Copyright 2011 The Korean Society of Neuro-Ophthalmology http://neuro-ophthalmology.co.kr 65
Analysis of Smooth Pursuit Initiation Using Step-ramp Stimuli in Normal Subjects: A Pilot Study
Eui-Joong Kim, MD1, Sun-Young Oh, MD1,2, Byoung-Soo Shin, MD1,2, Man-Wook Seo, MD1,2, Seul-Ki Jeong, MD1,2
Department of Neurology1, Chonbuk National University College of Medicine, Research Institute of Clinical Medicine2, Chonbuk National University Hospital, Jeonju, Korea
Background: Using step-ramp stimuli, the initiation of smooth pursuit was investigated in normal subjects.
Methods: We analyzed the onset latency and the initial acceleration during the first 100 ms of the horizontal smooth pursuit in 30 healthy subjects. In the rightward paradigm, the target was presented at the center for 0.5 seconds and moved 2.5° to the left (step) and began moving toward the right (ramp) with 3 target velocities (5°/sec, 10°/sec, and 20°/sec). Leftward paradigms were initiated with the target which is located center for 0.5 seconds moved to the right and then move to the leftward.
Results: Onset latency of pursuit appeared to be shortened as higher the target velocity. As target velocity progressively in- creased, acceleration of pursuit also increased. There were no significant directional differences in the latency and the acceleration of smooth pursuit between rightward and leftward movements. Older subjects showed significantly increased latencies at all fre- quencies and reduced initial accelerations at high frequency.
Conclusions: The target velocity of step-ramp paradigm appeared to affect on the latency and initial acceleration of pursuit sys- tem. The open-loop system of the pursuit was degraded in the elderly subjects. Decreasing sensitivity to retinal image motion or limiting the conversion of visual motion signals into commands for smooth pursuit might be the cause.
Keywords: Smooth pursuit; Step-ramp; Sinusoidal; Onset latency; Initial acceleration
INTRODUCTION
Smooth-pursuit eye movement is was used to stabilize the image of a moving object of interest on the fovea. Such movement iswas based on a cerebro-ponto-cerebellar pathway that had evolved in parallel with fovea vision. The pursuit deficits had been reported with lesions of a number of cortical areas, including frontal, parietal and occipitotemporal region.1,2,3 The initiation of smooth pursuit has been most conveniently studied by measuring eye position and velocity in the first second following presen- tation of either a ramp or a step-ramp stimulus. In most studies, the direc- tional deficit could be suggested the presence of nervous system disease using maintenance component (i.e. gain) of predictable horizontal sinu- soidal target moving. A clinical significance of the onset of pursuit has been overlooked, however, it might be valuable like pursuit maintenance.
Furthermore, it could allow an identification of directional defects by com- parison of pursuit initiations in each side.
There is scarce report about the effects of target velocity and aging on the onset components of smooth pursuit (i.e., onset latency and initial ac- celeration) using step-ramp stimuli. The purpose of this study was to in- vestigate the normal value of initial component of smooth pursuit of on- set latency and initial acceleration and to evaluate the correlation between these parameters and target velocities in young and elderly.
MATERIALS AND METHODS
This study adhered to the tenets of the Declaration of Helsinki and was approved by the Institutional Review Board of Chonbuk National Uni- versity Hospital. Thirty volunteers (21 men and 9 women) were included
Correspondence to: Sun-Young Oh, MD
Department of Neurology, Chonbuk National University Hospital, 634-18 Geumam-dong, Jeonju 561-712, Korea Tel: +82-63-250-1590; Fax: +82-63-251-9363; E-mail: [email protected]
Received: Jun. 8, 2011 / Accepted: Jun. 20, 2011
ORIGINAL ARTICLE
ISSN: 2234-0971 대한안신경의학회지: 제1권 제1호
Clin Neuroophthalmol 1(1):65-68, June 2011
Kim EJ, et al. • Analysis of Smooth Pursuit Initiation Using Step-ramp Stimuli in Normal Subjects
Clin Neuroophthalmol 1(1):65-68, June 2011 66 http://neuro-ophthalmology.co.kr
in this study. The mean subject age was 47.0±19.8 (range, 24-78) years.
Depending on the age, we divided these 30 volunteers into two groups: the young [n=15; mean age=29 (range: 24-34); female number=5] and the elderly groups [n=15; mean age= 66 (range: 48-78); female number= 4]
(Table 1). Exclusion criteria were a physical condition causing inability to complete the test, optic neuropathy, any visual field defect detected by confrontation test, neglect syndrome, ocular motility disorders such as congenital nystagmus or strabismus, and age younger than 15 years or older than 80 years.
Eye position data were scored in MATLAB (Mathwarks, Natick, MA, USA) using a partially automated program that determined the accuracy of each pursuit movement with respect to the required response and the latency from the target onset. To investigate the onset latency and initial acceleration of smooth pursuit, we used step-ramp stimuli that the target was a red spot back-projected onto a semitranslucent tangent screen 1 m from the subject.
Onset latency and initial acceleration of the horizontal pursuit were estimated by using the step-ramp stimulitarget with various rampeach velocitiesy (of 5°/sec, 10°/sec, and 20°/sec respectively in the rightward- and left-ward) paradigm were estimated. The stimulating target twinkles at the center for 0.5 seconds and move laterally 2.5° to one direction and then begins moving toward the opposite direction. It was repeated ten trials for each attempt of the step-ramp target velocity of 5°/sec, 10°/sec, and 20°/sec. For example, in the leftward paradigm, the target presenteds at the center for 0.5 seconds and moved 2.5° to the right and beganins moving toward the left with the velocity of at 5°/sec. It was repeated for ten trials. And then, pursuits leftward withat velocities of 10°/sec and 20°/
sec for each 10 trials were performed with the same methods.
Prior to onset of pursuit, while the eyes were still stationary, the mo- tion of the target on the retina was equal to the motion of the target in the environment. But after the eye began to follow the target, retinal image motion was reduced, being equal to real target motion minus eye motion.
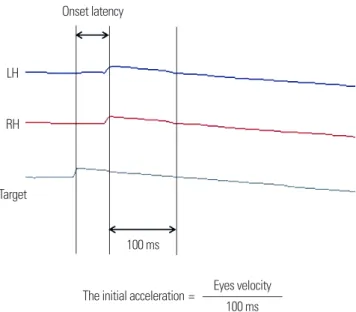
So we measured the onset latency, which was the interval between the on- set of target motion and pursuit onset, and it was the average of 10 attempts.
The initial acceleration was calculated as the ratio between average of eyes velocity and time for the first 100 ms, too (Fig. 1). Response time (latency and acceleration) greater than two standard deviations from the mean wasere also excluded. In addition, smooth pursuit gain wasis appropri- ately determined by the ratio of peak smooth eye movement velocity to peak target velocity using sinusoidal target motions withat the velocity of 10°/sec, 20°/sec, and 40°/sec.
RESULTS
The latency of the smooth-pursuit response was very consistent, with a mean of 147±2 ms to targets moving 5°/sec or faster. This result seemed consistent with the idea that sensory pathways act as a low-pass spatiotem- poral filter of the retinal input, effectively converting position steps into briefly moving stimuli. The latency increased for lower velocity targets;
197±3 ms for target velocities of 5°/sec and 109±1 ms for target veloci- ties of 20°/sec (P<0.001). The initial accelerations increased proportion- ally with elevated target velocity until an acceleration reached about 150°
/sec2 at target velocities of 20°/sec. The acceleration increased slightly with velocity for target velocities in excess of 5°/sec. And higher velocities pro- duced an increase in eye acceleration; there was a near saturation of re- sponse at 44.09±10.4°/sec2 for stimuli moving 5°/sec, 70.46±16.2°/sec2 for 10°/sec and 150.6±67.7°/sec2 for 20°/sec (P<0.001) (Table 2). Most of
Table 1. Participating subjects in this study Young group (age≤ 50,
n= 15)
Elderly group (age> 50,
n= 15)
Total (n= 30) Mean age± SD, yr (range) 28.5± 3.2
(24-34) 65.5± 8.4
(48-78) 47.0± 19.8 (24-78)
Sex ratio, n (men:women) 10:5 11:4 21:9
Fig. 1. In the left-ward paradigm for target velocity 5°/sec, onset latency was the interval between the onset of target motion and pursuit onset, and the ini- tial acceleration was calculated as ratio of eyes velocity and time during the first 100 msec.
LH
Onset latency
100 ms
The initial acceleration = Eyes velocity 100 ms RH
Target
Analysis of Smooth Pursuit Initiation Using Step-ramp Stimuli in Normal Subjects • Kim EJ, et al.
Clin Neuroophthalmol 1(1):65-68, June 2011 http://neuro-ophthalmology.co.kr 67
Table 3. Mean gain of horizontal smooth pursuit eye movement during each target stimuli
10°/sec 20°/sec 40°/sec P-value*
Gain
Rightward 0.82± 0.04 0.77± 0.01 0.61± 0.06 Leftward 0.80± 0.05 0.75± 0.01 0.47± 0.1
Mean 0.81± 0.01 0.76± 0.01 0.55± 0.1 < 0.001
*One way ANOVA. distributed transformation of sensory signal,10 and though the flocculus-
paraflocculus complex access ocular motoneurons via a direct projection to eye-head neuron in the vestibular nuclei.11 Visual area middle tempo- ral serve as a generic visual motion processor that contributes to smooth pursuit by extracting retinal motion of the target; one of the key input that drives smooth-pursuit eye movements. Correspondingly, lesions of selected parts of area of middle temporal render monkeys unable to pur- suit targets moving in the confines of the motion acotoma.7 The latencies were not only similar among subjects, but they were also very repeatable from day to day for individuals.7 This seemed consistent with the idea that sensory pathways act as a low-pass spatiotemporal filter of the retinal in- put, effectively converting position steps into briefly moving stimuli. Al- though latencies were very consistent, they were significantly increased for low target velocities (<5°/sec).7 This increase was well described by a latency model requiring a minimum target movement of 0.028 degrees, in addition to a fixed processing time of 98 ms.4 In our subjects, also as earlier studies, pursuit latencies were significantly increased for low tar- get velocities and elderly group.
The initial acceleration was more variable among subjects than latency.
In this study, the mean initial acceleration for the first 100 ms after gener- ating eye movement was 88.38°/sec2, and higher velocities produced in- crease in eye acceleration. Also, the difference between young and elderly group was significantly only at 20°/sec at which the young groupage showed increased initial eye acceleration. Mild cognitive impairment in Table 2. Mean onset latency and initial acceleration during each target stimuli
5°/sec 10°/sec 20°/sec P-value*
Onset latency (ms) < 0.001
Rightward 212± 28 131± 14 107± 11
Leftward 182± 29 140± 14 111± 14
Mean 197± 29 135.5± 14 109± 12
Initial acceleration (°/sec2) < 0.001
Rightward 45.21± 10.5 71.75± 18.0 160.28± 77.2 Leftward 42.97± 10.1 69.16± 14.3 140.95± 58.2
Mean 44.09± 10.4 70.46± 16.2 150.6± 67.7
*One way ANOVA.
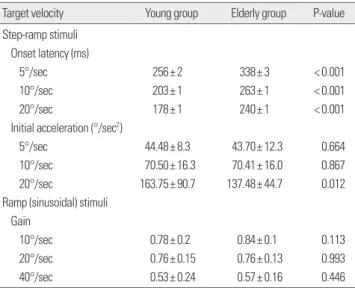
Table 4. Comparison of mean onset latency and initial acceleration in young and elderly subjects
Target velocity Young group Elderly group P-value Step-ramp stimuli
Onset latency (ms)
5°/sec 256± 2 338± 3 < 0.001
10°/sec 203± 1 263± 1 < 0.001
20°/sec 178± 1 240± 1 < 0.001
Initial acceleration (°/sec2)
5°/sec 44.48± 8.3 43.70± 12.3 0.664
10°/sec 70.50± 16.3 70.41± 16.0 0.867
20°/sec 163.75± 90.7 137.48± 44.7 0.012
Ramp (sinusoidal) stimuli Gain
10°/sec 0.78± 0.2 0.84± 0.1 0.113
20°/sec 0.76± 0.15 0.76± 0.13 0.993
40°/sec 0.53± 0.24 0.57± 0.16 0.446
*Gain.
ms; millisecond.
the volunteers showed a small asymmetry between leftward and right- ward onset latency and initial accelerations that was idiosyncratic and present when tested with binocular viewing of the stimulus.
Pursuit gain was measured 0.81±0.01, 0.76±0.01, and 0.55±0.1 at 10°/sec, 20°/sec, and 40°/sec of sinusoidal stimuli (Table 3). Higher veloci- ties produced a decrease in pursuit gain.
Young and elderly subjects had appreciable differences in tracking be- tween each velocities of target. Onset latencies in each target velocity were shorter in the young group than the elderly group (P<0.001), and initial acceleration was larger in the young group than the elderly group, but the statistically significance was found only at the target velocity of 20°/sec (P= 0.012) (Table 4).
DISCUSSION
The onset latency was the interval from onset target motion to gener- ating point of real eye movement. The general latency could be calculated as a sum of the detection time which target moved some threshold dis- tance before it is recognized as a stimulus for the pursuit system and a fixed processing delay.7 Retinal target motion image was mediated to middle temporal area through the sensory pathway, then movement of the target in world coordinates was reconstructed in lateral part of the middle su- perior temporal area.8,9 The dorsal pontine nuclei as intermediary station
Kim EJ, et al. • Analysis of Smooth Pursuit Initiation Using Step-ramp Stimuli in Normal Subjects
Clin Neuroophthalmol 1(1):65-68, June 2011 68 http://neuro-ophthalmology.co.kr
our elderly subjects cannot be excluded as a contributor to their poorer performance, but mental status appeared to be intact in these subjects.
Degraded conversion of visual sensory inputs into motor commands might also explain the senescent decline we recorded in ocular responses to moving stimuli. Cerebellar Purkinje cells in the flocculus and posterior vermis act as a sensory-motor interface for pursuit by constructing a neu- ral representation of target motion in space.12 This target motion signal is probably used as a pursuit motor comrnmand. Senile loss of Purkinje cells could contribute to diminished smooth pursuit function.13 Senes- cent changes in dopaminergic nigrostriatal pathways could also account for some impairment of visually guided eye movements.
In response to step-ramp target motion, elderly subjects had delayed pursuit onset latencies, lower pursuit accelerations than young subjects.
Our finding of diminished initial pursuit acceleration in older subjects indicated that aging degrades smooth pursuit in its first 100 msec, before retinal feedback, extraretinal eye motion feedback, or adaptation begin to influence pursuit performance. This decline in eye acceleration and delayed onset signifies a reduction in the open-loop gain of the pursuit system, which can explain the lower c1osed-loop, steady state pursuit gain observed in the elderly when they track targets with sinusoidal and trian- gular waveforms.5,6 Impaired pursuit initiation in elder1y subjects could be caused by senescent degradation of afferent visual motion inputs. Nor- mal elderly subjects have been shown to have subtle reductions in several visual functions, including visual acuity, visual fields and contrast sensi- tivity, compared to young subjects. Causes of this generalized visual de- cline in aging include diminished photoreceptor function and loss of ret- inal ganglion cell axons and striate and peristriate cortical neurons.
Pursuit maintenance with a sinusoidal target can use prediction to sup- plement retinal image motion in generating eye movement, factors lack- ing in pursuit initiation with random targets.4 The comparison of sinu- soidal pursuit maintenance to step-ramp pursuit initiation is important.
The relationship between sinusoidal and step-ramp directional pursuit defects is unnot clear. Subjects with ipsi-directional pursuit defects had absent pre-saccadic acceleration, and no patients with normal sinusoidal pursuit had abnormalities of step-ramp pursuit.1 Also there were sinusoi- dal pursuit defect alone or step-ramp pursuit defect alone and asymme- tries in both step-ramp and sinusoidal pursuit.2 Although there was like-
ly to fall the correlation between sinusoidal and step-ramp directional pursuit defects, the relationship of two parameters could be identified through further research.
In conclusion, we estimated and compared onset latency and initial acceleration as an initiation component of pursuit in normal young and elderly subjects. The velocity of target movements appeared to affect on latency, initial acceleration and on pursuit gain. Comparison of pursuit initiation in each direction allowed identification of directional defects.
These findings provided a baseline of quantitative information for further studies on the smooth pursuit in patients with the cortical, cerebellar or brainstem lesions.
REFERENCES
1. Thurston SE, Leigh RJ, Crawford T, Thompson A, Kennard C. Two dis- tinct deficits of visual tracking caused by unilateral lesions of cerebral cor- tex in humans. Ann Neurol 1988;23:266-273.
2. Morrow MJ, Sharpe JA. Retinotopic and directional deficits of smooth pursuit initiation after posterior cerebral hemispheric lesions. Neurology 1993;43:595-603.
3. Lekwuwa GU, Barnes GR. Cerebral control of eye movements. The rela- tionship between cerebral lesion sites and smooth pursuit deficits. Brain 1996;119:473-490.
4. Barton JJS, Sharpe JA. Ocular tracking of step-ramp targets by patients with unilateral cerebral lesions. Brain 1988;121:1165-1183.
5. Sharpe JA, Sylveser TO. Effects of aging on horizontal smooth pursuit.
Invest Ophthalmol Visual Sci 1978;17:465-468.
6. Spooner JW, Sakala SM, Baloh RW. Effect of aging on eye tracking. Arch Neurol 1980;37:575-576.
7. Carl JR, Gellman RS. Human smooth pursuit: stimulus-dependent re- sponses. J Neurophysiol 1987;57:1446-1463.
8. Dursteler MR, Wurtz RH. Pursuit and optokinetic deficits following chem- ical lesions of cortical areas MT and MST. J Neurophysiol 1988;60:940-965.
9. Ilg UJ, Schumann S, Thier P. Posterior parietal cortex neurons encode target motion in world-centered coordinates. Neuron 2004;43:145-151.
10. May JG, Keller EL, Suzuki DA. Smooth-pursuit eye movement deficits with chemical lesions in the dorsolateral pontine nucleus of the monkey.
J Neurophysiol 1988;59:952-977.
11. Roy JE, Cullen KE. Brain stem pursuit pathways: dissociating visual, ves- tibular, and proprioceptive inputs during combined eye-head gaze track- ing. J Neurophysiol 2003;90:271-290.
12. Suzuki DA, Keller EL. The role of the posterior vermis of monkey cere- bellum in smooth-pursuit eye movement control. II. Target velocity-re- lated Purkinje cell activity. J Neurophys iol 1988;59:19-40.
13. Hall TC, Miller AK, Corsellis JA. Variations in the human Purkinje cell population according to age and sex. Neuropathology and Applied Neuro- biology 1975;1:267-292.