Fisetin에 의한 비만세포 Th2 사이토카인 발현 하향 조절
윤수정·표명윤# 숙명여자대학교 약학대학
(Received June 26, 2012; Revised October 26, 2012; Accepted October 30, 2012)
Down-regulation of T Helper 2-Associated Cytokine Expression by Fisetin
Soo Jeong Yoon and Myoung Yun Pyo#
College of Pharmacy, Sookmyung Women’s University, Seoul 140-742, Korea
Abstract — Mast cells play pivotal pathologic roles in allergic disease involving T helper 2 (Th2) cytokine such as inter- leukin (IL)-4 and IL-13. Fisetin has been known as an anti-allergic agent having inhibitory effects on the IL-4 and IL-13 gene expressions in inflammatory immune cells. However, its molecular mechanisms for suppressive effects of fisetin on IL-4 and IL-13 in activated mast cells have been incompletely elucidated. In this study we found that fisetin significantly inhibited the phorbol 12-myristate 13-acetate (PMA) and ionomycin (PI)-induced production of IL-4 and IL-13 in mast cells. The lev- els of mRNA were dramatically decreased by fisetin, indicating the suppression might be regulated at the transcriptional lev- els. Western blot analysis of the nuclear expression of various transcription factors involved in the promoter activation indicated that suppression of c-Fos was prominent together with significant down-regulation of nuclear factor of activated T-cell (NF-AT) and NF-κB, but not c-Jun. Furthermore, the nuclear expression of GATA binding protein 2 (GATA-2) tran- scription factor was significantly down-regulated by fisetin. Taken together, our study indicated fisetin has suppressive effects on IL-4 and IL-13 gene expression through the regulation of selective transcription factors.
Keywords □ fisetin, IL-4, IL-13, mast cells, transcription factors
비만세포는 주로 피부, 위장관 점막, 혈관, 림프관주위, 호흡기 등 전신의 장기에 고루 분포되어 있고 천식, 염증성 장 질환, 아 토피와 같은 알러지성 질환의 염증반응에 관여하는 세포로 알려 져 있다.1,2)비만세포가 알러지 항원에 노출되면 B세포에서 생성 된 IgE와 비만세포 표면에 부착된 FcεRI(IgE 고친화성 수용체) 가 결합되어 활성화 상태가 되고, 세포의 과립 내에 저장되어있 던 히스타민, 류코트리엔류 대사물질 및 염증반응을 일으키는 사 이토카인을 분비한다. 이러한 물질들은 점막 부종, 기관지 평활 근 수축, 점액분비증가 등의 알러지 반응을 일으킨다.3)비만세포 는 이러한 즉각적인 반응에만 국한되어 작용하는 것은 아니며, 만성 알러지 반응에도 깊이 연관되어 있다는 연구가 보고된 바 있다.4)비만세포에서 생성되는 인터루킨-4(Interleukin-4, IL-4), IL-13 같은 Th2 사이토카인은 면역반응을 증가시키고, IgE의 생 산을 증가시킴으로써 만성 알러지 반응에 기여하는데, 특히 천 식과 깊은 연관이 있는 것으로 알려져 있다.5,6)활성화된 비만세
포에서 분비된 IL-4는 Ig의 class switching을 유도하여 isotype 전환을 일으켜 알러지 반응을 확대시킨다. 한편 IL-13은 천식 기 관지에서 호산구성 염증반응, 기도상피하의 섬유화, 기관지 과민 반응, 점액 과분비를 일으키는데 직접적으로 관여하여 알러지 증 상을 확대한다.7)또한 IL-13 결핍마우스에 항원을 투여하였을 때 기도 과민성이 나타나고, IL-4 및 IL-5가 발현되었음에도 불구하 고 점막생산과 기도 과민 반응이 나타나지 않는다는 실험결과는 IL-13이 천식을 유발하는데 있어 매우 특이적인 역할을 수행함 을 나타낸다.8)이러한 결과들은 IL-13이 다른 Th2 사이토카인 으로부터 독립적인 작용을 수행하며 알러지 염증을 야기시키기 에 충분하며 필수적이란 것을 의미한다. 따라서 비만세포가 분 비하는 Th2 사이토카인의 유전자발현을 조절하는 것은 알러지 성 질환을 치료하는 하나의 방법으로 제시될 수 있다.
IL-4와 IL-13 유전자 발현은 세포 종류에 따라서 nuclear factor of activated T cells(NF-AT), NF-κB, activator protein 1(AP- 1), GATA 등의 여러 전사인자들에 의해 조절된다(Fig. 1). NF- AT는 Ca2+-dependent phosphatase calcineurin과 nuclear localization에 의해 조절되며, 비만세포 IL-13의 발현에 중요하게 관여할 뿐만 아니라9) IL-4 전사를 조절한다.10) NF-κB는 p65와
#본 논문에 관한 문의는 저자에게로
(전화) 02-710-9573 (팩스) 02-710-9573 (E-mail) [email protected]
종설
p50 단백질이 세포질 내에 존재하고 있으며 적절한 자극에 의하 여 인산화된 후 핵 내로 이동하여 heterodimer를 형성하여 특이 적 DNA 결합부위에 결합하는데, 사람 비만세포연구에 따르면 NF-κB가 IgE-FcεRI 반응에 의한 IL-13 생산에 필수적이라는 것 이 알려져 있다.11) AP-1은 c-Jun과 c-Fos로 구성되며 이들은 유 전자의 발현이 유도되고 인산화가 일어남에 따라 활성화된다. 최 근 연구에 따르면, 비만세포 FcεRI에 IgE가 결합되면 protein kinase D를 통해 AP-1을 활성화시켜 IL-13 생산을 유도한다고 알려져 있다.12) GATA는 비만세포와 호염기구에서 주로 발현되 는 전사인자이며 염색질 재조합 과정 조절과 프로모터 활성화를 통해 비만세포에서 IL-4와 IL-13 발현에 필수적인 역할을 한다 고 알려져 있다.13,14)
Fisetin(3,7,3',4'-tetrahydroxyflavone)은 플라보노이드의 일종으 로 딸기, 사과, 감, 양파와 같은 과일과 채소류에 존재하는데 약 2~160µg/g의 농도로 존재하고 항암작용,15)항산화작용,16)항염 증작용17)등에 대한 많은 연구결과들이 있다. 또한 fisetin은 호 염기구에서 Th2 사이토카인 발현을 억제하는 항알러지 효과를 가지며,18)비만세포에서 전 염증 사이토카인 발현을 억제하는 효 과를 지니는 것으로 잘 알려져 있다.19)그러나 이러한 작용의 세 포분자적 메커니즘에 대한 연구는 거의 이루어지지 않았다. 그 러므로 본 연구에서는 비만세포에서 fisetin이 IL-4와 IL-13 사이 토카인 발현억제를 세포 분자적 수준에서 어떻게 조절하는지를 이해함으로써 알러지 질환의 예방 및 치료를 위한 기능성 소재 개발로의 가능성을 알아보고자 하였다.
실험방법
실험재료
Fisetin은 Sigma Aldrich에서 구매하여 에탄올에 녹인 후 0.2 µm syringe filter로 여과한 후 PBS로 농도에 맞게 희석하여 세 포에 가하였다. Phorbol 12-myristate 13-acetate(PMA)와 ionomycin은 Sigma Aldrich에서 구입하였다.
세포배양
본 실험에 사용된 P815 비만세포는 American Type Culture Collection(ATCC)에서 구매하였다. Lonza에서 구입한 Dulbecco's
Modified Eagle's Medium(DMEM)에 15% fetal bovine serum (FBS), penicillin 100 U/ml, streptomycin 100µg/ml을 첨가한 세포배양액을 세포에 가하여 37oC, 5% CO2 incubator에서 배양 하였다.
세포생존율 측정
Fisetin에 의한 비만세포생존율을 알아보기 위하여 MTT assay 를 수행하였다. 비만세포(5×105cells/ml)를 96-well plate에 분 주한 뒤 fisetin을 다양한 농도(0.09, 0.18, 0.37, 0.75, 1.5, 3, 6 µM)로 24시간 동안 처리한 후, Thiazolyl blue tetrazolium bromide(MTT) reagent 50µl를 가하고 37oC에서 4시간 동안 배 양 후에 DMSO 50 µl를 가하여 formazan을 용해시킨 뒤 ELISA Microplate Reader를 이용하여 540 nm에서 흡광도를 측정하였다.
사이토카인측정
비만세포(5×105cells/ml)를 48-well plate에 분주하고, fisetin 을 세포 생존에 영향을 미치지 않는 농도(0.75, 1.5, 3 µM)로 1 시간 동안 처리한 다음, 비만세포를 활성화시키기 위해 PMA(10 ng/ml)와 ionomycin(0.25 µM)을 가하고14) 16시간 배양하여 얻은 세포배양액을 원심분리하여 상등액 중의 사이토카인을 sandwich ELISA kit(BD Science, Invitrogen)을 이용하여 측정하였다.
RT-PCR
Fisetin이 사이토카인 mRNA 발현 정도에 미치는 영향을 측정 하기 위하여, 비만세포(5×105cells/ml)에 fisetin을 처리한 후 PI 를 가하여 6시간 동안 배양한 후 TRIzol(Invitrogen)을 이용하여 total RNA를 분리하였다. 각각의 샘플은 cDNA synthesis (iNtRON Biotechnology)를 사용하여 45oC에서 60분, 95oC에서 5분간 반응시켜 cDNA를 만들었다. cDNA는 각 primer와 반응 시켜 세포가 발현한 사이토카인의 mRNA의 양을 gel running을 통해 확인하였다. 반응은 95oC에서 10분 동안 변성과정을 거친 후, 95oC 30초, 52oC 40초, 72oC 30초 순으로 35회 반복한 후 72oC에서 5분간 elongation 후 증폭을 완료하였다. 사용된 primer 의 sequences는 다음과 같다. IL-4 sense, 5'-ATG GGT CTC AAC CCC CAG C-3'; IL-4 antisense, 5'-GCT CTT TAC GCT TTC CAG GAA GTC-3'; IL-13 sense, 5'-GGA GCT GAG CAA CAT CAC ACA-3'; IL-13 antisense, 5'-GGT CCT GTA GAT GGC ATT GCA-3'. β-actin sense, 5'-ACC GTG AAA AGA TGA CCC AG-3'; β-actin antisense, 5'-TCT CAG CTG TGG TGG TGA AG-3'.9,19,20)
Western blot analysis
Fisetin이 사이토카인 발현에 관여하는 전사인자들에 미치는 영향을 알아보기 위하여 비만세포(5×105cells/ml)에 fisetin을 처 Fig. 1− Overview of the regulatory regions in the IL-4 and IL-13
promoter.
리한 후 PI를 가하여 3시간 동안 배양한 후 Nuclear extract kit(Active Motif)를 이용하여 세포로부터 nuclear extract를 얻었 다. Nuclear extract는 nuclear extract kit(Active motif)를 사용 하였다. Hypotonic buffer를 넣고 얼음에서 15분 동안 방치한 후 detergent를 넣고 vortexing을 하였다. 4oC, 14000×g에서 30초 간 원심분리하여 상등액을 버린 후 남아있는 pellet에 complete lysis buffer를 넣고 얼음에서 30분 동안 방치한 후 vortexing을 하였다. 4oC, 14000×g에서 10분간 원심분리하여 상등액을 얻었 다. 이렇게 얻은 단백질은 BCA 단백질 정량법(Pierce)으로 정량 하여 10% SDS-PAGE로 분리한 후 PVDF membrane에 transfer 하였다. Membrane을 2% skim milk(0.1% PBS/T buffer)로 실 온에서 1시간 동안 blocking한 뒤, 일차항체 NF-ATc1, GATA-2, YY-1(Santa Cruz Biotechnology), NF-κB p65, c-Fos(Cell signaling), c-Jun(Gene Tex) 을 4oC에서 하룻밤 방치한 후 이차 항체 Anti-mouse IgG, Anti-Rabbit IgG(Cell signaling)를 실온 에서 1시간 동안 반응시켰다. ECL detection용액을 사용하여 Luminescent Image Analyzer로 나타난 band의 두께를 비교하 여 단백질 발현 유무 및 그 차이를 확인하였다.
통계처리
본 연구에서 얻은 실험결과는 mean±S.D. 값으로 표시하였고, 실험군간의 데이터는 student's t-test로 분석하여 유의성을 검정 하였으며, p값이 0.05 이하인 것만을 통계적으로 유의성이 있는 것으로 간주하였다.
실험결과
Fisetin이 비만세포생존율에 미치는 영향
Fisetin이 비만세포생존율에 미치는 영향에 대해 알아보기 위 하여 다양한 농도(0.09, 0.18, 0.37, 0.75, 1.5, 3, 6 µM)로 비만세 포에 24시간 처리하고 MTT assay를 수행하였다. 아무것도 처 리하지 않은 세포를 대조군으로 하여 100%로 나타냈을 때, fisetin
Table I− Effect of fisetin on cell viability of mast cells. The data presented are expressed as mean±SD from three- independent experiments
Fisetin P815
Concentration (µM) Cell viability (%)
0 00.100±0.67
0.09 100.85±0.25
0.18 100.52±0.50
0.37 099.76±0.46
0.75 099.88±0.29
1.5 095.16±1.71
3 091.03±1.68
6 076.49±5.33**
**p<0.01 vs. nontreated group.
Fig. 2− Suppressive effects of fisetin on IL-4 cytokine production and gene expression in mast cells. Other experimental conditions were as described in material and methods. (A) The levels of IL-4 and (B) the mRNA expression in mast cells were determined by ELISA and RT-PCR, respectively.
The mRNA expression was quantitated by densitomeric analysis with that of the control being 1 fold. The data presented are expressed as mean±SD from three- independent experiments. #p<0.05; ###p<0.001 vs. normal group. *p<0.05; **p<0.01; ***p<0.001 vs. PI-treated group.
0.09~3µM 농도 사이에서 세포생존율에 큰 영향을 미치지 않았 다(Table I).
PI로 활성화된 비만세포에서 fisetin의 IL-4 생성 억제 효과 Fisetin이 비만세포에서 PI 유도로 생성되는 IL-4에 미치는 영 향에 대해 알아보기 위하여 ELISA assay를 수행하였다. 본 실 험결과에서 보면 PI를 처리하지 않은 정상세포에 비해 PI를 처 리한 세포에서 IL-4 분비가 4.4배 증가하였다. Fisetin을 처리하 였을 경우 PI를 처리한 군의 IL-4 분비를 농도의존적(0.75, 1.5, 3µM)으로 억제하였으며, 특히 3 µM fisetin을 처리했을 때 약 89% 정도의 억제효과를 보였다(Fig. 2A). Cyclosporine A는 면 역억제제로 잘 알려져 있어,21)본 연구에서 positive control로 사 용하였고 PI 처리군의 IL-4 생성을 현저히 감소시키는 것으로 나 타났다.
IL-4 mRNA 발현 정도는 RT-PCR로 측정하였으며, PI에 의해 증가된 IL-4 mRNA의 발현은 fisetin을 0.75, 1.5 µM 처리하였
을 경우 유의적인 억제효과가 보이지 않았고, 3 µM fisetin을 처 리하였을 때 가장 효과적으로 억제되었다(Fig. 2B). 이러한 연구 결과는 PI로 활성화된 비만세포에서 fisetin은 3 µM의 농도에서 IL-4의 생성과 mRNA 발현을 조절할 수 있음을 나타내었다.
PI로 활성화된 비만세포에서 fisetin의 IL-13 생성 억제 효과 Fisetin이 비만세포에서 PI 유도로 생성되는 IL-13에 미치는 영향에 대해 알아보기 위하여 ELISA assay를 수행하였다. PI를 처리하지 않은 정상세포에 비해 PI를 처리한 세포에서 IL-13 분 비가 2.7배 증가하였고, fisetin 처리군에서는 PI를 처리한 군의 IL-13 분비를 농도의존적(0.75, 1.5, 3 µM)으로 억제하였으며, 특 히 3 µM을 처리했을 때 약 73.5%의 억제효과를 보였다(Fig. 3A).
IL-13 mRNA 발현 정도는 RT-PCR로 측정되었고, PI에 의해 증가된 IL-13 mRNA의 발현은 fisetin을 0.75, 1.5 µM 처리하였 을 경우 유의적인 억제효과가 보이지 않았고, 3 µM fisetin을 처 리하였을 때 가장 효과적으로 억제되었다(Fig. 3B). 이러한 연구
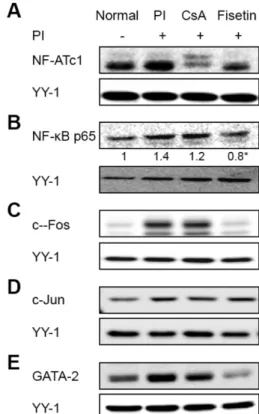
Fig. 4− Effects of fisetin on PI-induced activation of transcription factors involved in the expression of the cytokines expression. Cells were pre-treated with 3µM of fisetin and 1µM of CsA for 1 hr, and then stimulated with PI for 3 hr.
Nuclear extracts were isolated and separated by SDS- PAGE. The level of each transcription factor, (A) NF-ATc1, (B) NF-κB p65, (C) c-Fos, (D) c-Jun and (E) GATA-2 was determined by Western blot analysis using specific antibodies. The numeric values indicate average density of bands from these experiments. The protein expression was quantitated by densitometric analysis with that of the normal being 1 fold. The data presented are representative of three independent experiments. Values are expressed as mean±SD from three-independent experiments. *p<0.05 vs. PI-treated group.
Fig. 3− Suppressive effects of fisetin on IL-13 cytokine production and gene expression in mast cells. Other experimental conditions were as described in material and methods. (A) The levels of IL-13 and (B) the mRNA expression in mast cells were determined by ELISA and RT-PCR, respectively.
The mRNA expression was quantitated by densitomeric analysis with that of the control being 1 fold. The data presented are expressed as mean±SD from three- independent experiments. #p<0.05; ###p<0.001 vs. normal group. *p<0.05; **p<0.01; ***p<0.001 vs. PI-treated group.
결과는 PI로 활성화된 비만세포에서 fisetin은 3 µM의 농도에서 IL-13의 mRNA 발현을 조절함으로써 사이토카인 생성을 억제한 다는 것을 나타냈다.
PI로 활성화된 비만세포에서 fisetin이 사이토카인 발현에 관여하는 전사인자에 미치는 영향
IL-4와 IL-13 promoter에는 NF-ATs, NF-κB, AP-1, GATA 전 사인자의 binding site가 존재한다(Fig. 1). Fisetin이 사이토카인 발현을 억제하는데 어떠한 메커니즘으로 작용하는지 알아보기 위하여 각 전사인자들의 핵 내 발현 정도를 Western blot 분석 법으로 실험하였다. 핵 내 NF-ATc1 발현은 PI를 처리 했을 때 약간 증가하였고, cyclosporine A와 fisetin을 처리한 세포의 경 우에는 PI보다 발현양이 억제된 것을 확인할 수 있었다(Fig. 4A).
NF-κB p65의 경우 PI를 처리했을 때 핵 내 발현 양이 약간 증 가하였고, cyclosporine A를 처리했을 때 변화가 없었으며, fisetin 처리군은 PI에 비해 NF-κB p65가 감소된 것을 확인할 수 있었 다(Fig. 4B). c-Jun과 c-Fos는 AP-1 단백질 복합체의 구성원으로 서 heterodimer 형태로 AP-1 결합부위를 통해 전사를 조절한다.
c-Fos는 PI를 처리했을 때 핵 내 발현 양이 현저하게 증가하였 고, cyclosporine A의 경우에는 PI와 비교했을 때 변화가 없었으 나, fisetin을 처리한 세포는 PI군과 비교했을 때 c-Fos 발현양이 현저하게 억제된 것을 확인할 수 있었다(Fig. 4C). 한편, PI를 처 리하지 않은 정상세포에서 c-Jun의 발현을 확인할 수 있었고 PI 로 활성화된 세포에서 c-Jun의 발현은 약간 증가하였으며(Fig.
4D), cyclosporine A를 처리했을 때는 약간 감소하였고, fisetin 의 경우에는 PI와 비교했을 때 변화가 보이지 않았다. GATA-2 단백질은 세포질 보다는 핵 내에 지속적으로 존재하였고14) PI에 의해 핵 내 발현이 약간 증가했으며 fisetin을 처리했을 때 현저 히 억제된 것을 확인할 수 있었다(Fig. 4E). Cyclosporine A는 GATA-2 단백질 발현양에 영향을 미치지 않는 것으로 나타났다.
이러한 결과로 fisetin이 IL-4와 IL-13 promoter 활성에 관여하 는 전사인자인 NF-ATc1, NF-κB p65, c-Fos, GATA-2의 핵 내 발현에 대해 억제 효과를 가지고 있는 것으로 나타났다.
고 찰
최근 플라보노이드가 함유된 식품의 섭취가 알러지 질환을 개 선시킬 수 있는 잠재성을 가지고 있음을 연구한 결과들이 제시 되고 있다. 플라보노이드가 함유된 과일 등의 섭취가 많은 사람 에게서 천식 발병률이 더 낮았고,22) Knekt 등23)은 플라보노이드 섭취가 천식 및 만성질환을 예방할 수 있는 가능성을 제시하였 다. Astragalin을 NC/Nga 마우스에 경구 투여했을 때 아토피 피 부염 증상의 억제와 혈청 IgE의 수치가 감소된 연구 결과가 보 고된 바 있다.24) Fisetin이 사람 호염기구에서 Th2 사이토카인 억제 효과가 있음이 보고된 바 있으나,18)이러한 작용이 어떠한 세포분자적 메커니즘을 통해 일어나는지에 대한 연구는 아직 완 전히 밝혀져 있지 않다. 본 연구에서는 fisetin이 PI로 활성화된 마우스 비만세포에서 분비된 IL-4와 IL-13의 사이토카인 발현을 농도의존적으로 억제함을 보였고(Fig. 2A and 3A), fisetin 3 µM 의 농도에서만 IL-4와 IL-13 유전자 발현이 현저히 억제됨을 확 인하였다(Fig. 2B and 3B). 또한 3 µM의 fisetin을 처리 했을 때 유전자 발현에 관여하는 전사인자인 NF-AT, NF-κB, c-Fos, GATA-2의 핵 내 발현을 하향 조절할 수 있음을 확인하였다. 비 만세포에서 fisetin은 NF-AT 및 NF-κB 억제를 통하여 IL-4와 IL-13 발현을 감소시키는 것으로 나타났는데, 이는 다른 종류의 세포를 사용한 이전의 연구결과와 일치한다. 즉, Higa 등18)은 A23187로 자극을 준 사람 호염기구에서 fisetin이 NF-AT-DNA
binding 활성을 감소시킴으로써 Th2 사이토카인 발현 억제를 매 개한다고 밝혔고, Park 등25)은 HMC-1 세포에서 사이토카인의 발현이 감소되는 것은 NF-κB와 mitogen activated protein kinases(MAPK)의 억제로 인한 것이라고 보고했다. GATA 단백 질은 T세포에서는 발현되지 않으며 비만세포와 호염기구에서 주 로 발현되고, 특히 비만세포의 분화, 생존, 활성화에 중요한 역할 을 한다고 알려져 있다.26) GATA 공통배열(consensus sequence) 은 IL-4와 IL-13 프로모터에 존재하며, GATA-1 또는 GATA-2의 ectopic 발현은 IL-4 와 IL-13 유전자 프로모터를 활성화시킨다 는 연구가 보고된 바 있다.13,14)이러한 결과들로 봤을 때 본 연 구결과에서 fisetin이 GATA-2 단백질의 핵 내 발현을 억제시킨 것으로 보아 fisetin에 의한 비만세포 조절 효과를 기대할 수 있 다. 뿐만 아니라 GATA 단백질은 IL-4 유전자 인트론에 결합하 여 IL-4 전사를 유도하는데 IL-4 유전자 두 번째 인트론에 PU.1, GATA, STAT5 등의 전사인자가 결합하여 히스톤 탈메틸화를 일 으키고 chromatin remodeling을 조절하여 전사가 잘 일어나도록 하여 프로모터 활성을 통한 전사비율을 증가시켜 준다.13)비만 세포에서 IL-13 생산을 조절하는 메커니즘에 관한 연구는 많지 는 않으나 AP-1과 GATA의 직접적인 상호작용은 IL-13 전사를 증가시킬 수 있고,14)비만세포에 존재하는 NF-AT는 IL-13 프로 모터에 직접 결합하고 GATA 단백질과 상호작용하여 프로모터 의 활성을 증가시킨다는 보고가 있다.27) 본 연구에서는 fisetin이 비만세포에서 c-Fos와 GATA-2 전사인자를 조절하여 IL-4와 IL- 13의 발현조절에 관여할 수 있음을 새롭게 밝혔다. 또한 비만세 포에서 cyclosporine A와 fisetin은 모두 IL-4와 IL-13의 발현을 억제하는 작용을 가졌지만 cyclosporine A는 c-Fos와 GATA-2 단백질 발현에도 영향을 미치지 않았으므로(Fig. 4C and E), 각 각은 서로 다른 메커니즘을 통하여 작용할 것으로 보인다.
본 연구의 결과를 종합하여 볼 때 fisetin은 3 µM의 농도에서 전사인자 NF-ATc1, NF-κB, c-Fos, GATA-2를 선택적으로 조절 하여 IL-4 및 IL-13 유전자 발현을 억제하는 작용을 가졌음이 확 인되었다. 이러한 결과는 fisetin이 비만세포 Th2 사이토카인 조 절을 통해 알러지 질환을 예방할 수 있는 물질로서의 잠재성이 있음을 시사하며 또한 fisetin의 비만세포 Th2 사이토카인 기전 연구에 중요부분 기여할 수 있을 것으로 사료된다. Fisetin이 어 떠한 신호전달 분자와 경로를 통하여 전사인자 조절에 관여하는 지에 대한 추후 실험이 필요하다고 사료된다.
결 론
본 연구에서 비만세포에서 fisetin의 항알러지 작용을 알아보 기 위해 PI로 자극된 비만세포의 IL-4 및 IL-13 사이토카인을 실 험한 결과, 사이토카인 분비뿐만 아니라 mRNA 발현 또한 억제 된 것을 확인하였다. 각 사이토카인 발현에 관여하는 전사인자
들의 핵 내 발현 정도를 확인한 결과 fisetin 3 µM에서 NF-AT, NF-κB, c-Fos, GATA-2의 억제를 확인할 수 있었다. 이러한 연 구결과를 종합해 봤을 때 fisetin이 비만세포 Th2 사이토카인 유 전자발현을 조절하는 효과를 지니는 면역 조절물질로서 잠재적 인 가능성이 있을 것으로 사료되며 알러지 예방에 효과적으로 작 용할 것으로 기대된다.
감사의 말씀
본 연구는 숙명여자대학교 2011학년도 교내연구비 지원에 의 해 수행되었음.
참고문헌
1) Grimbaldeston, M. A., Metz, M., Yu, M., Tsai, M. and Galli, S.
J. : Effector and potential immunoregulatory roles of mast cells in IgE-associated acquired immune responses. Curr. Opin.
Immunol. 18, 751 (2006).
2) Galli, S. J. : Mast cells and basophils. Curr. Opin. Hematol. 7, 32 (2000).
3) Metz, M. and Maurer, M. : Mast cells--key effector cells in immune responses. Trends Immunol. 28, 234 (2007).
4) Galli, S. J., Nakae, S. and Tsai, M. : Mast cells in the development of adaptive immune responses. Nat. Immunol. 6, 135 (2005).
5) Venkayya, R., Lam, M., Willkom, M., Grunig, G., Corry, D. B.
and Erle, D. J. : The Th2 lymphocyte products IL-4 and IL-13 rapidly induce airway hyperresponsiveness through direct effects on resident airway cells. Am. J. Respir. Cell. Mol. Biol.
26, 202 (2002).
6) Brightling, C. E., Symon, F. A., Holgate, S. T., Wardlaw, A. J., Pavord, I. D. and Bradding, P. : Interleukin-4 and -13 expression is co-localized to mast cells within the airway smooth muscle in asthma. Clin. Exp. Allergy 33, 1711 (2003).
7) Wills-Karp, M. and Chiaramonte, M. : Interleukin-13 in asthma. Curr. Opin. Pulm. Med. 9, 21 (2003).
8) Walter, D. M., McIntire, J. J., Berry, G., McKenzie, A. N., Donaldson, D. D., DeKruyff, R. H. and Umetsu, D. T. : Critical role for IL-13 in the development of allergen-induced airway hyperreactivity. J. Immunol. 167, 4668 (2001).
9) Klein, M., Klein-Hessling, S., Palmetshofer, A., Serfling, E., Tertilt, C., Bopp, T., Heib, V., Becker, M., Taube, C., Schild, H., Schmitt, E. and Stassen, M. : Specific and redundant roles for NFAT transcription factors in the expression of mast cell- derived cytokines. J. Immunol. 177, 6667 (2006).
10) Weiss, D. L., Hural, J., Tara, D., Timmerman, L. A., Henkel, G.
and Brown, M. A. : Nuclear factor of activated T cells is associated with a mast cell interleukin 4 transcription
complex. Mol. Cell. Biol. 16, 228 (1996).
11) Lorentz, A., Klopp, I., Gebhardt, T., Manns, M. P. and Bischoff, S. C. : Role of activator protein 1, nuclear factor-kappaB, and nuclear factor of activated T cells in IgE receptor-mediated cytokine expression in mature human mast cells. J. Allergy Clin. Immunol. 111, 1062 (2003).
12) Yamashita, K., Gon, Y., Shimokawa, T., Nunomura, S., Endo, D., Miyata, N., Hashimoto, S., Van Lint, J. and Ra, C. : High affinity receptor for IgE stimulation activates protein kinase D augmenting activator protein-1 activity for cytokine producing in mast cells. Int. Immunopharmacol. 10, 277 (2010).
13) Kwan, M., Powell, D. R., Nachman, T. Y. and Brown, M. A. : An intron GATA-binding site regulates chromatin accessibility and is essential for IL-4 gene expression in mast cells. Eur. J.
Immunol. 35, 1267 (2005).
14) Masuda, A., Yoshikai, Y., Kume, H. and Matsuguchi, T. : The interaction between GATA proteins and activator protein-1 promotes the transcription of IL-13 in mast cells. J. Immunol.
173, 5564 (2004).
15) Suh, Y., Afaq, F., Johnson, J. J. and Mukhtar, H. : A plant flavonoid fisetin induces apoptosis in colon cancer cells by inhibition of COX2 and Wnt/EGFR/NF-kappaB-signaling pathways. Carcinogenesis 30, 300 (2009).
16) Arai, Y., Watanabe, S., Kimira, M., Shimoi, K., Mochizuki, R.
and Kinae, N. : Dietary intakes of flavonols, flavones and isoflavones by Japanese women and the inverse correlation between quercetin intake and plasma LDL cholesterol concentration. J. Nutr. 130, 2243 (2000).
17) Nagai, K., Takahashi, Y., Mikami, I., Fukusima, T., Oike, H. and Kobori, M. : The hydroxyflavone, fisetin, suppresses mast cell activation induced by interaction with activated T cell membranes. Br. J. Pharmacol. 158, 907 (2009).
18) Higa, S., Hirano, T., Kotani, M., Matsumoto, M., Fujita, A., Suemura, M., Kawase, I. and Tanaka, T. : Fisetin, a flavonol, inhibits TH2-type cytokine production by activated human basophils. J. Allergy Clin. Immunol. 111, 1299 (2003).
19) Jin, M., Park, S. and Pyo, M. Y. : Suppressive effects of T-412, a flavone on interleukin-4 production in T cells. Biol. Pharm.
Bull. 32, 1875 (2009).
20) Wu, X., McKenna, M. A., Feng, X., Nagy, T. R. and McDonald, J. M. : Osteoclast apoptosis: the role of Fas in vivo and in vitro.
Endocrinology 144, 5545 (2003).
21) Choi, J. J., Park, B. K., Song, G. Y., Kim, J. S., Kim, J. H., Kim, D. H. and Jin, M. : Establishment of an in vitro test system to evaluate the down-regulatory activities of natural products on IL-4. Arch. Pharm. Res. 30, 1102 (2007).
22) Garcia, V., Arts, I. C., Sterne, J. A., Thompson, R. L. and Shaheen, S. O. : Dietary intake of flavonoids and asthma in adults. Eur. Respir. J. 26, 449 (2005).
23) Knekt, P., Kumpulainen, J., Jarvinen, R., Rissanen, H., Heliovaara, M., Reunanen, A., Hakulinen, T. and Aromaa, A. : Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr.
76, 560 (2002).
24) Kotani, M., Matsumoto, M., Fujita, A., Higa, S., Wang, W., Suemura, M., Kishimoto, T. and Tanaka, T. : Persimmon leaf extract and astragalin inhibit development of dermatitis and IgE elevation in NC/Nga mice. J. Allergy Clin. Immunol. 106, 159 (2000).
25) Park, H. H., Lee, S., Oh, J. M., Lee, M. S., Yoon, K. H., Park,
B. H., Kim, J. W., Song, H. and Kim, S. H. : Anti-inflammatory activity of fisetin in human mast cells (HMC-1). Pharmacol.
Res. 55, 31 (2007).
26) Masuda, A., Hashimoto, K., Yokoi, T., Doi, T., Kodama, T., Kume, H., Ohno, K. and Matsuguchi, T. : Essential role of GATA transcriptional factors in the activation of mast cells. J.
Immunol. 178, 360 (2007).
27) Monticelli, S., Solymar, D. C. and Rao, A. : Role of NFAT proteins in IL13 gene transcription in mast cells. J. Biol. Chem.
279, 36210 (2004).