서 론
비만은 과도한 지방 축적에 의해 야기되어 지방조직 및 체중이 증가된 상태를 말한다1). 지방조직은 insulin 감수성과 생체에너지 의 항상성을 조절하는 중요한 대사기관으로, 지방조직의 증가는 지 방세포의 수가 증가하는 hyperplasia와 지방세포의 크기가 커지는 hypertrophy가 원인이 되어 비만을 초래한다1,2). 특히, 비만은 이 상지질혈증, 고혈압, 심혈관 질환, 제2형 당뇨와 같은 대사성 질환 과 특정 유형의 암 발생과 밀접하게 관련되어 있으며, 세계적인 공 중보건 문제로 대두되고 있다3). 또한 비만은 다양한 사회적 스트레 스로 인한 불안감과 정서적 스트레스 증가, 자기 존중감 저하, 우 울, 그리고 사회적응 수준에 영향을 미쳐 삶의 질을 감소시킨다고 보고되었다4).
Adipogenesis는 미분화된 지방전구세포(preadipocytes)가 지 방세포 (adipocytes)로 완전히 분화되는 과정으로, 지방세포의 형 성 및 지질 축적에 있어 중요한 역할을 한다5). 지방전구세포가 지
방세포로 분화해 나가는 과정에서는 세포의 형태적 변화와 유전자 발현양상의 변화 등이 함께 일어난다6). Adipogenesis는 지방세포 로의 분화하는 과정에서 특이적 전사 인자들과 신호전달기전에 의 해 조절된다. 지방세포분화의 초기 단계에서는 extracellular signal-related kinase (ERK)와 phosphatidylinositide 3-kinase (PI3K)/Akt에 의해 조절되며7,8), 후기단계에서는 CCAAT/enhancer binding proteins α (C/EBPα)와 peroxisome proliferator-activated receptor γ (PPARγ)와 같은 특정한 전사인자들이 성숙한 지방세 포로의 분화를 유도한다9,10). 따라서 지방세포의 분화과정에 관여하 는 전사인자 및 세포신호기전은 비만을 예방하는데 있어 중요한 표 적으로 활용되고 있다11). 전 세계적으로 비만 치료에 대한 우수한 효능과 함께 안전성이 높은 물질의 개발이 요구되고 있는 가운데 특히 천연물 유래의 소재로부터 독성 및 부작용이 없는 항비만 효 능을 가진 소재를 발굴하기 위해 많은 노력을 집중하고 있다12,13). Cymbopogon citratus(국명: 향아, 香芽)는 일반적으로 레몬 그라스로 불리고 있으며 화본과의 여러해살이풀로 풀 전체에 레몬
레몬그라스 에탄올 추출물의 3T3-L1 지방세포 분화 억제효과
조용석1·주성민1,3·황금희1·김민숙2·김광상2·전병훈1,3*
1 : 원광대학교 한의과대학 병리학교실, 2 : 하이솔, 3 : 원광대학교 한국전통의학연구소
Inhibitory Effect of Cymbopogon Citratus Ethanol Extracts on Adipogenesis in 3T3-L1 Preadipocytes
Yong Seok Jo1, Sung Min Ju1,3, Keum Hee Hwang1, Kwang Sang Kim2, Min Sook Kim2, Byung Hun Jeon1,3*
1 : Department of Pathology, College of Korean Medicine, Wonkwang University, 2 : Hisol Co., Ltd., 3 : Research Center of Traditional Korean Medicine, Wonkwang University
Cymbopogon citratus, commonly know as lemongrass, prossesses strong antioxidant, anti-tumor and anti-inflammatory properties. Howerver, its anti‑obesity activity remains to be elucidated. This study investigated the effect of ethanol extract of Cymbopogon citratus on adipogenesis, and its underlying mechanism, in 3T3-L1 preadipocytes. The results demonstrated that ethanol extracts of Cymbopogon citratus effectively suppressed intercellular lipid accumulation at non-toxic concentrations, and was associated with the down-regulation of adipocyte-specific transcription factors, including C/EBPα and PPARγ, and phosphorylation of AMPKα. Furthermore, ethanol extracts of Cymbopogon citratus increased p21 and p21 expression, while the expression of CDK2, cyclin A and cyclin B1 was reduced. As a result, ethanol extracts of Cymbopogon citratus seems to induce G0/G1 cell cycle arrest of 3T3-L1 cells. On the other hand, ERK and Akt signaling pathways were not involved in anti-adipogenesis by ethanol extracts of Cymbopogon citratus. Taken together, theses results suggest that ethanol extracts of Cymbopogon citratus inhibits adipocyte differentiation in 3T3-L1 cells and can be used as a safe and efficient natural substance to manage anti-obesity.
keywords : 3T3-L1 cells, Adipocyte differentiation, Adipogenesis, Cell cycle arrest, Cymbopogon citratus, Lemongrass, Obesity
* Corresponding author
Byung Hun Jeon, Department of Pathology, College of Korean Medicine, Wonkwang University, 344-2, Sinyong-dong, Iksan-si, Jeollabuk-do, Korea
·E-mail : [email protected] ·Tel : +82-63-850-6843
·Received : 2019/01/21 ·Revised : 2019/02/08 ·Accepted : 2019/02/19
ⓒ The Society of Pathology in Korean Medicine, The Physiological Society of Korean Medicine pISSN 1738-7698 eISSN 2288-2529 http://dx.doi.org/10.15188/kjopp.2019.02.33.1.17
Available online at https://kmpath.jams.or.kr
향기가 나며 인도 및 말레이시아와 같은 열대지방에서 주로 재배되 었으나, 최근에는 기후의 변화와 허브의 활용이 늘어나면서 우리나 라에서도 온실재배가 이루어지고 있다14). 레몬그라스 잎으로 우려 낸 차는 근육이완, 진통, 해열 및 항경련성 작용과 스트레스 조절 등의 효과가 있는 것으로 밝혀졌으며15), 최근에는 항산화, 항염증, 항암효과 등이 있는 것으로 보고되었다16,17). 레몬그라스의 성분은 α-pinene, camphene, limonene, myrcene 등으로 구성되어 있 으며, 주성분은 citral이다15,18). Citral은 레몬그라스 정유성분의 65~85%를 차지하며, 강한 항균효과와 해충방제효과를 가지고 있다
18). 최근 citral이 항비만 효과를 가지고 있는 것으로 보고되었는데
19,20), citral 성분을 함유하고 있는 레몬그라스의 조추출물(crude
extracts)에 대한 항비만 연구는 아직 보고된 바 없었다.
본 연구에서는 레몬그라스 에탄올 추출물을 제조하여 3T3-L1 지방전구세포에서 지방생성 억제활성을 평가하였고, adipogenesis 과정과 관련된 전사인자의 발현 및 세포신호기전과 세포주기진행에 미치는 영향을 분석하여 항비만 소재로의 가능성에 대해 고찰하였다.
재료 및 방법
1. 시약 및 항체
Dimethyl sulfoxide (DMSO), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT), propidium iodine (PI), ribonuclease A (RNase A), anti-β-actin 항체 등은 Sigma-Aldrich Chemical (St. Louis, MO, USA)로부터 구입하였다. Dexamethasone, IBMX, insulin 등은 Cayman Chemical (Ann Arbor, MI, USA) 로부터 구입하였다. Halt protease&phosphatase inhibitor cocktail와 RIPA lysis buffer 등은 Pierce Biotechnology (Rockford, IL, USA)로부터 구입하였다. Anti-Akt, anti-phospho-Akt (Ser473), anti-AMPKα, anti-phospho-AMPKα (Thr172), anti-C/EBPα, anti-ERK, anti-phospho-ERK (Thr202/Tyr204), anti-PPARγ 등의 항체들 은 Cell Signaling Technology INC. (Beverly, MA, USA)로부터 구입하였다. Anti-CDK2, anti-cyclin A, anti-cyclin B1, anti-cyclin E, anti-p21, anti-p27 등의 항체들은 Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA)로부터 구입하였다.
2. 레몬그라스 에탄올 추출물의 제조
레몬그라스(Cymbopogon citratus)는 ㈜남원허브(남원, 한국) 에서 구입한 후 정선하여 사용하였다. 레몬그라스 에탄올 추출물은 레몬그라스의 100 g을 20배 가량의 70% 에탄올과 혼합하여 상온 에서 48시간 추출한 후, 여과지(Whatmann No. 5)를 통해 여과하 였다. 레몬그라스 여액은 40℃ 수욕상에서 회전진공농축기 (N-1000, EYELA, Tokyo, Japan)로 감압농축한 후, 동결건조기를 통해 동결 건조하여 총 12.39 g을 획득하여 사용 전까지 -20℃에 보관하였다.
3. HPLC 분석
HPLC 분석기기는 Agilent 1200 시리즈(Agilent Technologies, Santa Clara, CA, USA)를 column은 Shiseido Capcell PRAK MG II C18 (4.6×150 mm, 3 μm), 온도는 35℃로 하였다. 검출기는 Agilent DAD를 사용하였고, wavelength는 254 nm로 하였다. 유속은 0.5 ml/min, 15 μl를 injection volume으 로 설정하였다. 이동상은 0.1% formic acid 수용액(A)과 acetonitrile (B)로 5% B-5 min, 10% B-13 min, 25% B-20 min, 30% B-24 min, 35% B-28 min, 45% B-35 min, 50%
B-40 min, 55% B-43 min, 60% B-50 min, 70% B-56 min까지 gradient 조건으로 분석하였다. 표준물질은 caffeic acid와 isoorientin을 사용하였으며, 각각의 표준물질의 정성 및 정량은 retention time과 고유의 표준폼과 비교하여 검량선으로부터 DLLEM와 DLLFEA 분획물 및 silica column chromatography를 이용하여 분획의 함량 결과를 얻었다.
4. 세포배양 및 분화유도
3T3-L1 지방전구세포는 American Type Culture Collection (ATCC, Rockville, MD, USA)로 부터 분양 받았고, 10% FCS (Invitrogen, Burlington, ON, Canada), 100 U/ml penicillin, 100 μg/ml streptomycin이 포함된 DMEM 배지(Invitrogen, Burlington, ON, Canada)를 사용하여 37℃와 5% CO2 상태에서 배양하였다. 3T3-L1 지방전구세포의 분화를 유도하기 위하여 세포 는 1.5×105 cells/ml로 분주하여 세포가 confluent 상태가 된 2일 후(Day 0), MDI용액(IBMX, dexamethasone, insulin)과 10% FBS (Invitrogen, Burlington, ON, Canada)를 포함하는 DMEM 배지 에서 3일간 배양함으로써 분화를 개시하였다(Day 3). 지방세포로의 완전 분화를 촉진하기 위해 2일에 한번씩 insulin과 10% FBS를 포함하는 DMEM 배지로 교체하여 4일간 배양하였다(Day 7).
5. 세포생존 및 독성검사
레몬그라스 에탄올 추출물에 대한 세포의 생존율을 측정하기 위해 MTT 방법을 이용하여 측정하였다. 3T3-L1 세포는 48 well 플레이트에 24시간 전배양 후, 레몬그라스 에탄올 추출물을 50, 100, 200, 500, 1000 μg/ml 농도로 24 또는 48시간 처리하였다.
0.5 mg/ml MTT용액 500 μl를 각각의 well에 첨가하여 37℃에서
4시간 배양한 후에, 상층액을 제거하고 비수용성의
MTT-formazan 결정들은 DMSO용액 200 μl 첨가하여 용해하였 다. Formazan의 양은 SpectraMAX 250 microplate spectrophotometer (Molecular Devices, Sunnyvale, CA)를 이 용하여 570 nm에서 흡광도를 측정하였다. 세포생존율은 다음과 같 이 계산하여 백분율(%)로 나타내었다. Cell viability (%) = (OD570
of treated cells/OD570 of control cells)×100.
레몬그라스 에탄올 추출물에 대한 세포독성을 측정하기 위해 LDH cytotoxicity assay kit (Pierce, Rockford, IL, USA)를 이 용하여 측정하였다. 3T3-L1 세포는 48 well 플레이트에 24시간 전 배양 후, 레몬그라스 에탄올 추출물을 50, 100, 200, 500, 1000 μ g/ml 농도로 24 또는 48시간 처리하였다. 제조사의 방법에 따라 각각의 샘플배지 50 μl와 Reaction Mixture 용액 50 μl를 혼합하
여 암실조건에서 30분간 방치시킨 후, 50 μl의 Stop 용액을 첨가 하여 반응을 종료하였다. 그 후 SpectraMAX 250 microplate spectrophotometer (Molecular Devices, Sunnyvale, CA)를 이 용하여 490 nm와 680 nm에서 흡광도를 측정하였다. 세포독성은 다음과 같이 계산하여 백분율(%)로 나타내었다. Cytotoxicity (%)
= [(OD490-680 of treated-cells - OD490-680 of spontaneous control cells)/(OD490-680 of maximum control cells - OD490-680 of spontaneous control cells)] ×100.
6. Oil Red O 염색 및 정량
세포내 지방구 생성을 확인하기 위하여 Adipogenesis assay kit (Cayman Chemical, Ann Arbor, MI, USA)를 이용하여 Oil Red O 염색을 실시하였다. 3T3-L1 세포는 12 well 플레이트에 48시간 전배양 후, MDI배지에서 레몬그라스 에탄올 추출물을 50, 100, 200 μg/ml 농도로 처리하였다. 제조사의 방법에 따라서 배지 를 제거한 세포에 1X Fixative 용액을 첨가하여 15분간 고정시킨 후, Wash 용액으로 2회 세척하여 Oil Red O 용액을 첨가하여 20 분간 염색하였다. 그 후 염색액을 제거하고 증류수로 1회 세척한 다음 Wash 용액으로 2회 세척하여 염색된 세포를 광학현미경 (IX71, Olympus, Tokyo, Japan)으로 관찰하였다. 또한 정량을 위 해 Dye extraction 용액을 첨가하여 지방을 추출한 후 SpectraMAX 250 microplate spectrophotometer (Molecular Devices, Sunnyvale, CA)를 이용하여 490 nm에서 흡광도를 측 정하였다. 지방함량은 다음과 같이 계산하여 백분율(%)로 나타내었 다. Lipid contents (%) = (OD490 of treated cells/OD490 of control cells)×100.
7. 세포주기 분석
세포주기의 비율은 PI 염색을 통하여 세포주기를 분석하였다.
세포는 모두 수확하여 PBS (pH 7.4)로 1회 세척하였고, 세척된 세 포는 냉각된 70% 에탄올로 4℃에서 1시간 고정하였다. 고정된 세 포는 PBS (pH 7.4)로 2회 세척한 후, 1 ml의 PI/RNase A 혼합용 액으로 재 부유시켜 37℃에서 암실조건으로 1시간 배양하였다.
DNA 양은 FACS-Calibur (BD Biosciences, CA, USA)를 사용하 여 측정하였다. 세포주기는 CellQuest Pro (BD Biosciences, CA, USA) 소프트웨어를 이용하여 분석하였다.
8. 단백질발현 분석
세포는 얼음으로 냉장된 PBS (pH 7.4)로 세척하고, 1% halt protease&phosphatase inhibitor cocktail이 포함된 RIPA lysis buffer로 부유시킨 후 얼음 위에서 30분간 용해하였다. 세포용해물 들은 4℃에서 14,000 rpm으로 15분간 원심분리하였고, 단백질 농 도는 Bradford 검사법을 이용하여 측정하였다. 총 40 μg의 단백질 샘플을 SDS-PAGE 젤로 분리하였고, 40V에서 3시간 nitrocellulose막 위로 단백질을 전이시켰다. 단백질이 전이된 막은 5% skim milk가 포함된 Tris-buffered saline+Tween-20 (20 mM Tris-HCl, pH 7.6, 150 mM NaCl, 0.05% Tween-20)으로 비특이 결합을 차단시킨 후, 1차 항체(1:1000)를 결합시켰다. 면역
활성은 peroxidase가 붙어있는 anti-rabbit 또는 anti-mouse 2차 항체(1:2000)를 사용하여 SuperSignal West Pico Chemiluminescent (Pierce, Rockford, IL, USA)에 의해 탐지하 였고, FluorChem E System (ProteinSimple, San Jose, CA, USA)을 이용하여 이미지화 하였다.
9. 통계처리
모든 데이터의 결과는 그래프패드 프리즘 6(La Jolla, CA) 프 로그램을 통하여 통계처리 하여 mean±standard deviation (SD) 로 기록하였다. 실험군간의 유의성 검정은 Student’s t-test를 통 해 분석하였으며, p<0.05의 경우에만 통계적 유의성이 있는 것으로 간주하였다.
결 과
1. 레몬그라스 에탄올 추출물의 HPLC 분석
폴리페놀 화합물인 caffeic acid와 isoorientin은 레몬그라스 의 함유되어 있는 성분 중의 하나이다21). 시료의 분석은 HPLC를 이용하였으며 각 시료의 표준용액과 레몬그라스 에탄올 추출물에서 검출된 표준성분인 caffeic acid와 isoorientin의 chromatogram 결과는 Fig. 1에서 나타낸 바와 같다. 또한 표준성분인 caffeic acid 와 isoorientin의 검량선을 확인한 결과 레몬그라스 에탄올 추출물 의 caffeic acid 함량은 3.07 μg/mg dry wt. 이였고, isoorientin 의 함량은 55.44 μg/mg dry wt. 으로 검출되었다(Table 1).
A
B
Fig. 1. Chromatograms of the major compound in Cymbopogon citratus ethanol extracts. (A) HPLC chromatogram of caffeic acid standard and Cymbopogon citratus, and (B) isoorientin standard and Cymbopogon citratus.
Table 1. Caffeic acid and isoorientin contents in Cymbopogon citratus extracts
Extract Caffeic acid content (μ
g/mg dry wt.) Isoorientin content (μ g/mg dry wt.)
EtOH extract 3.07 55.44
2. 레몬그라스 에탄올 추출물의 3T3-L1 지방전구세포에서 세포독 성 효과
레몬그라스 에탄올 추출물을 3T3-L1 세포에 여러 농도(50, 100, 200, 500, 1000 μg/ml)로 24시간과 48시간 처리한 후 세포 생존율을 측정하기 위해 MTT 환원 검사를 실시하였고, 세포독성을 측정하기 위해 LDH 활성 검사를 실시하였다. 레몬그라스 에탄올 추출물에 대한 3T3-L1 세포의 생존율은 농도 의존적으로 감소하였 다(Fig. 2A). 레몬그라스 에탄올 추출물을 24시간 처리하였을 경우 1000 μg/ml 농도에서 세포생존율이 78.63%로 유의적인 감소를 보였으며, 48시간 처리하였을 경우 500 μg/ml과 1000 μg/ml 농 도에서 세포생존율이 각각 80.6%와 57.5%로 유의적인 감소를 보 였다. 레몬그라스 에탄올 추출물에 대한 세포독성은 세포생존율 감 소와 비례하여 농도 의존적으로 증가하였다(Fig. 2B). 레몬그라스 에탄올 추출물을 24시간 처리하였을 경우 1000 μg/ml 농도에서 LDH 활성이 4.8%로 유의적인 증가를 보였으며, 48시간 처리하였 을 경우 200 μg/ml, 500 μg/ml, 1000 μg/ml 농도에서 각각 6.0%, 7.3%, 9.1%로 유의적인 증가를 보였다.
A
B
Fig. 2. Effects of Cymbopogon citratus ethanol extracts on cell viability and cytotoxicity of 3T3-L1 preadipocytes. The cells were treated with 50, 100, 200, 500 and 1000 μg/ml Cymbopogon citratus ethanol extracts for 24 and 48 hr. (A) Cell viability was measured by MTT reduction assay (B) Cytotoxicity was measured by LDH activity assay.
Values are means ± SD, N = 3. The statistically significant differences compared with non-treated cells were calculated by Student’s t-test.
*p<0.05.
3. 레몬그라스 에탄올 추출물의 3T3-L1 지방전구세포에 대한 지방 생성 억제효과
레몬그라스 에탄올 추출물의 지방세포분화 억제활성을 평가하 기 위해, 3T3-L1 세포 내 지방축적량을 Oil Red O 염색을 통해 현미경 관찰과 정량분석 하였다(Fig. 3). 분화시키지 않은 3T3-L1 세포에서는 지방구(Lipid droplet)가 형성이 되지 않았으며 분화를 유도한 대조군 세포에서는 다량의 지방구가 형성된 것을 관찰하였 다. 그러나 레몬그라스 에탄올 추출물을 처리하였을 경우 100 μ g/ml과 200 μg/ml 농도에서 현저한 지방구 감소를 관찰할 수 있 었다(Fig. 3A). 또한 레몬그라스 에탄올 추출물의 3T3-L1 세포 내 지방구 생성 정도를 정량적으로 비교분석하였을 경우 100 μg/ml 과 200 μg/ml 농도에서 각각 37.3%와 22.3%로 유의적인 감소를 보였다(Fig. 3B).
A
B
Fig. 3. Effects of Cymbopogon citratus ethanol extracts on lipid droplet accumulation of 3T3-L1 adipocytes. The cells were differentiated with MDI in the absence of presence of Cymbopogon citratus ethaol extracts (CCEE; 50, 100, 200 μg/ml) for 7 days and then lipid contents were measure by Oil Red O staining. (A) Lipid droplet accumulation was visualized by a light microsope. Magnification, ×200. (B) The lipid contents was quantified by measuring the absorbance at 490 nm. Values are means ± SD, N = 3. The statistically significant differences compared with non-treated cells were calculated by Student’s t-test.
*p<0.05.
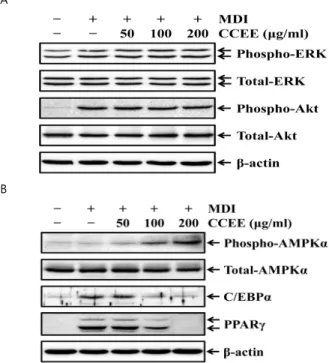
4. 레몬그라스 에탄올 추출물이 3T3-L1 지방전구세포 내 ERK, Akt, AMPK 활성 및 Adipogenic transcription factor들의 발현 에 미치는 효과
호르몬 유도제인 IBMX, dexamethasome, insulin은 3T3-L1 세포가 지방세포로 분화하는 과정 중에 여러 개의 신호전달기전을 활성화 시킨다. 그 중에서 insulin은 지방세포분화를 개시하기 위 한 ERK와 Akt 신호기전을 활성화 시킨다7,8). 따라서 레몬그라스 에탄올 추출물에 의한 3T3-L1 세포의 지방세포 분화억제가 ERK 및 Akt 활성에 미치는 효과를 조사하였다. 호르몬 유도제(MDI)에
의해 분화를 개시한 3T3-L1 세포에서 ERK와 Akt의 인산화 수준 이 증가되는 것을 관찰할 수 있었다. 그러나 레몬그라스 에탄올 추 출물을 처리하였을 경우 ERK와 Akt의 인산화 수준은 MDI만 처리 한 세포와 비교하여 명확한 차이를 보이지 않았다(Fig. 4A).
지방세포분화는 C/EBPα 및 PPARγ와 같은 특정한 전사인자 에 의해 조절된다9,10). 또한 세포의 에너지 대사에 있어 중요한 조 절인자인 adenosine monophosphate-activated protein kinase (AMPK)는 지방세포분화를 조절하는데 관여한다22). 따라서 레몬그 라스 에탄올 추출물에 의한 3T3-L1 세포의 지방세포 분화억제가 C/EBPα 및 PPARγ의 발현과 AMPK의 활성에 미치는 효과를 조사 하였다. 지방세포로 분화되고 있는 3T3-L1 세포에서 C/EBPα와 PPARγ의 발현은 명확하게 증가하였으나, AMPK의 인산화 수준은 분화를 유도하지 않은 세포와 비교하여 차이가 없었다. 그러나 레 몬그라스 에탄올 추출물을 처리하였을 경우 C/EBPα와 PPARγ의 발현이 MDI만 처리한 세포와 비교하여 현저하게 감소되는 것을 관 찰할 수 있었으며, 또한 AMPK의 인산화 수준이 증가하는 것을 관 찰할 수 있었다(Fig. 4B).
A
B
Fig. 4. Effects of Cymbopogon citratus ethanol extracts on phospholyation of ERK, Akt and AMPKα, and adopogenic transcription factor in 3T3-L1 adipocytes. The cells were differentiated with MDI in the absence of presence of Cymbopogon citratus ethanol extracts (CCEE; 50, 100, 200 μg/ml) for 30 min (A) or 7 days (B) and then Whole cell lysates were subjected to SDS-PAGE followed by Western blot analysis with anti-ERK, anti-phospho-ERK, anti-Akt, anti-phospho-Akt, anti-AMPKα, anti-phospho-AMPKα, anti-C/EBPα and PPARγ antibodies.
5. 레몬그라스 에탄올 추출물이 3T3-L1세포의 세포주기 진행 및 조절 관련 단백질들의 발현에 미치는 효과
Post-confluent에 의해 성장이 정지된 3T3-L1 지방전구세포 를 지방세포로의 분화를 유도하게 되면 mitotic clonal expansion (MCE)을 위해 세포주기진행이 개시된다23). 따라서 레몬그라스 에 탄올 추출물이 지방세포 분화를 유도한 3T3-L1 세포의 세포주기진
행과 세포주기 조절과 관련된 단백질들의 발현에 미치는 효과를 조 사하였다. 지방세포 분화를 유도하지 않은 3T3-L1 세포에서 G0/G1기는 66.05%, S기는 15.91%, G2/M기는 12.99%로 관찰되 었으나, MDI에 의해 지방세포 분화를 유도한 3T3-L1 세포는 G0/G1기가 40.17%로 감소하였고 S기와 G2/M기는 각각 35.13%
와 19.57%로 증가하였다. 그러나 레몬그라스 에탄올 추출물을 처 리하였을 경우 MDI만 처리한 3T3-L1 세포와 비교하여 G0/G1기는 53.76%로 증가하였고, S기와 G2/M기는 각각 25.09%와 16.05%로 감소하였다(Fig. 5A). 세포주기 조절과 관련된 단백질들의 발현분석 에서 레몬그라스 에탄올 추출물에 의해 p21과 p27의 발현은 증가 되었고, CDK2, cyclin A, cyclin B1의 발현을 감소되었으나 cyclin E의 발현에는 영향이 없었다(Fig. 5B).
A
B
Fig. 5. Effects of Cymbopogon citratus ethanol extracts on cell cycle progression and expression of cell cycle-related protein in 3T3-L1 adipocytes. The cells were differentiated with MDI in the absence of presence of Cymbopogon citratus ethanol extracts (CCEE; 50, 100, 200 μg/ml) for 2 days. (A) The cells were fixed and stained with PI and cell cycle progression was analyzed by flow cytometry. (B) Whole cell lysates were subjected to SDS-PAGE followed by Western blot analysis with anti-p21, anti-p27, anti-CDK2, anti-cyclin A, anti-cyclin B1 and anti-cyclin E antibodies.
고 찰
3T3-L1 세포는 비만 연구에 가장 널리 사용되는 지방전구 세 포주이다24). 3T3-L1 지방전구세포는 호르몬유도제인 IBMX, dexamethasone, insulin에 의해 지방세포로 분화되면서 세포 내 지방을 축적한다24). 지방전구세포가 지방세포로 전환되는 과정인 adipogenesis는 비만과 밀접한 관련이 있으며 이를 효과적으로 조 절하는 것은 비만을 예방하는데 있어 중요한 부분이다24). 지방세포
분화는 C/EBP 패밀리와 PPARγ와 같은 특이적 전사인자에 의해 조절되며, 이 전사인자들은 다양한 지방생성 단백질들의 발현에 관
여한다25,26). C/EBP 패밀리 중에서도 α, β, δ 형태가 adipogenesis
와 밀접하게 관련된 것으로 알려져 있으며, 특히 C/EBPβ와 C/EBPδ는 분화 초기에 발현되어 adipogenesis를 조절하는 것으로 알려져 있다11). 또한 C/EBPα와 PPARγ의 발현은 adipogenesis를 진행하는데 있어 매우 중요하게 작용하는 전사인자로 지방축적과 insulin 감수성에 관여하는 다수의 유전자들의 발현을 활성화 시킨 다7). 또한 AMPK는 에너지 대사과정에서 중요한 효소로 생체 내에 서 포도당 수치와 지질흡수를 조절하는데 관여한다22). AMPK의 인 산화는 지방산과 콜레스테롤 합성에 관여하는 대사효소들의 활성을
억제한다27-29). 특히 AMPK경로는 C/EBPα와 PPARγ의 상위신호기
전으로 AMPK가 활성화 되면 지방세포분화를 억제하고 세포 내 지 방산과 콜레스테롤을 감소시킨다30-32). 레몬그라스 에탄올 추출물은 3T3-L1 지방전구세포의 지방세포분화에 의한 세포 내 지방축적을 감소시켰으며, 지방축적에 관여하는 유전자들의 전사인자인 C/EBP α와 PPARγ의 발현 억제뿐만 아니라 상위신호인 AMPK의 활성도 감소시켰다. 이 결과들은 레몬그라스 에탄올 추출물에 의한 3T3-L1 지방전구세포의 apdipogenesis 억제효과가 C/EBPα와 PPARγ의 발현감소를 통하여 이루어졌으며, C/EBPα와 PPARγ의 발현감소는 AMPK의 활성증가와 관련되어 있다는 것을 보여주고 있다.
ERK 활성과 PI3K/Akt 경로는 adipogenesis에 필요한 세포신 호기전이다7,33). ERK 활성은 지방전구세포의 분화개시를 위해 요구 되며 지방세포분화의 초기단계에서 증가하는데 ERK 경로를 억제할 경우 adipogenesis가 감소되는 것으로 보고되었다34,35). 또한 PI3K/Akt 경로도 지방세포의 분화에 관여하며 지방세포분화시 Akt 의 활성이 억제되면 adipogenesis가 감소되는 것으로 보고되었다
36,37). 그러나 본 연구에서 레몬그라스 에탄올 추출물은 3T3-L1 지
방전구세포의 분화에 의해 증가된 ERK 및 Akt 활성을 억제하지 못하였다. 이 결과들은 레몬그라스 에탄올 추출물이 3T3-L1 지방 전구세포의 분화 초기단계에서 증가된 ERK와 Akt 활성에는 관여 하지 않는다는 것을 보여주고 있으며, 레몬그라스 에탄올 추출물의 지방세포분화 억제 효과가 지방전구세포의 분화초기 단계에 관여하 는 세포신호기전과는 연관성이 없었다.
지방생성 과정은 지방전구세포의 증식과 성숙한 지방세포로의 분화로 이루어져 있으며, 증식은 aidpogenesis의 필수조건이다38). 성장이 정지된 3T3-L1 지방전구세포가 지방세포분화를 개시하게 되면 MCE를 위해 세포주기진행으로 다시 들어간 후, 지방세포 특 이적 유전자의 발현을 유도한다6,38). Cyclin-dependent kinases (CDKs)는 세포주기를 진행하게 하는 효소로, cyclin과 복합체를 이 루어 세포주기진행을 조절하며, 이 Cyclin-CDK 복합체의 형성은 cyclin-dependent kinase inhibitors (CDKIs)에 의해 제어된다39). 레몬그라스 에탄올 추출물은 지방세포분화가 개시된 3T3-L1 세포 의 세포주기를 G0/G1기에서 정지시켰으며, CDK 활성을 억제하는 p21과 p27의 발현을 증가시키고, CDK2, cyclin A, cyclin B1의 발현을 감소시켰다. 이 결과들은 레몬그라스 에탄올 추출물이 3T3-L1 지방세포에서 p21과 p27의 발현증가를 통해 G0/G1기에
서 세포주기정지를 유도한다는 것을 보여주고 있다.
결 론
비만은 체내 지방이 과도하게 축적되어 일어나는 현상으로 당 뇨, 고혈압, 심혈관 질환 및 암과 같은 질병의 원인이 된다. 본 연 구는 레몬그라스 에탄올 추출물의 항비만 효과에 대한 연구를 위해 3T3-L1 지방전구세포에서 지방분화에 미치는 영향을 조사하였다.
레몬그라스 에탄올 추출물은 3T3-L1 지방전구세포에서 지방구의 형성을 농도 의존적으로 억제시켰다. 이는 C/EBPα와 PPARγ 등의 지방세포분화 전사인자들의 단백질 발현감소와 이들의 발현을 음성 적으로 조절하는 AMPK의 활성이 증가하면서 지방구의 축적이 억 제된 것으로 나타났다. 또한 레몬그라스 에탄올 추출물은 세포주기 정지를 통해 지방세포 분화개시에 의한 3T3-L1 지방전구세포의 MCE 유도를 저해하여 adipogenesis를 효과적으로 억제하였다. 따 라서 레몬그라스는 항비만 소재로써 활용 가능성이 있을 것으로 사 료된다.
감사의 글
이 논문은 2017학년도 원광대학교의 교비지원에 의해 수행됨
References
1. Peng Y, Yu S, Li H, Xiang H, Peng J, Jiang S.
MicroRNAs: Emerging roles in adipogenesis and obesity.
Cell Signal. 2014;26(9):1888-96.
2. Jo J, Gavrilova O, Pack S, Jou W, Mullen S, Sumner AE, Cushman SW, Periwal V. Hypertrophy and/or hyperplasia: dynamics of adipose tissue growth. PLoS Comput Biol. 2009;5(3):e1000324.
3. Oh JS, Kim JS, Lee BM, Lee SK, Chung MY. A Comparative Study on Obesity-related Eating Habits and Lifestyle Behaviors of Depressed and Normal Middle-aged Female in Korea. Kor J Aesthet Cosmetol.
2013;11(2):231-8.
4. Chung MY, Kim SH, Oh JS. Comparison of obesity-related life patterns with according to body mass index and self-awareness of body shape. Kor J Aesthet Cosmetol. 2013;11(6):1109-1115.
5. Gwon SY, Ahn JY, Jung CH, Moon BK, Ha TY. Shikonin suppresses ERK 1/2 phosphorylation during the early stages of adipocyte differentiation in 3T3-L1 cells. BMC Complem Altern Med. 2013;13:207.
6. Cornelius P, MacDougald OA, Lane MD. Regulation of adipocyte development. Annu Rev Nutr. 1994;14:99-129.
7. Prusty D, Park BH, Davis KE, Farmer SR. Activation of MEK/ERK signaling promotes adipogenesis by enhancing
peroxisome proliferator-activated receptor gamma (PPARgamma ) and C/EBPalpha gene expression during the differentiation of 3T3-L1 preadipocytes. J Biol Chem.
2002;277(48):46226-32.
8. Xu J, Liao K. Protein kinase B/AKT 1 plays a pivotal role in insulin-like growth factor-1 receptor signaling induced 3T3-L1 adipocyte differentiation. J Biol Chem.
2004;279(34):35914-22.
9. Rosen ED, Sarraf P, Troy AE, Bradwin G, Moore K, Milstone DS, Spiegelman BM, Mortensen RM. PPAR gamma is required for the differentiation of adipose tissue in vivo and in vitro. Mol Cell. 1999;4(4):611-7.
10. Wu Z, Rosen ED, Brun R, Hauser S, Adelmant G, Troy AE, McKeon C, Darlington GJ, Spiegelman BM.
Cross-regulation of C/EBP alpha and PPAR gamma controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol Cell. 1999;3(2):151-8.
11. Shon MS, Shin YJ, Lee SC, Kim GN. Anti-obese activities of Kaki-tannins prepared from various persimmons through inhibition of 3T3-L1 adipogenesis.
Kor J Aesthet Cosmetol. 2014;12(4):539-46.
12. Gupta R, Rathi P, Gupta N, Bradoo S. Lipase assays for conventional and molecular screening: an overview.
Biotechnol Appl Biochem. 2003;37(Pt 1):63-71.
13. Kim SH, Kim JY, Ryu KA, Sohn CM. Evaluation of the dietary diversity and nutrient intakes in obese adults.
Korean J Community Nutr. 2007;12(5):583-91.
14. Yi MR, Kang CH, Bu HJ. Acetic acid fermentation properties and antioxidant activity of lemongrass vinegar. Korean J Food Preserv. 2017;24(5):680-7.
15. Bhattacharya AK, Kaul PN, Rajeswara Rao BR. Effect of Prolonged Storage on the Quality of Lemongrass (Cymbopogon flexuosus (Nees ex Steud.) Wats.) Essential Oil. J Essent Oil Bear Pl. 1998;1(2-3):104-9.
16. Sforcin JM, Amaral JT, Fernandes A Jr, Sousa JP, Bastos JK. Lemongrass effects on IL-1beta and IL-6 production by macrophages. Nat Prod Res.
2009;23(12):1151-9.
17. Francisco V, Figueirinha A, Costa G, Liberal J, Lopes MC, Carmen GR, Carlos FG, Cruz MT, Batista MT.
Chemical characterization and anti-inflammatory activity of luteolin glycosides isolated lemongrass. J Funct Foods. 2014;10:436-43.
18. Silva CB, Guterres SS, Weisheimer V, Schapoval ES.
Antifungal activity of the lemongrass oil and citral against Candida spp. Braz J Infect Dis. 2008;12(1):63-6.
19. Sri Devi S, Ashokkumar N. Citral, a Monoterpene Inhibits Adipogenesis Through Modulation of Adipogenic
Transcription Factors in 3T3-L1 Cells. Indian J Clin Biochem. 2018;33(4):414-21.
20. Modak T, Mukhopadhaya A. Effects of citral, a naturally occurring antiadipogenic molecule, on an energy-intense diet model of obesity. Indian J Pharmacol. 2011, 43(3):300-5.
21. Cheel J, Theoduloz C, Rodríguez J, Schmeda-Hirschmann G. Free radical scavengers and antioxidants from Lemongrass (Cymbopogon citratus (DC.) Stapf.). J Agric Food Chem. 2005;53(7):2511-7.
22. Lee MS, Cho SM, Lee MH, Lee EO, Kim SH, Lee HJ.
Ethanol extract of Pinus koraiensis leaves containing lambertianic acid exerts anti-obesity and hypolipidemic effects by activating adenosine monophosphate-activated protein kinase (AMPK). BMC Complement Altern Med. 2016;16:51.
23. Tang QQ, Otto TC, Lane MD. Mitotic clonal expansion:
a synchronous process required for adipogenesis. Proc Natl Acad Sci USA. 2003;100(1):44-9.
24. Shon MS, Kim GN. Anti-oxidant and anti-obese activities of turmeric (Curcuma longa L.) extract in 3T3-L1 Cells. Kor J Aesthet Cosmetol. 2014;12(2):169-5.
25. Farmer SR (2006) Transcriptional control of adipocyte formation. Cell Metab. 2006;4(4):263-3.
26. Tontonoz P, Hu E, Spiegelman BM. Stimulation of adipogenesis in fibroblasts by PPAR gamma 2, a lipid-activated transcription factor. Cell.
1994;79(7):1147-56.
27. Carling D, Zammit VA, Hardie DG. A common bicyclic protein kinase cascade inactivates the regulatory enzymes of fatty acid and cholesterol biosynthesis.
FEBS Lett. 1987;223(2):217-22.
28. Hardie DG, Corton J, Ching YP, Davies SP, Hawley S.
Regulation of lipid metabolism by the AMP-activated protein kinase. Biochem Soc Trans. 1997;25(4):1229-31.
29. Hardie DG, Pan DA. Regulation of fatty acid synthesis and oxidation by the AMP-activated protein kinase.
Biochem Soc Trans. 2002;30(Pt 6):1064-70.
30. Chen S, Li Z, Li W, Shan Z, Zhu W. Resveratrol inhibits cell differentiation in 3T3-L1 adipocytes via activation of AMPK. Can J Physiol Pharmacol.
2011;89(11):793-9.
31. Lee YK, Lee WS, Hwang JT, Kwon DY, Surh YJ, Park OJ. Curcumin exerts antidifferentiation effect through AMPKalpha-PPAR-gamma in 3T3-L1 adipocytes and antiproliferatory effect through AMPKalpha-COX-2 in cancer cells. J Agric Food Chem. 2009;57(1): 305-10.
32. Rosen ED, Spiegelman BM. PPARgamma : a nuclear
regulator of metabolism, differentiation, and cell growth. J Biol Chem. 2001;276(41):37731-4.
33. Rosen ED, MacDougald OA. Adipocyte differentiation from the inside out. Nat Rev Mol Cell Biol.
2006;7(12):885-96.
34. Smith PJ, Wise LS, Berkowitz R, Wan C, Rubin CS.
Insulin‑like growth factor‑I is an essential regulator of the differentiation of 3T3‑L1 adipocytes. J Biol Chem.
1988;263(19):9402-8.
35. Bost F, Aouadi M, Caron L and Binétruy B. The role of MAPKs in adipocyte differentiation and obesity.
Biochimie 2005;87(1):51-6.
36. Magun R, Burgering BM, Coffer PJ, Pardasani D, Lin Y, Chabot J, Sorisky A. Expression of a constitutively activated form of protein kinase B (c‑Akt) in 3T3‑L1 preadipose cells causes spontaneous differentiation.
Endocrinology. 1996;137(8):3590-3.
37. Peng XD, Xu PZ, Chen ML, Hahn‑Windgassen A, Skeen J, Jacobs J, Sundararajan D, Chen WS, Crawford SE, Coleman KG, Hay N. Dwarfism, impaired skin development, skeletal muscle atrophy, delayed bone development and impeded adipogenesis in mice lacking Akt1 and Akt2. Genes Dev. 2003;17(11):1352-65.
38. Kim SH, Park HS, Lee MS, Cho YJ, Kim YS, Hwang JT, Sung MJ, Kim MS, Kwon DY. Vitisin A inhibits adipocyte differentiation through cell cycle arrest in 3T3-L1 cells.
Biochem Biophys Res Commun. 2008;372(1):108-13.
39. Jang YJ, Koo HJ, Sohn EH, Kang SC, Rhee DK, Pyo S.
Theobromine inhibits differentiation of 3T3-L1 cells during the early stage of adipogenesis via AMPK and MAPK signaling pathways. Food Funct.
2015;6(7):2365-74.