent kinases (Cdks)의 활성화에 의해 진행된다.13) 세포주기의 각 단계에는 각 세포 주기별로 특이적인 Cyclin이 발현되 어 각각 다른 종류의 Cdks에 결합하고, 이를 통해 Cdks 가 활성화된다.14) 활성화된 Cyclin-Cdk 복합체는 세포 주 기의 각 단계에 작용하는 단백질들을 인산화시켜 세포 주기의 진행을 조절하는데, 대표적으로 G1기 동안 발현 되는 Cyclin D-Cdk4, Cyclin D-Cdk6 혹은 Cyclin E-Cdk2가 pRb의 인산화를 유도하여 활성화시킨다.14) 불활성형의 pRb는 전사인자인 E2F와 결합하여 S기에 필수적인 E2F 의존성 유전자의 전사를 막아 S기로의 진행이 저해된다.
G1기 동안 Cyclin-Cdk 복합체에 의해 pRb가 인산화되면, E2F와의 결합이 붕괴되고 E2F가 방출되어 S기로 진입하 게 된다.
본 연구에서는 부작용이 적고 효과적인 이상적인 항암 성분을 천연물에서 탐색하고자 예부터 많이 사용되어오 던 한약재 데이터를 검토하여 항암 활성에 대한 보고가 없는 7종의 한약재를 선정하였다. 노로통(Fructus liquidam- baris)을 포함한 7종의 한약재 메탄올 추출물을 사용하여,
매년 발생율이 증가하고 있는 대장암 세포주의 증식을 억제 시키는 물질을 검토한 결과 노로통(Fructus liquidambaris) 메탄 올 추출물에서 가장 높은 활성을 확인하였다. 노로통(Fructus liquidambaris)은 풍향수(楓香樹) Liquidambar formosana Hance (조 록나무과 Hamamelidaceae)의 성숙한 과실로써 냄새가 약간 있 고 맛은 덤덤하다.15) 성미(性味)는 평(平), 신(辛), 고(苦)이 고 공효는 거풍통락해 주며 이수하유(利水下乳)해 주고 지양(止痒) 작용이 있어, 전통적으로 관절통, 산부인과 질병, 피부병 등 치료에 이용되었다.16) 노로통 추출물에 대한 연구로는 항 allergy 작용이 보고되었으며,17) 항암 효과의 가능성이 있으나 정확한 항암 기전을 제시한 보 고는 아직 없다. 따라서 본 연구에서는 노로통(Fructus liq- uidambaris) 메탄올 추출물에 의한 HT29 인체 대장암 세포 주의 증식 억제 기전을 연구한 결과 G1기에서의 세포 주기 정지 효과를 확인하였기에 보고하는 바이다.
재료 및 방법 1. 한약재 추출
본 실험에서 사용된 노로통(Fructus liquidambaris), 구맥 (Dianthus chinensis L.), 금전초(Glechoma hederacea var. longituba), 대복피(Areca catechu L.), 괴화(Sophora japonica L.), 구척 (Cibotium barometz J. Smith), 노근[Phragmites australis (Cav.) Trin ex Steud]은 대한생약에서 구입하였으며, 분쇄한 한약재 10 g에 100% 메탄올 200 ml를 넣고 70oC에서 3시간 동안 2회 추출하였다. 그 후 추출액을 Whatman No. 2 paper (Whatman International Ltd., England)로 여과한 후, 여액을 rotary evaporator (45oC)에서 감압 농축한 후 동결 건조하 여 사용하였다. 노로통의 HT29 세포에 대한 항암 기작 연구를 위한 시료는 분쇄한 노로통 100 g에 100% 메탄 올 1 L를 넣어 75oC에서 3시간 동안 3회 추출하였다. 추 출액을 Whatman No. 2 paper (Whatman International Ltd., England)로 여과한 후, 그 여액을 rotary evaporator에서 45oC에서 감압 농축하여 4.81 g의 고형물을 얻었다. 노로 통 메탄올 추출물(MeOH extracts of Fructus liquidambaris)은 MEFL이라 명명하였다.
2. 암세포 배양
본 연구에 사용된 HT29 인체 대장암 세포는 ATCC (American Type Culture Collection, Manassas, VA, USA)에서 구입하여 사용하였으며, 세포 배양은 RPMI 1640 medium 에 10% (v/v)의 FBS (fetal bovine serum, Omega Scientific Inc., Tarzana, CA, USA)와 10 U/ml의 penicillin과 10 μg/ml의 streptomycin을 첨가한 배지를 사용하였으며 37oC, 5%
CO2 조건 하에서 배양하였다.
3. 한약재 메탄올 추출물의 세포 독성 확인
HT29 세포에서 한약재 메탄올 추출물의 세포 독성 효 과를 확인하기 위해 WST assay를 수행하였다. HT29 세 포를 96-well plate에 1×104 cells/well의 농도로 분주하여 24시간 동안 배양한 후, 7종의 한약재 메탄올 추출물의 초기 농도를 500 μg/well로 처리하고, 단계적으로 희석 해 나가는 방법을 이용하여 시료를 처리하였다. 24시간 반응 후 premixed WST-1 (Cell Proliferation Assay System, Takara, Otsu, Shiga, Japan) 용액을 첨가하고 30분 후 효소 면역 측정(ELISA, Enzyme linked immunosolvent assay) 판독 기를 이용하여 540 nm에서 측정하였다. 세포 독성 확인 후 선정한 노로통의 세포 독성은 위와 동일한 방법으로 재확인하였고 시료 반응 시간은 48시간으로 하였다.
4. 세포 증식 억제 효과 검토
HT29 인체 대장암 세포를 60 mm culture dish에 0.75×106 cells/ml 농도로 분주하여 24시간 배양 후 MEFL을 농도별 (10, 20, 30, 40 μg/ml)로 처리하였으며, 대조군은 0.1%
DMSO로 처리하였다. 배양 시작 후 2일마다(day 0, 1, 3, 5, 7) 회수하여 trypan blue solution (0.4%, Sigma-Aldrich Co., LLC, St. Louis, MO, USA)으로 염색 후 heamocytometer 를 이용하여 생 세포수를 측정하였다. 7일간 배양하였 으며 2일에 한번씩 MEFL이 각 농도별로 포함된 신선한 배지로 교환하였다.
5. Flow cytometry를 이용한 세포 주기 분석
MEFL이 HT29 인체 대장암 세포의 세포 주기에 미치 는 영향을 알아보기 위한 세포주기 분석에는 Cycle TEST PLUS DNA reagent kit (Becton, Dickinson & Co., Franklin Lakes, NJ, USA)를 사용하였다. HT29 세포를 60 mm cul- ture dish에 0.75×106 cells/ml 농도로 분주하여 24시간 배 양 후 MEFL을 농도별(10, 20, 30, 40 μg/ml)로 처리하였 으며, 대조군은 0.1% DMSO로 처리하였다. 48시간 후 세 포를 회수하여 buffer solution으로 세척한 후 Cycle TEST PLUS solution으로 염색시켜 세포를 분석하였다.
6. Western blot analysis
HT29 인체 대장암 세포를 100 mm culture dish에 2×106 cells/ml 농도로 분주하여 24시간 배양 후 MEFL을 농도별 (10, 20, 30, 40 μg/ml)로 처리하였으며, 대조군은 0.1%
DMSO로 처리하였다. 48시간 후에 세포를 회수하여 PBS 로 한번 세척한 후 CSK 완충액 [100 mM Pipes (pH 6.8),
100 mM NaCl2, 1 mM EGTA, 1 mM dithiothreitol (DTT)]에 0.1% Triton X-100, 1 mM ATP와 1% Protease inhibitor Cocktail (BD Pharmingen, San Diego, CA, USA)이 첨가된 용액으로 현탁하여 15분간 반응시킨 후 초음파 파쇄기 로 파쇄하였다. 파쇄된 세포는 14,000 rpm으로 20분간 원 심분리하였으며, Bio-Rad Protein-Assay kit (Bio-Rad Labora- tories Inc., Hercules, CA, USA)를 이용하여 단백질 정량을 실시하고 30 μg의 단백질을 sodium dodecyl sulfate (SDS) - polyacrylamide gel을 이용하여 전기영동하였다. 전기영 동 후 gel 내의 단백질을 PVDF (Polyvinylidene fluoride) membrane (Pall Corporation, Port Washington, NY, USA)에 전사시킨 후 blocking solution [0.15 M NaCl, 1 M Tris-HCl (pH 7.5), 0.1% Triton X-100, 5% BSA]을 사용하여 4oC, 16 시간 blocking 시켰다. 1차 항체는 4oC, 20시간 반응시켰 으며, TBS buffer [50 mM Tris-HCl (pH 7.5), 0.15 M NaCl, 0.1% Triton X-100]를 사용하여 membrane을 세척한 다음 2차 항체를 넣고 4oC, 16시간 반응시켰다. Membrane을 세 척한 후 면역반응 단백질은 화학발광시스템(Super Signal West Femto Maximum sensitivity Substrate, Thermo Fisher Scientific Inc., Waltham, MA, USA)로 검출하였다. 반응의 정도는 FluorChem5500 (Alpha Innotech, Santa Clara, CA, USA)을 사용하여 정량하였다. 사용된 1차 항체 중 Cyclin E, Cyclin D, Cdk2, Cdk4, E2F-1, pRb, Actin, Cdc25A, p53, p21, p27은 Santa Cruz Biothechnology Inc. (San Diego, CA, USA)에서 구입하였고, p-Chk2 (T68)과 p-p53 (S15)는 Cell signaling Technology (Danvers, MA, USA)에서 구입하였다.
결 과
1. HT29 대장암 세포에서 한약재 메탄올 추출물의 세 포 독성 효과
노로통(Fructus liquidambaris), 구맥(Dianthus chinensis L.), 금 전초(Glechoma hederacea var. longituba), 대복피(Areca catechu L.), 괴화(Sophora japonica L.), 구척(Cibotium barometz J. Smith), 노근[Phragmites australis (Cav.) Trin ex Steud] 7종의 한약재 각각 100 g에 100% 메탄올 1 L를 넣어 75oC에서 3시간 동안 3회 추출한 후 감압 농축하였다. 이들 메탄올 추출 물이 HT29 세포에 대해 세포 독성을 나타내는지 확인하 기위해 HT29 세포가 배양된 96-well plate에 각 추출물을 농도별로 첨가하여 24시간 반응한 후 WST assay를 시행 하였다. 그 결과 모든 약재에서 HT29 세포에 대한 독성 효과를 나타내었고, 약 50% 정도의 세포 성장을 억제시 키는 농도인 IC50값을 확인한 결과 노로통 111.5 μg/ml, 구맥 136.7 μg/ml, 금전초 432.2 μg/ml, 대복피 790.9 μg/
Table 1. IC50 of human colon adenocarcinoma HT29 cells treated with MeOH extract of various medicinal herbs
Medicinal herbs IC50 (μg/ml)
Fructus liquidambaris 111.5
Dianthus chinensis L 136.7
Glechoma hederacea var. longituba 432.2
Areca catechu L. 790.9
Sophora japonica L 1,039.4
Cibotium barometz J. Smith 1,276.2
Phragmites australis (Cav.) Trin ex Steud 2,500 The cells were exposed to various concentrations of indicated medicinal herbs for 24 hours. And IC50 of each medicinal herbs in HT29 cells detected by WST assay.
Fig. 2. Antiproliferative effects of MEFL in HT29 cells detected by trypan blue exclusion assay. Cells were treated with MEFL ranging in doses from 0 to 40 μg/ml for 7 days. Control cells were treated with 0.1% DMSO.
Fig. 1. Cytotoxic activity of MEFL in HT29 cells detected by WST assay. The cells were exposed to various concentrations of MEFL for 48 hours.
ml, 괴화 1,039.4 μg/ml, 구척 1,276.2 μg/ml, 노근 2,500 μg/ml으로 노로통이 가장 효과가 좋았다(Table 1). 따라 서 HT29 인간 대장암 세포주에 대한 항암 활성 성분으 로 노로통 메탄올 추출물을 선택하였고, MEFL에 의한 HT29 세포의 성장 억제 기전을 연구하였다.
2. MEFL에 의한 HT29 세포의 세포 독성 효과
MEFL에 의한 HT29 세포의 독성 효과를 더 확인하기 위해 96-well plate에 배양된 세포에 MEFL을 농도별로 처 리하고 48시간 반응 후 WST assay를 시행하였다. WST assay는 수용성의 tetrazolium salt로서 살아있는 세포와 반 응하여 formazan을 생성하여 생세포수를 측정하는 방법 이다. 그 결과 Fig. 1에서 보는 바와 같이 MEFL의 농도가 높아질수록 세포 생장률이 감소하여 20 μg/ml 농도에서 50.98%의 생존률을 보였으며 농도 80 μg/ml에서 28.91%
의 생존률을 나타내었다.
3. MEFL에 의한 HT29 세포의 성장 억제 효과
MEFL의 HT29 대장암 세포에 대한 세포 성장 억제 효 과를 확인하기 위하여, MEFL을 농도별로 처리한 후 7일 간 배양하면서, 2일 간격으로 trypan blue exclusion assay를 통해 생세포수를 측정하였다. Fig. 2에서 보는 것처럼 0.1% DMSO를 처리한 대조군에 비해 MEFL를 처리한 세 포의 생세포수는 배양 1일째부터 현저한 차이를 보여 50 μg/ml의 MEFL을 처리한 세포의 생세포수는 56.7%의 생 존율을 나타내었다. 또한 10, 20, 30, 40, 50 μg/ml의 MEFL를 7일동안 처리 시 생세포수는 대조군에 비해 각 각 15.3%, 15.1%, 12.6%, 8.8%, 5.1%로 대부분의 세포 생 육이 억제되었다. 따라서 MEFL은 농도 및 시간 의존적 으로 HT29 세포의 성장을 억제시킴을 확인하였다.
4. MEFL를 처리한 HT29 세포의 세포 주기 변화
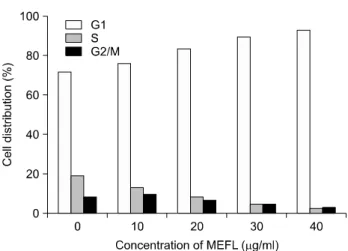
MEFL이 HT29 대장암 세포의 세포주기에 미치는 영향 을 조사하기 위하여 세포에 MEFL을 농도별로 48시간 처 리하고 핵 염색한 후 flow cytometry를 사용하여 세포주기 를 분석하였다. 그 결과 Fig. 3에서 보는 바와 같이 0.1%
DMSO를 처리한 control 세포의 경우 G1, S, G2/M phase의 세포군이 각각 72.17%, 19.26%, 8.56%로 분포하였다. 그 러나 MEFL을 10 μg/ml로 처리한 세포의 경우 G1기의 세포군이 76.37%로 증가하였으며, S기와 G2/M기는 각각 13.6%, 10.03%로 나타났다. 또한 MEFL의 농도가 20 μg/
ml, 30 μg/ml, 40 μg/ml로 증가할수록 G1기의 세포군이 각각 84.09%, 89.96%. 93.26%로 증가하였으며, S기와
Fig. 3. Cell cycle distribution of HT29 cells treated with various concentrations of MEFL and 0.1% DMSO for 48 hours. The cells were collected and stained with PI, and then flow cytometric analysis was performed.
Fig. 5. Expression of Chk2, Cdc25A, p53, p21, p27 and Actin in HT29 cells after treatment of MEFL. Cells wrer treated with various concentration of MEFL for 48 hours and western blot analysis was performend. Actin was used as internal control.
Fig. 4. Western blot anaysis of G1 phase arrest-related proteins in HT29 cells treated with MEFL for 48 hours. Total protein (30 μg) was resolved by SDS-PAGE and western blot performed. Actin was used as internal control.
G2/M기는 감소하는 것으로 나타났다. 따라서 MEFL에 의한 HT29 세포의 성장 저해 현상은 G1 phase에서의 세 포 주기 정지를 통한 것으로 사료된다.
5. MEFL 처리 후 G1기 관련 단백질의 발현 변화
앞에서 FACS anaysis를 통해 MEFL에 의한 HT29 세포 의 G1 arrest 유발 효과를 확인하였다. 따라서 이에 관련 된 단백질의 발현 변화를 확인하기 위해 western blot ana- ysis를 수행하였다. 그 결과(Fig. 4), 세포 주기의 G1기 동 안 Cyclin D와 복합체를 형성하는 Cdk4는 발현량에 큰 변화가 없었고, Cdk4를 활성화시키는 Cyclin D는 MEFL의 농도 의존적인 발현감소를 보여주었다. 또한 G1기 후기
에 가장 발현량이 증가하여 G1/S기의 전이시기에 발현 이 최고조에 달하는 Cyclin E와, Cyclin E와 복합체를 형성 하여 세포주기를 G1기에서 S기로 진행시키는데 중요한 Cdk2의 경우 MEFL의 농도가 증가할수록 발현량이 감소 하는 것으로 나타났다. 또한 세포주기 S기의 개시에 필 요한 전사 인자인 E2F와 결합하여 S기의 개시를 저해하 고 있는 pRb는 Cyclin-Cdks 복합체의 활성화에 의해 인산 화되고, 인산화된 pRb는 전사인자 E2F를 방출하여 S기가 개시된다. MEFL에 의해 pRb와 E2F의 발현에 영향을 주 는지 확인한 결과 인산화된 pRB와 E2F-1 모두 MEFL의 처리에 의해 감소됨을 확인할 수 있었다. 따라서 MEFL 은 Cyclin-Cdks 복합체의 활성을 억제시키고, 이로 인해 pRb에 결합되어 있던 전사인자 E2F의 방출이 저해되어 G1기에서 세포 주기가 억제되는 것으로 사료된다.
6. MEFL에 의한 G1기 세포주기 정지의 기작 검토
G1기 관련 단백질의 발현 변화는 FACS analysis의 결과 와 동일하게 세포주기 G1기에서 S기로 진행하는데 필요 한 단백질의 발현이 유도되지 않았다. 이러한 현상이 어 떤 기작을 통한 결과인지 확인하기 위해 세포주기를 조 절하는 것으로 알려진 단백질들의 발현을 조사하였다 (Fig. 5). 세포주기의 진행에 중요한 조절자인 Cyclin/Cdks 복합체는 Cyclin dependent kinase inhibitor (CKI)인 p21과 p27에 의해 그 활성이 저해된다.13) 따라서 MEFL 처리에 의해 CKIs가 발현되어 세포주기 정지 효과를 나타내는 지 확인해본 결과 MEFL의 농도가 증가함에 따라 p21과 p27의 발현이 크게 증가하였고, p21의 발현을 유도하는 전사인자인 p53은 발현량에 큰 변화가 없었으나 인산화 된 p53 (Ser 15)은 MEFL의 농도 의존적으로 증가함을 확
인할 수 있었다. 또다른 세포주기의 조절자인 Cdc25A는 Cdk2의 Thr 14와 Thr 15 잔기의 탈인산화를 통해 Cyclin E-Cdk2 복합체를 활성화시켜 G1에서 S기로 세포주기를 진행시킨다.18,19) 이러한 Cdc25A의 발현을 검토해 본 결 과 MEFL의 농도 의존적으로 감소하는 것으로 나타났으 며, Cdc25A를 조절하는 것으로 알려진 Chk2 단백질의 발 현을 확인해 본 결과, 활성형인 인산화형 Chk2 (Thr 68) 가 증가하는 것을 확인할 수 있었다. 따라서 MEFL은 Chk2 의 활성화를 통해 Cdc25A 단백질을 감소시켜 Cyclin-Cdks 복합체의 활성을 억제시키고, 동시에 p53의 활성화를 통 해 p21 및 p27의 CKIs를 유도하여 Cyclin-Cdks 복합체의 활 성을 억제시켜 세포주기를 G1기에서 정지시키는 것으로 사료된다.
고 찰
암이란 세포 주기의 조절에 이상이 생겨 비정상적으 로 세포가 증식하는 것으로 발병 원인은 다양하며 지속 적으로 암 발병률과 암 사망률이 증가하고 있는 추세이 다. 그에 따라 다양한 화학요법, 수술요법, 방사선 요법 등의 암 치료법이 개발되고 있으며, 최근에는 합성 물질 로 만들어진 항암제의 부작용이 문제시 되면서 비교적 부작용이 적고 안전성이 높은 천연물부터 항암 효과가 있는 생리활성물질을 탐색하는 연구가 활발하게 이뤄지 고 있다.8~10) 이러한 추세에 따라 본 연구도 천연물에서 항암 효능을 가지는 활성 물질을 찾고자 하였다. 오래전 부터 사용되어 오던 한약재를 대상으로 HT29 세포에 독 성을 나타내는 물질을 스크리닝한 결과 노로통 메탄올 추출물(MEFL)이 가장 높은 효과를 보여(Table 1) MEFL을 이용하여 본 실험을 행하였다. 노로통은 민간요법에서 다양한 질병의 치료약으로 이용되고 있으나 아직 항암 효과에 대한 연구 보고가 없는 천연물로 본 연구에서는 노로통의 항암 효과와 그 기전에 대해 연구하였다.
HT29 세포는 MEFL로 48시간 처리에 의해 농도 의존 적인 세포 독성 효과를 나타내었고(Fig. 1) 농도 및 시간 의존적인 증식 억제 현상이 나타났다(Fig. 2). 이러한 효 과가 세포주기 정지 유도와 관련되는지 확인하기 위해 MEFL을 HT29 세포에 농도별로 48시간 처리한 후 FACS analysis를 시행한 결과 농도 의존적으로 G1기 세포군이 증가하였고(Fig. 3), 어떤 단백질이 관여하는지 확인하고 자 G1기의 세포주기 조절 단백질의 발현 변화를 western blot analysis로 확인하였다(Fig. 4). 세포 주기는 기본적으 로 각 세포 주기에서 활성화 또는 불활성화되는 Cdks에 의해 조절되며 Cdk가 Cyclin과 복합체를 형성함으로써
활성화된다. 활성화된 Cyclin-Cdk 복합체에 의해 세포주 기가 다음 단계로 진행되어 세포 증식이 이루어진다. 그 중 Cdk4는 Cyclin D와 복합체를 형성하며 G1기 동안 활 성화 되며, Cdk2는 Cyclin E와 복합체를 형성하여 G1기 후기에 활성화된다.20) Cyclin D-Cdk4 및 Cyclin E-Cdk2 복 합체는 E2F 전사인자와 결합 상태인 pRB를 인산화시켜 E2F를 방출시켜 세포 주기가 G1기에서 S기로 진행하게 한다. Cdk inhibitors 중 Cip/Kip family에 속하는 p27과 p21 은 Cyclin-Cdk 복합체에 결합하여 그 활성을 억제시킴으 로서 세포 주기의 진행을 억제시킨다.13) p21은 종양억제 유전자인 p53에 의해 활성화되어 전체 세포 주기의 진행 을 억제시키는 것으로 알려져 있다. p53은 다양한 세포 내 반응에 관여하는 단백질로, 세포 내 DNA에 손상이 생겼을 경우 이를 인식하고 신호를 전달하는 단백질 중 하나인 ATM (ataxia telangiectasia-mutated)에 의해 활성화 되는 단백질 중 하나이다.21) ATM은 Ser 15 위치에 p53을 인산화시켜 DNA 손상 신호를 전달시키고, 이렇게 활성 화된 p53은 p21 및 p27의 CKIs를 발현시켜 G1기에서 세 포주기를 정지시킴으로써 손상된 DNA의 회복을 위한 시간을 마련한다. Cyclin-Cdk 복합체의 활성을 조절하는 또 다른 분자인 Cdc25A는 Cdk2의 Thr 14와 Thr 15 잔기 의 탈인산화를 통해 Cyclin E-Cdk2 복합체를 활성화시켜 G1에서 S기로 세포 주기를 진행시킨다.18,19) 세포주기의 진행을 유도하는 Cdc25A는 몇몇 암세포에서 과발현되는 것으로 보고되었다.22,23) Cdc25A가 감소되면 세포 주기의 진행이 정지될 수 있고, 따라서 Cdc25A의 작용을 억제시 키는 물질은 암 치료제로 개발 가능하다. Cdc25A 또한 DNA 손상에 의해 억제될 수 있는데, DNA 손상에 의해 ATM이 활성화되면 하부 단백질인 Chk2의 Thr 68을 인 산화시켜 신호를 전달시키고, 인산화된 Chk2는 Cdc25A 를 인산화시켜 가수분해시킨다.24) Cdc25A가 가수분해되 면 Cdk의 저해적 인산기의 제거가 일어나지 않아 세포 주기가 G1기에서 정지된다. MEFL이 HT29 세포내에서 어떤 기작을 통해 세포주기가 정지되는지 확인하기 위 해, 세포주기를 조절하는 것으로 알려진 단백질들의 발 현을 조사하였다. 그 결과, MEFL의 처리에 따라 Cyclin D와 Cyclin E의 발현이 농도 의존적으로 감소하였고 Cdk2의 감소도 확인되었다(Fig. 4). 또한 Cyclin-Cdk 복합 체의 저해제인 p21과 p27이 증가하였고, 이 현상은 p53 단백질의 활성화에 의한 것으로 생각된다(Fig. 5). 따라서 Cyclin D와 Cyclin E의 발현 감소로 S기 전이에 필요한 Cyclin-Cdk 복합체의 양이 줄어들었고, 생성된 복합체도 Cdk inhibitor인 p21과 p27의 발현 증가로 활성이 저해되 었을 것으로 사료된다. 또한 MEFL의 처리에 의해
Cdc25A가 농도의존적으로 감소하였고, 이 현상은 Chk2 의 활성화에 의한 것으로 생각된다(Fig. 5). MEFL의 처리 에 의해 인산화된 Chk2 (Thr 68)와 p53 (Ser 15)은 세포내 DNA 손상이 발생하면 그것을 인식하고 하부 단백질에 신호를 전달시키는 단백질인 ATM에 의해 인산화되는 것으로 알려져있다.21) 따라서 MEFL은 HT29세포내에서 DNA 손상을 유도하고 이 손상을 회복하기위해 G1기에 서 세포주기를 정지시키는 것으로 사료되며, 그 과정 중 ATM이 p53에 신호를 전달하여 p21 및 p27의 발현을 유 도하고 동시에 ATM이 Chk2에 신호를 전달하여 Cdc25A 의 발현을 감소시켜 p21 및 p27이 효과적으로 Cyclin-Cdk 복합체에 결합, 그 활성을 저해하였을 것으로 사료된다.
또한 MEFL의 처리로 인산화된 pRb의 감소는 Cyclin-Cdk 복합체의 활성 저해에 의한 것으로 사료되며 이에 따라 유리된 E2F-1도 감소한 것으로 판단된다.
이상의 결과로부터 노로통 추출물(MEFL)은 HT29 대장 암 세포에서 Chk2와 p53을 인산화시킴으로써 Cdk in- hibitor인 p21과 p27의 발현을 강력하게 유도하고, 동시에 Cdc25A의 발현을 감소시켜 Cyclin-Cdk 복합체의 활성을 저해시킴으로써 세포주기를 G1기에서 정지시키며, 이러 한 기작에 의해 HT29 세포의 증식을 억제시키는 것으로 사료된다. 이 현상은 DNA 손상에 의해 유도된 현상일 것이라 사료되나 이에 대하여는 추가 연구가 필요하다.
결 론
대장암은 해마다 그 발생률이 증가하고 있는 질병으 로 수술 및 화학요법제의 병용치료로 환자의 생존율이 높아지고 있으나, 화학요법제의 부작용 때문에 암세포 특이적인 항암제의 개발이 필요하다. 본 연구에서는 비 교적 유효성과 안전성이 높은 천연물 유래 한약재를 대 상으로 대장암 세포의 생육 및 성장을 억제시키는 물질 을 탐색하였고, 그 결과 노로통 메탄올 추출물을 활성 물질로 선정하였다. 노로통은 농도 의존적으로 인체 대 장암 세포주 HT29의 성장을 억제시켰고, HT29의 세포 주기를 G1기에서 정지시켰다. 이는 p21 및 p27의 Cyclin- dependent kinase inhibitor의 발현을 강력하게 유도함과 동 시에 Cdc25A의 발현 저해로 Cyclin D와 Cyclin E, Cdk2의 발현을 저해하고 Cyclin-Cdk 복합체의 활성을 저해함으 로써 HT29의 세포 주기를 G1기에서 정지시킴으로서 나 타나는 결과로 사료된다. 또한 이러한 p21 및 p27의 발현 증가와 Cdc25A의 감소는 세포내 DNA 손상 정보를 전달 시키는 단백질인 Chk2와 p53 단백질의 활성화에 의한 결과로 판단된다. 본 연구는 노로통의 항암 활성 기작에
대한 최초의 보고로, 노로통 메탄올 추출물은 G1기에서 세포주기를 정지시켜 항암 활성을 나타내는 물질로 세 포내 DNA의 손상을 유도하는 것으로 사료되나 이에 대 한 추가 연구가 필요하다.
감사의 글
본 논문은 2012년도 동의대학교 연구비(2012AA100) 지원을 받아 연구되었음.
참 고 문 헌
1) Douglas H, Robert A. The hallmarks of cancer. Cell 100, 57-60, 2000.
2) Seo GS. The role of NF-κB in colon cancer. Korean J Gas- troenterol 57, 3-7, 2011.
3) Ahmedin J, Freddie B, Melissa M. Jacques F, Elizabeth W, David F. Global cancer statistics. Ca Cancer J Clin 61, 69-90, 2011.
4) Cha MR, Yoon MY, Kim JY, Hwang JH, Park HR. The cytotoxic effect of the gleditsiae semen extracts on human colon carcinoma cells. J Korean Soc Appl Biol Chem 49, 248- 253, 2006.
5) An YO. Prospective and retrospective studies on risk factors of four most common cancers in korean males: stomach, live, lung and colorectun. Seoul, Ministry of Health & Welfare, 1999.
6) Oh MJ, Song HS. Inhibitory effect of snake venom on colon cancer cell growth through induction of death receptor depen- dent Apoptosis. J Korean Acupuncture & Moxibustion Society 29, 25-35, 2012.
7) Hong YS, Kim TW. Chemotherapy for colorectal cancer.
Korean J Gastroenterol 54, 355-363, 2009.
8) Jo KJ, Yoon MY, Lee MR, Cha MR, Park HR. The anticancer effect of extracts from Vitex rotundifolia on human colon carcinoma cell lines. J Korean Soc Appl Bio Che 50, 228-232, 2007.
9) Youn MJ, Song JH. Cha-ga mushroom water extract induced G0/G1 arrest in b16-f10 melanoma cells. Kor J Orient Physiol Pathol 21, 204-208, 2007.
10) Newman DJ, Cragg GM. Natural products as sources of new drugs over the last 25 years. J Nat Prod 70, 461-477, 2007.
11) Houghton JA. Apoptosis and drug response. Curr Opin Oncol 11, 475-481, 1999.
12) Shapiro GI. Harper JW. Anticancer drug targets: cell cycle and checkpoint control. J Clin Invest 104, 1645-1653, 1999.
13) Sherr CJ. The pezcoller lecture: cancer cell cycle revisited.
Cancer Res 60, 3689-3695, 2000.
14) Michelle DG. Cell cycle control and cancer. Curr Science 81, 515-522, 2001.
15) Yun YP. Coloured illustrations for discrimination of herbal
medicine. Seoul, Korea Food & Drug Administration, p 215, 2009.
16) 동양의학대사전편찬위원회. 東洋醫學大事典. 서울, 경 희대학교출판국, pp 379-380, 1999.
17) Ryu HC, Jung SK, Jung HJ, Lim S, Rhee HG. Fructus Liquidambaris contributes to the chemotaxis of eosinophils and secretion of cytokines in A549 human epithelial cells. J Kor Orient Medic Soc 29, 146-160, 2008.
18) Sørensen CS, Syljuåsen RG, Falck J, Schroeder T, Rönnstrand L, Khanna KK, Zhou BB, Bartek J, Lukas J. Chk1 regulates the S phase checkpoint by coupling the physiological turnover and ionizing radiation-induced accelerated proteolysis of Cdc25A. Cancer Cell 3, 247-258, 2003.
19) Hoffmann I, Draetta G, Karsenti E. Activation of the phos- phatase activity of human cdc25A by a cdk2-cyclin E depen- dent phosphorylation at the G1/S transition. EMBO J 13, 4302-4310, 1994.
20) Katrien V, Dirk R. Van B, Zwi NB. The cell cycle: a review of regulation, deregulation and therapeutic targets in cancer.
Cell Prolif 36, 131-149, 2003.
21) Yang J, Yu Y, Hamrick HE, Duerksen-Hughes PJ. ATM, ATR and DNA-PK: initiators of the cellular genotoxic stress responses. Carcinogenesis 24, 1571-1580, 2003.
22) Gasparotto D, Maestro R, Piccinin S, Vukosavljevic T, Barzan L, Sulfaro S, Boiocchi M. Overexpression of CDC25A and CDC25B in head and neck cancers. Cancer Res 57, 2366-2368, 1997.
23) Galaktionov K, Lee AK, Eckstein J, Draetta G, Meckler J, Loda M, Beach D. CDC25 phosphatases as potential human oncogenes. Science 269, 1575-1577, 1995.
24) Bartek J, Lukas J. Pathways governing G1/S transition and their response to DNA damage. FEBS Letters 490, 117-122, 2001.