INTRODUCTION
1. Research Necessity
Adolescence is a crucial and transition period from childhood to adulthood. Adolescence experience changes in physical and mental ca- pabilities [1]. In physiological maturation, secondary sex-characteristics are appeared by hormonal changes and body size is increased. Mean- while, adolescence is emotional sensitive period so emotional response is delicate, flexible [2,3]. Thus, the adolescents appear prominent tenden- cies toward risk-taking, sensation-seeking, and reckless behaviors (e.g., experimentation with alcohol and smoking) [3,4].
National survey on drug use and health (NSDUH) [6] reported 11.6%
of adolescent aged 12 to 17 had experiences of alcohol use. Furthermore, binge drinking and heavy drinking rates were reported 6.2% and 1.2%, re- spectively. Alcohol use during adolescence is more harmful in cognitive
function than the adulthood because cortico-limbic regions (e.g., hippo- campus) are developed through neuronal plasticity during in this period [1,7]. In alcohol used adolescence, hippocampal volume as well as verbal and learning and memory function were diminished significantly [8,9].
In adolescence animal model, intermittent ethanol exposure caused in- flammatory damage in prefrontal cortex and hippocampus and impair- ment of short-term memory, long-term memory and motor function [10].
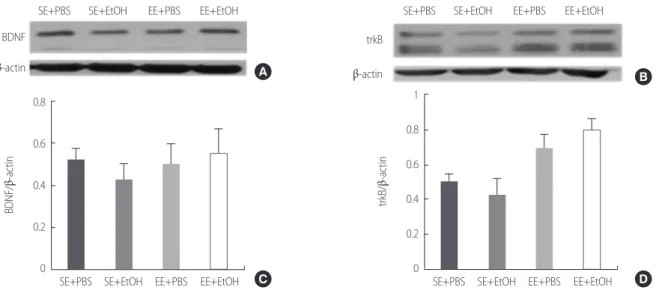
Hippocampus plays important role in the learning and memory. Hip- pocampal neuroplasticity is regulated by neurotrophic factors, such as brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), neutrophin-3 (NT-3). BDNF is the most abundant factor in hippocam- pus [11,12]. BDNF is a major regulator of synaptic transmission, play a role in plasticity such as long-term potentiation (LTP) [12] and regulated by cAMP response element-binding (CREB), c-fos that are known as transcription factors [13,14].
Effect of Environmental Enrichment on Cognitive Impairment- induced by Ethanol Exposure in Adolescent Rat
Yoon Ju Kim
1, Jong Min Park
1,2, Min Kyung Song
1, Ho Hyun Seong
1, Youn Jung Kim
1,31