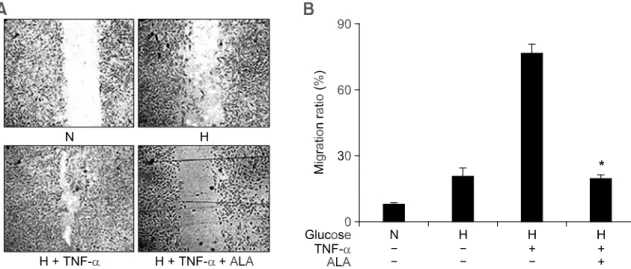
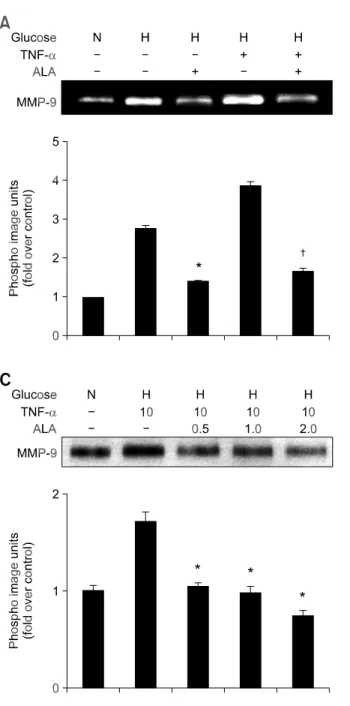
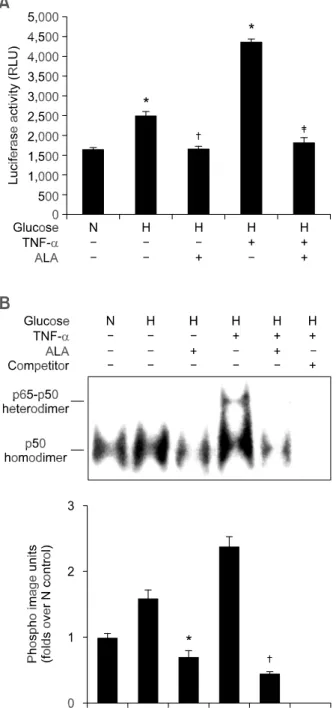
관련 문서
In this way, it is possible to shorten the time of accident restoration and maintenance by minimizing the expression error of the high-strength facial
This study tried to find out the effect of physical expression activity in the childlcare classroom influencing to the aggressions and the adjusting to school of the
The matrix A show the cost per computer (in thousands of dollars) and B the production figures for the year 2005 (in multiples of 1000 units).. Find a matrix C that
This study suggested diversity in the configuration of expression through expression of abstract images, the style of modern painting, which are expressed
The results of this study are as follows: ⑴ 4 basic directions, 16 themes, and 64 sub-themes of the vocational high school development are established, and
These results suggest that the bilobalide inhibits the cell proliferation and induces the apoptotic cell death in FaDu human pharyngeal squamous cell carcinoma via both
In this study, we tested whether a potent AMP-activated protein kinase (AMPK) activator, nectandrin B inhibits VSMC proliferation and neointiima formation and tried to
Generally urban freeway is the principal arterial corridor that is intended to provide for high levels of safety and efficiency in the movement of