Original Article
원고 접수일 2013년 2월 20일, 원고 수정일 2013년 3월 14일, 게재 확정일 2013년 5월 14일
책임저자 김욱규
(626-870) 양산시 물금읍 부산대학로 49, 부산대학교 치의학전문대학원 구강악안면 외과학교실
Tel: 055-360-5112, Fax: 055-360-5104, E-mail: [email protected]
RECEIVED February 20, 2013, REVISED March 14, 2013, ACCEPTED May 14, 2013
Correspondence to Uk-Kyu Kim
Department of Oral and Maxillofacial Surgery, School of Dentistry, Pusan National University
49 Busandaehak-ro, Mulgeum-eup, Yangsan 626-870, Korea Tel: 82-55-360-5112, Fax: 82-55-360-5104, E-mail: [email protected]
CC This is an open access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/
by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
인테그린 α
2와 상피성장인자수용체 차단항체의 저해작용을 통한 구강편평상피암 세포의 선택적 제거
최연식ㆍ김규천1ㆍ윤 식2ㆍ황대석ㆍ김철훈3ㆍ전영찬4ㆍ변준호5ㆍ신상훈ㆍ김욱규
부산대학교 치의학전문대학원 구강악안면외과학교실, 1구강해부학교실, 4보철학교실, 2부산대학교 의학전문대학원 해부학교실,
3동아대학교 의과대학 구강악안면외과학교실, 5경상대학교 의학전문대학원 구강악안면외과학교실
Abstract
Induction of Selective Cell Death of Oral Squamous Carcinoma Cells by Integrin α
2Antibody and EGFR Antibody
Yeon-Sik Choi, Gyoo-Cheon Kim1, Sik Yoon2, Dae-Seok Hwang, Cheol-Hun Kim3, Young-Chan Jeon4, June-Ho Byun5, Sang-Hun Shin, Uk-Kyu Kim
Departments of Oral and Maxillofacial Surgery, 1Oral Anatomy, 4Prothodontics, School of Dentistry, Pusan National University,
2Department of Anatomy, School of Medicine, Pusan National University, 3Department of Oral and Maxillofacial Surgery, College of Medicine, Dong-A University, 5Department of Oral and Maxillofacial Surgery, School of Medicine, Gyeongsang National University
Purpose: This study was to find efficacy of integrin alpha2 (α2) and epidermal growth factor receptor (EGFR) as tumor marker of oral squamous cell carcinoma (SCC) and clarify the selective cell death effect of anti-integrin α2 and anti-EGFR on SCC cells, additionally testify conjugated gold nanoparticles (GNP) with air plasma for selective cell death of oral SCC.
Methods: Expression of integrin α2, EGFR on human SCC cells (SCC25) were examined by western blot. SCC25 cells were treated with anti-integrin α2, anti-EGFR and analysed by Hemacolor staining, immunoflorescence staining, FACS flow cytometry.
Conjugated GNP with integrin α2, EGFR antibody were treated by air plasma on SCC cells.
Results: Integrin α2 and EGFR were over-expressed on SCC25 cells than normal lung WI-38 cells. The cell viability rate of SCC25 cells treated with anti-integrin α2, anti-EGFR was lower than WI-38 cells. The concentration changes of nucleus, releasing cytochrome c and apoptosis inducing factor (AIF) from mitochondria to cytosol were observed. The changes of proteins related with apoptosis were observed. Increase of bax, bcl-xL, activation of caspase-3, -7, -9, and fragmentation of PARP, DFF45 and decrease of lamin A/C in SCC25 cells were observed. In FACS, increase of sub-G1 and S phase was observed. Cell cycle related proteins, Such as cyclin D1, cyclin dependent kinase (CDK) 4, cyclin A, cyclin E, CDK 2, p27 were decreased. After SCC25 cells treated with conjugatged GNP-Integrin α2, GNP-EGFR, additionally air plasma, the cell death rate was significantly increased.
Conclusion: Integrin α2, EGFR were over-expressed in oral SCC cells. Anti-integrin α2, anti-EGFR in SCC25 cells induced apoptosis selectively. When GNP-anti integrin α2, GNP-anti EGFR were treated with air plasma on SCC25 cells, cancer cells were died more selectively. GNP-anti integrin α2, GNP-anti EGFR with air plasma could be treatment choice of oral SCC.
Key words: OSCC, Integrin alpha2, EGFR, Gold nanoparticles, Cell death
서 론
구강편평상피암(oral squamous cell carcinoma)은 구강영역 에서 가장 호발하는 암으로서, 구강점막 상피에서 유래되어 결합조 직으로 침투하는 암이다. 이는, 구강암 중 가장 전이 및 재발률이 높아 장기 생존율이 50% 미만일 정도로 예후가 불량하다. 구강편 평상피암의 치료는 외과적 수술, 화학요법 및 방사선요법이 주로 사용되고 있으나, 치료기술의 많은 발전에도 불구하고 국소적 재발과 전이로 인하여 그 예후가 크게 개선되지 않았다. 이에 보존적, 비외과적 개념의 선택적 암세포 사멸유도 기술의 필요성이 제기되어 왔으며 특정 암세포에서 많이 발현되는 단백질에 대한 항체로 표적인자를 공격함으로써 세포사멸을 유도하는 암치료 접근방식이 대두되었다. 이에 본 연구에서는, 구강편평상피암의 세포에서 과잉 발현되는 단백질들을 분석하여, 이에 대한 차단항체 를 사용하여 구강암세포 사멸 효과를 연구하고자 하였다.
인테그린은 세포부착과 세포 사이의 상호작용에 관여하는 수용 체로서 인테그린 α, β 두 소단위의 이형이합체로 구성되는 막관 통형 당단백질이다. 이는, 세포 내에 생기는 신호전달계의 활성화, 유전자 발현의 조절, 세포골격의 재구축 등에 관여하는 기능적 수용체로 작용하며 특히 암 전이에 중요한 역할을 하는 것으로 알려져 있다[1]. 또한, 최근에는 종양의 증식, 침습, 전이와 항암제 저항성의 유발 같은 세포 내 기능을 중재하는 것으로 알려져 암의 진단과 치료에 대한 효율적인 표적인자로 각광받고 있다[2].
폐의 암종에서 인테그린의 발현에 대한 상관관계를 밝힌 연구가 있으나 아직 구강편평상피암에서 인테그린의 역할에 대해서는 자세히 알려져 있지 않기 때문에, 본 연구에서는 인테그린 α2
작용을 저해함으로써 암세포의 세포사멸 유도효과를 분석하고자 하였다[3].
상피성장인자수용체(epidermal growth factor receptor, EGFR) 는 세포 내 tyrosine kinase 부위와 세포 외 리간드 결합부위를 가지는 막관통형 당단백질이며 다양한 세포의 세포막에 존재한다 [4,5]. 이는, 성장이 빠르게 일어나는 상피세포에서 많이 관찰된 다. EGFR은 종양세포의 증식 및 발암기전에서도 중요한 역할을 하며, 인체 내 여러 부위에서 발생하는 상피성 암종에서 그 발현과 임상적 소견과의 연관성이 연구되었고, 치료결과에 대한 예후 예측이나 재발 가능성 등에 대한 중요한 지표로서 연구되고 있으 며 임상에서도 적용되고 있다[6-8]. 최근 구강편평상피암과 두경 부편평상피암에서 EGFR의 발현 연관성에 대하여 보고된 바 있으 며 gefitinib, cetuximab, erlotinib 등의 약제가 출시되어 임상에 서 사용되고 있다[9,10]. 이에 본 연구에서는 EGFR 차단항체 (blocking antibody)를 이용하여 EGFR의 작용을 저해함으로써 암세포 사멸유도효과를 확인하고 더불어 그 사멸기작을 연구하고 자 하였다[11,12].
아울러 본 연구에서 표적 단백질 항체에 의한 암세포 사멸효과
를 증대시키기 위해 금나노입자(gold nanoparticle, GNP)를 항 체에 결합시켜 중합체를 만든 후 저온플라즈마를 적용하였다.
제 4의 물질상태로 분류되는 플라즈마는 전자, 이온, 래디컬, 광자, 전자기장 등으로 구성된 이온화된 기체이며 살균, 지혈, 상처 치유, 최근에는 암세포의 제거까지 다양한 분야까지 연구가 확대 되고 있다[13]. 최근 저온 플라즈마가 암세포 제거의 중요한 기작 인 세포자멸사(apoptosis)를 유도하는 것이 증명되었다[14-17].
하지만 항암 치료에 있어서는 암세포의 선택적 제거가 중요하나, 플라즈마는 정상세포와 암세포의 선택적 제거가 힘들다는 점이 단점으로 지목되고 있다. 본 연구에서는 이러한 문제를 해결하기 위해서 암세포에서 과잉 발현되는 단백질에 대한 항체와 인체에 무해한 GNP를 중합시켜 암세포의 선택적 제거를 유도하였다.
GNP는 세포독성이 없으며 화학적 안정성을 가지며 생체 적합성 이 뛰어나고, 생물물질과 쉽게 결합하여 각광받고 있다[18,19].
따라서, 본 연구에서는 구강편평상피암 세포에서 과잉 발현된 인테그린 α2와 EGFR이 종양 표적인자로 사용될 수 있는지를 확인하기 위하여 이에 대한 차단항체를 처리하였을 때, 그 생존율 과 사멸기작을 규명하고자 하였다. 또한 그 효율성을 높이기 위해 서 인테그린 α2와 EGFR에 대한 차단항체와 GNP 중합체를 제작 하여 구강편평상피암세포와 결합시킨 후 저온플라즈마를 사용하 여 암세포의 선택적 제거를 유도하여 그 효과를 규명하고자 계획 하였다.
연구방법
1. 연구재료
GNP는 BB International (Cardiff, UK)에서, 그리고 RPMI1640, Dulbecco’s modified eagle’s medium/F12 (DMEM/F12)는 Gibco (Carlsbad, CA, USA)에서, propidium iodide (PI)와 RNase A, protease inhibitor cocktail은 Sigma-Aldrich (St. Louis, MO, USA)에서, 현미경 검사용 Hemacolor는 MERCK (Darmstadt, Germany)로부터 구입해 사용했다. Rabbit polyclonal anti-human cyclin D1 항체는 Cell Signaling Technology Inc. (Boston, MA, USA)에서, rabbit polyclonal anti-human cyclin A, cyclin E, cyclin dependent kinase (CDK) 4, apoptosis inducing factor (AIF), cytochrome c, DFF45 (ICAD), p27 항체들은 Santa Cruz Biotechnology (Santa Cruz, CA, USA)에서, monoclonal mouse anti-human CDK 2, GAPDH, caspase-3, caspase-7, caspase 9, PARP-1, EGFR, 인테그린 α2, bax, bcl-xL, lamin A/C 항체들은 Santa Cruz Biotechnology에서 구입했다.
2. 세포배양
사람설암 세포주인 squamous cell carcinoma 25 (SCC25)와 폐섬유모세포주인 WI38 VA-13 subline 2RA (WI-38)은 ATCC (Rockville, MD, USA)에서 구입하였고, 각각 DMEM/F12 1:1 배지와 RPMI1640 배지에 100 μg/mL penicillin/streptomycin, 4 mM L-glutamine, 10% fetal bovine serum (FBS)를 첨가한 다음 37oC의 5% CO2 배양기에서 배양되었다.
3. WST-1 방법을 이용한 세포 생존율 측정
세포 생존율은 WST-1 방법으로 사용되었다. 96-웰 플레이트 에서 각 웰당 5×103개의 세포를 파종한 다음 24시간 배양하였다.
세포의 배지를 제거한 다음, 인테그린 α2 및 EGFR 차단항체를 24, 48, 72시간 동안 반응시켰다. 그 후 각 웰에 WST-1 용액을 10 μL씩 첨가하고 2시간 동안 배양기에서 반응을 시켰다. 이후, ELISA reader (Tecan, Mannedorf, Switzerland)로 450 nm 파장에서 흡광도를 측정했다. 각각의 분석은 세 번 시행되었다.
4. 세포형태 변화 관찰
3×104개의 세포들을 콜라겐이 코팅된 덮개유리에서 24시간 배양을 한 후 인테그린 α2와 EGFR 차단항체를 72시간 처리했다.
그런 다음 배지를 제거하고 생리식염수(phosphate buffered saline, PBS)로 세척한 후 Hemacolor를 사용해 염색했다. 용액 1, 2, 3 (용액1. fixing solution, 용액2. colour reagent red, 용액3. colour reagent blue)을 차례대로 염색 후 광학현미경 (Carl Zeiss, Goettingen, Germany)에서 세포의 형태변화를 관찰하였다.
5. 세포 내 AIF와 cytochrome c 위치변화 관찰
인테그린 α2와 EGFR 차단항체를 처리한 다음, 배지를 제거하 고, PBS로 세척한 후, cytospin에 500 rpm에서 2분간 원심분리를 한 다음 유리슬라이드에 부착을 시켰다. 4% paraformaldehyde 로 고정하고, 3% bovine serum albumin을 처리해 1시간 동안 blocking을 하고 1차 항체(AIF, cytochrome c)를 처리해 37oC에 서 1시간 동안 반응시켰다. 다시 PBS로 세척하고 2차 항체를 사용해 실온에서 1시간 동안 반응시켰다. 세포들을 공초점현미경 (Zeiss LSM 700 laser-scanning confocal microscope, Carl Zeiss)으로 AIF와 cytochrome c의 위치변화를 관찰하고 분석했다.
6. Western blot 분석을 통한 세포자멸사와 연관된 단백질 들의 발현조사
세포를 35 mm 배양접시에 24시간 배양한 다음, 인테그린 α2와 EGFR 차단항체로 각각의 정해진 시간 동안 처리했다. 4oC 에서 세포를 모은 다음 PBS로 세척했다. 이후 12,000 rpm에
10분 동안 원심분리한 다음 차가운 PBS로 세척했다. 적당량의 lysis buffer (10 mM Tris/HCl, pH 7.2, 1% Tripton X-100, 150 nM NaCl, 5 mM EDTA, protease inhibitor cocktail)를 첨가해 4oC에서 1시간 동안 반응시킨 후, 12,000 rpm에 20분간 원심분리해 상층액에 있는 총 단백질을 분리했다. 단백질의 농도 는 Bio-Rad 단백질 정량시약(Bio-Rad Laboratories, Hercules, CA, USA)을 사용해 정량한 다음 sample buffer를 섞어 sample 을 만들었다. 동량의 sample을 polyacrylamide SDS gel을 이용 해 전기영동으로 분리한 후, PVDF membrane으로 전이시켰다.
5% skim milk를 처리해 1시간 동안 blocking을 하고, 1차 항체 를 처리해 4oC에서 밤새 적용시킨 다음, TBS-T (20 mM Tris, 150 mM NaCl, 0.1% Tween 20)로 세척하고 2차 항체를 사용해 1시간 동안 반응시켰다. 반응이 끝난 다음 TBS-T로 세척하고 ECL western blotting reagent을 적용시킨 다음 PARP-1, DFF45, caspase-3, caspase-7, caspase-9, bax, bcl-xL, lamin A/C의 발현량을 분석했다.
7. 유세포분석기를 이용한 세포주기 분석
세포주기 및 sub-G1기 분석을 위해, SCC25 세포들을 각 웰당 1×104개로 35 mm 배양접시에서 각각 24시간 동안 배양하고 배지를 제거한 다음, 인테그린 α2와 EGFR 차단항체를 각각 처리 했다. 세포들을 모아서 PBS로 세척하고, 10분 동안 2,000 rpm으 로 모은 다음, 70% 알코올에 고정했다. 고정된 각각의 세포들은 다시 PBS로 세척하고 200 μg/mL 농도의 RNase A를 4oC에서 1시간 동안 반응시켰다. 이후, PI 용액을 최종 100 μg/mL로 4oC에서 배양하였다. 유세포분석기(FC500, Beckman Coulter, Brea, CA, USA)에서 분리한 다음 측정했다.
8. 섬유형 액틴(filamentous actin) 염색을 통한 세포질의 변화관찰
3×104개의 세포들을 콜라겐이 코팅된 덮개유리에 24시간 배 양을 한 후 인테그린 α2와 EGFR 차단 항체를 처리했다. 그런 다음 배지를 제거하고 PBS로 세척한 후 4% paraformaldehyde 로 고정하고, 3% bovine serum albumin을 처리해 1시간 동안 blocking을 하고 rhodamine F-actin을 37oC에서 20분 동안 반응시켰다. 세포들을 공초점현미경으로 관찰하고 분석했다.
9. GNP와 인테그린 α2와 EGFR 차단항체의 중합
11-mercaptoundecanoic acid (MUA) 0.1 mg/mL 농도의 시약을 30 nm의 colloidal GNP 용액에 밤새 적용시켰다. 그런 다음 1 mM N-hydroxysuccinimide (NHS)와 1 mM N-eth- yl-N’-(3-dimethylaminopropyl) carbodiimide (EDC) 용액을 20분간 4oC에서 처리하였다. 이후, 250 μg/mL 인테그린 α2와 EGFR 차단항체를 PBS (1 mM, pH 7.0)에서 1시간 동안 처리해
Fig. 1. The expression of integrin α2 and EGFR proteins in WI-38 cells and SCC25 cells by western blot assay. The total protein lysates were used for SDS-PAGE followed by western blot using antibodies against integrin α2 and EGFR proteins. SCC, squamous cell carcinoma; EGFR, epidermal growth factor receptor.
Fig. 2. Inhibition of cell proliferation by antibodies. The viability of SCC25 cells and WI-38 cells was measured using a WST-1 assay after treatment of anti-integrin α2 antibody (A) and anti-EGFR antibody (B). EGFR, epidermal growth factor receptor; SCC, squamous cell carcinoma.
반응시켰다.
10. 중합체에 대한 저온 플라즈마 처리
4×104개의 세포들을 콜라겐이 코팅된 덮개유리에 파종하고 6시간 배양을 한 후 GNP-인테그린 α2와 GNP-EGFR 차단항체를 24시간 처리했다. 세포들을 FBS가 없는 배양액으로 세척하고 플라즈마 장치와 2 mm 간격에서 30초간 조사하였다(전압 554.9 Vrms, 전류 10.19 mArms, 평균소비전력 5.654 W, 30초간 면적 당 소비 에너지 16.56 J/cm2).
11. Trypan blue를 이용한 세포생존율 측정
플라즈마를 처리 후 PBS로 세척을 한 후 trypsin-EDTA에 5분간 37oC에서 적용하였다. 떨어진 세포들을 모은 다음 trypan blue를 5분간 적용하고 hemacytometer를 사용하여 죽은 세포 들을 측정하였다.
12. 통계분석
실험군과 대조군의 결과는 통계프로그램인 SPSS (Window 12.0, SPSS Inc., Chicago, IL, USA)에 의해 paired t-test를 적용해 분석했다. P<0.05를 통계학적으로 유의성이 있는 것으 로 판단하였다.
결 과
1. SCC25와 WI-38에서의 인테그린 α2와 EGFR 단백질 발현량 비교
인테그린 α2와 EGFR 단백질이 SCC25 세포에 대한 선택적
표적인자로서의 가능성을 알아보기 위해, 사람설암의 편평암종세 포인 SCC25 세포와 정상 폐섬유모세포인 WI-38 세포에서 인테 그린 α2와 EGFR의 발현량을 western blot 분석법으로 비교해 보았다. 인테그린 α2와 EGFR은 SCC25 세포에서 정상 세포인 WI-38 세포보다 훨씬 더 높은 발현량을 보였다(Fig. 1).
2. 인테그린 α2와 EGFR 차단항체에 의한 암세포의 선택적 세포사멸 유도
SCC25 세포와 WI-38 세포에서 인테그린 α2와 EGFR 차단항 체가 생존율에 미치는 영향을 보기 위해서 WST-1 분석을 했다.
먼저 SCC25 세포에 인테그린 α2와 EGFR 차단항체를 처리한 후 24시간이 지났을 때, 각각 94%와 96%로 감소됨을 보였으며 48시간에는 89%와 86%, 72시간에는 45%, 48%로 감소됨을 보였 다. 반면, 정상 세포인 WI-38 세포에서는 각각 24시간에는 137%, 134%, 48시간에는 144%, 141%, 72시간에는 89%, 88%의 증감 률을 나타내어, 이는 정상세포군주 배양에서 3일 이후는 세포밀도
Fig. 3. (A) SCC25 cells stained by Hemacolor. (B) After treatment of anti-integrin α2 antibody, cells were stained by Hemacolor. (C) After treatment of anti EGFR antibody, cells were stained by Hemacolor (A∼C: ×400). SCC, squamous cell carcinoma; EGFR, epidermal growth factor receptor.
Fig. 4. Translocation of AIF and cytochrome c. After 72 hours incubation in the presence of anti-integrin α2 and anti-EGFR antibody, subcellular location of AIF and cytochrome c were visualized by immunocytochemistry under confocal laser scanning microscope.
DAPI, diamidino phenylindole; AIF, apoptosis inducing factor; EGFR, epidermal growth factor receptor.
가 웰 플레이트상 100% confluence를 이루어 세포활성도가 떨어 지는 일반적인 정상세포배양 양상과 비슷한 결과를 보였다. 이 결과로 볼 때 인테그린 α2와 EGFR 차단항체가 폐섬유모세포보 다 사람설암의 편평암종세포의 생존율을 급격히 감소시키고, 정 상세포의 생존율에는 별다른 영향을 주지 않는다는 사실을 알 수 있었다(Fig. 2).
3. 인테그린 α2와 EGFR 차단항체 처리 후, SCC25 세포의 형태학적 변화
인테그린 α2와 EGFR 차단항체 처리 후, SCC25 세포의 형태학 적 변화를 관찰한 결과 SCC25 세포의 대조군에서는 핵이 둥글고
세포막의 모양이 방추형으로 정상적인 세포모양인 반면, 인테그 린 α2와 EGFR 차단항체를 72시간 처리했을 때는 세포의 핵이 응축되고 세포막이 터져 있는 것을 확인했다. 암세포의 형태학적 인 변화를 관찰했을 때, 세포사멸의 경로로 진행됐다는 사실을 알 수 있었다(Fig. 3).
4. AIF와 cytochrome c의 위치변화
면역형광염색 후, 공초점현미경으로 세포자멸사 과정이 진행 되고 있는 사립체(mitochondria)와 관련된 AIF와 cytochrome c의 위치변화를 관찰했다. 대조군에서는 AIF와 cytochrome c의 위치가 사립체 내에 위치하고 있는 반면, 인테그린 α2와 EGFR
Fig. 5. The activation of apoptosis related proteins. The treatment of anti-integrin α2 and anti-EGFR antibodies activates the apoptosis related proteins in SCC25 cells. The SCC25 cells were incubated in the presence of anti-integrin α2 or anti-EGFR antibody for the indicated times. The total protein lysates were used for SDS-PAGE followed by western blot using antibodies against lamin A/C, bax, bcl-xL, caspase-3, caspase-7, caspase-9, PARP and DFF45 antibodies. GAPDH was used as the loading control. EGFR, epidermal growth factor receptor; SCC, squamous cell carcinoma.
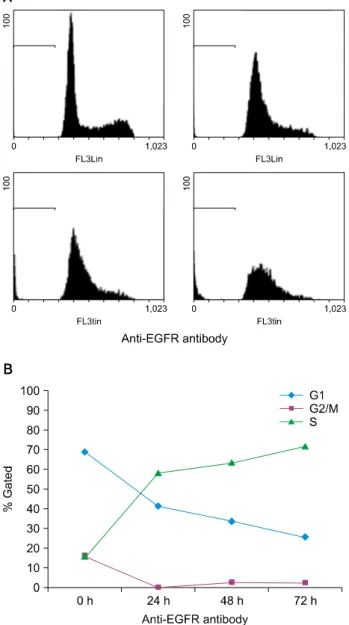
Fig. 6. The cell cycle arrest of SCC25 cells after treatment of anti-integrin α2. SCC25 cells treated with anti-integrin α2 blocking antibody for 72 hours show the induction of apoptosis and cell cycle arrest. At each time point, the cells are subjected to FACS analysis using PI solution as indicated (A). Cell cycle arrest was evaluated in terms of the proportion of cells in the S phase (B).
SCC: squamous cell carcinoma, PI: propidium iodide.
차단항체가 처리된 군에서는 AIF가 사립체에서 핵으로, cyto- chrome c가 사립체에서 세포질로 이동했다는 것을 관찰할 수 있었다(Fig. 4).
5. 세포자멸사와 관련된 단백질들의 활성화
Western blot 분석으로 세포자멸사 신호전달에서 대표적인 Bcl-2 (B-cell lymphoma 2) family에 대한 효과를 관찰했다.
실험 결과 인테그린 α2와 EGFR 차단항체를 처리를 했을 때, 시간이 지남에 따라 bax는 증가했고, bcl-xL 또한 증가했다. 이는 bax를 통해서 세포자멸사는 일어나지만 bcl-xL과는 독립적으로 세포자멸사 유도된다는 것을 알 수 있었다. 전형적인 세포자멸사 를 유도하는 조절인자인 caspase-3, caspase-7과 caspase-9도 마찬가지로 각각 24시간에서부터 각각의 쪼개진 형태로 나타나면 서 시간이 지남에 따라 활성화되고 있음을 알 수 있다. 또한 caspase-3의 활성으로 인해, 세포의 DNA를 회복시키는 기능을
수행하는 PARP가 각각 48시간에서부터 쪼개진 형태로 나타나며, DNase인 DFF40을 억제하고 있는 DFF45 (ICAD)가 시간이 지 남에 따라 파괴됨으로써 세포자멸사가 촉진됨을 알 수 있었다.
Caspase-6와 관련되어 세포핵의 골격을 담당하고 있는 lamin A/C의 발현이 감소되었다(Fig. 5).
6. 유세포분석법에 의한 세포주기와 sub-G1 지수 확인 세포자멸사와 관련된 sub-G1과 세포주기정지 변화를 알아보
Fig. 8. The deactivation of cell cycle related proteins. Anti-integrin α2 and anti-EGFR blocking antibodies effectively decrease the protein levels related cell cycle arrest in SCC25 cells. After incubating the SCC25 cells with anti-integrin α2 or anti-EGFR antibodies for the indicated times, the cells were subjected to SDS-PAGE coupled with western blot analysis. EGFR, epidermal growth factor receptor; CDK, cyclin dependent kinase; SCC, squamous cell carcinoma.
Fig. 7. The cell cycle arrest of SCC25 cells after treatment of anti-EGFR. SCC25 cells treated with anti-EGFR blocking antibody for 72 hours show the induction of apoptosis and cell cycle arrest.
At each time point, the cells are subjected to FACS analysis using PI solution as indicated (A). Cell cycle arrest was evaluated in terms of the proportion of cells in the S phase (B). SCC, squamous cell carcinoma; EGFR, epidermal growth factor receptor; PI, propidium iodide.
기 위해 유세포분석을 했다. 유세포분석은 DNA에 특이적으로 결합해 형광을 나타나내는 물질인 PI로 염색한 후 유세포분석기를 이용해 세포 내 DNA 함량을 측정하여 자멸사세포와 세포주기정 지를 백분율로 나타내는 것이다. 인테그린 α2와 EGFR 차단항체 처리 시, 대조군에 비해 세포자멸사의 지표인 sub-G1의 증가를 확인할 수 있었다(인테그린 α2 차단항체: 0 h 1.8%, 24 h 1.9%, 48 h 6.2%, 72 h 18.6%; EGFR 차단항체: 0 h 1.6%, 24 h 2.6%, 48 h 7.5%, 72 h 18.7%). 또한 인테그린 α2 차단항체 처리한 후 S기가 0시간에서 21.2%, 24시간에서 64.2%, 48시간
에서 77.6%, 72시간에서 84.9%로 증가되고 EGFR 차단항체 처 리 시 S가 0시간에서 15%, 24시간에서 58.7%, 48시간에서 63.3%, 72시간에서 72.3%으로 증가되었다. 이는 세포주기가 S기에서 증가된 것으로 보아 S기에서 세포주기정지가 진행되었음 을 알 수 있었다(Fig. 6, 7).
7. 세포주기에 관련된 단백질(cyclin D1, cyclin A, cyclin E, CDK 2, CDK 4, p27)량 변화
세포주기에 관련된 단백질의 변화를 알아보기 위해 western blot 분석을 시행했다. 인테그린 α2와 EGFR 차단항체를 처리했 을 때, 세포주기 G1기에 관련된 cyclin D1, CDK 4가 감소되었으 며, S기와 G2/M기가 관련된 cyclin A, cyclin E, CDK 2도 시간이 지남에 따라 발현이 감소된 것을 확인할 수 있었다. 하지만 CDK 저해단백질 합성을 유도를 하고, CDK 활성을 억제하는 것으로 알려져 있는 p27은 24시간부터 계속 감소된 것을 관찰할 수 있었 다(Fig. 8).
8. 섬유형 액틴의 붕괴
섬유형 액틴의 배열에 대해 관찰하기 위해 면역형광염색 검사 를 시행했다. 인테그린 α2와 EGFR 차단항체를 처리했을 때, 처리하지 않은 대조군에 비해 focal adhesion이 파괴되고, 세포 골격인 섬유형 액틴들이 붕괴되어 세포가 심하게 수축된 것이
Fig. 9. Destruction of actin filaments in SCC25 cells treated by anti-integrin α2 and anti-EGFR antibodies. Anti-integrin α2 and anti-EGFR blocking antibodies effectively induce the disruption of F-actin. F-actin staining of SCC25 cells treated with anti-integrin α2 or anti-EGFR for 72 hours. DAPI, diamidino phenylindole; EGFR, epidermal growth factor receptor; SCC, squamous cell carcinoma.
관찰되었다(Fig. 9).
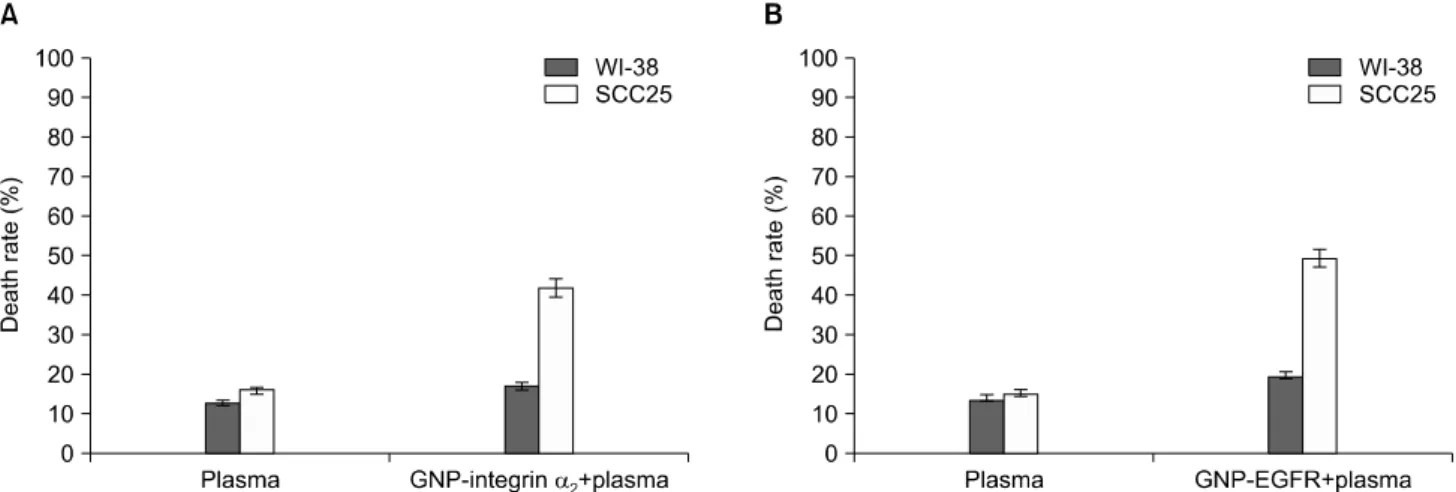
9. 저온 플라즈마와 중합체를 처리한 후의 세포사멸률 측정
좀 더 효과적인 암세포 사멸율 유도를 위하여 인테그린 α2와 EGFR에 대한 차단항체를 GNP에 결합시키고 저온 플라즈마를 조사하였다. 플라즈마를 30초간 처리 시 SCC25 세포에서 플라즈 마만 처리한 세포사멸률은 15.8%였고, 그에 반해, GNP-인테그 린 α2, GNP-EGFR 차단항체를 적용하고 플라즈마를 처리한 세
포사멸률은 각각 41.7%, 49.5%였다. 정상세포인 WI-38 세포에 서 플라즈마만 처리한 세포사멸률은 15.3%였고 GNP-인테그린 α2, GNP-EGFR를 적용하고 플라즈마를 처리한 세포사멸률은 각각 17.1%, 19.8%였다. 이는 중합체와 플라즈마 처리 시 정상세 포인 WI-38 세포보다 SCC25 세포에서 효과가 뛰어나며 인테그 린 α2와 EGFR 차단항체를 단독처리한 것보다 효과적인 세포사 멸이 일어난 것을 알 수 있었다(Fig. 10).
Fig. 10. Effects of plasma and GNP-integrin α2 (A) or GNP-EGFR (B) on the death rate of SCC25 cells and WI-38 cells. After plasma treatment, the cell death rate was measured using a trypan blue. GNP, gold nanoparticles; EGFR, epidermal growth factor receptor;
SCC, squamous cell carcinoma.
고 찰
기존의 항암제는 세포 독성 약물로서 세포 내에 일반적으로 존재하는 DNA나 미세관(microtuble)을 표적으로 하기 때문에 암세포에 대한 치료 효과를 나타내지만 정상 세포에도 악영향을 끼치는 부작용을 초래한다[20]. 최근 들어 암의 분자생물학적 특 징이 많이 규명되면서 특정 암세포의 표적인자만을 공격하는 표적 치료가 주목을 받고 있다.
본 연구에서는 구강편평상피암세포에서 많이 발현되는 표적인 자 단백질을 찾아 암세포의 선택적 제거를 유도하였다. 이를 위해 먼저 사람설암의 편평상피암세포인 SCC25 세포에서 발현된 인테 그린 α2와 EGFR의 양을 정상 폐섬유모세포인 WI-38 세포에서 발현된 양과 western blot 분석법으로 비교하였다. 인테그린 α2
와 EGFR은 암세포인 SCC25 세포에서 정상세포인 WI-38 세포보 다 훨씬 더 많이 발현되는 것을 관찰할 수 있었다. 이 결과를 토대로 SCC25 세포와 WI-38 세포에서 인테그린 α2와 EGFR 차단항체를 처리한 후 생존율을 비교하였을 때 시간이 지남에 따라 SCC25 세포에서 생존율이 확연히 감소하였으며 페섬유모세 포에서는 별다른 영향을 주지 않았다. 인테그린 α2와 EGFR 차단 항체를 처리한 이 두 군의 세포생존율 사이에는 유의한 차이가 나타나지 않았다.
인테그린은 세포의 신호전달 수용체로 세포와 세포외 기질을 연결하는 구조로 세포 내 골격에 연결해주는 focal adhesion을 형성한다[21]. 인테그린을 통한 신호전달은 focal adhesion을 형성하는 FAK, paxillin, talin 등의 다양한 단백질을 통해 세포 내로 이루어지며 세포의 결합을 통제하고 부착 후 세포 내 반응을 조절하는 역할을 한다[22,23]. 이러한 focal adhesion이 파괴되 면, 세포외 기질로부터 세포가 떨어져 세포는 사멸된다. 차단항체 결합 시 신호전달이 차단되면 그 하부 단위의 단백질의 인산화가
차단되어 focal adhesion이 파괴되고 세포사멸이 일어나는 것으 로 추정된다.
상피성장인자와 세포막에 존재하는 EGFR과의 결합은 수용체 의 세포 내 tyrosine kinase를 활성화시켜 DNA의 복제와 세포 분열을 유도하고 세포의 증식을 촉진시키는 데 중요한 역할을 한다[5]. 현재 EGFR을 차단하여 tyrosine kinase의 활성을 막는 항암치료제 연구가 다양하게 시도되고 있으며 임상에서 사용되고 있다[9,10,24,25]. 본 연구에서 EGFR과 차단항체의 결합시 ty- rosine kinase의 활성이 일어나지 않아 하부단위 단백질로의 신 호전달이 차단되어 세포사멸이 일어났으리라 추정된다.
인테그린 α2와 EGFR 차단항체 처리 후 SCC25 세포에서 생존 율 감소 현상이 암세포의 형태 변화에 영향을 미치는지를 알아본 결과, 핵이 응축되고 세포막이 터져 있는 것을 확인했고 세포사멸 의 경로로 진행됐다는 것을 알 수 있었다.
세포의 죽음에는 물리적, 화학적 자극에 의한 세포손상에 따른 세포괴사와 세포자멸사가 있다. 이 중 세포자멸사는 세포의 성장 과 조직의 항상성을 조절하는 데 있어서 매우 중요한 역할을 하며, 암의 치료에 중요한 방식으로 생각된다. 일반적으로 세포자 멸사는 death receptor를 통한 외재적 경로(extrinsic pathway) 와 caspases, bcl-2 family 등과 같은 다양한 유전자가 관여하는 내재적 경로(intrinsic pathway)를 통해서 유발된다[26,27]. 본 연구에서는 먼저 인테그린 α2와 EGFR 차단항체 처리 후 나타난 SCC25 세포의 사멸이 세포자멸사인지 여부를 확인하기 위해서 세포자멸사를 나타내는 내재적 경로 유전자 요소인 AIF와 cyto- chrome c의 위치변화를 관찰하였다[28,29]. 이들은 사립체 내에 존재하다 사립체의 외막 투과성이 증가되면 세포질로 방출된다.
AIF는 caspase와 독립적으로 세포핵 안으로 이동하여 염색질을 농축, 분절화시켜 세포자멸사를 유발한다. Cytochrome c는 세포 질로 방출되어 apoptotic protease activating factor-1 (Apaf-1)
에 결합하여 apoptosome을 형성하여 caspase-9를 활성화시켜 세포자멸사를 유발한다[30]. 대조군에서는 AIF와 cytochrome c의 위치가 사립체 내에 위치하고 있는 반면, 인테그린 α2와 EGFR 차단항체를 처리된 군에서는 AIF가 사립체에서 핵으로, cytochrome c가 사립체에서 세포질로 이동했다는 것을 관찰하 였으며 이는 내재적 경로에 의해 세포자멸사가 일어났다는 사실을 보여준다. 또한 내재적 경로에서 세포자멸사 신호전달에서 대표 적인 bcl-2 family에 대한 효과를 관찰하였다. bcl-2 family는 사립체 내막과 핵막, 소포체 등에 존재하는 세포자멸사의 억제와 촉진을 조절하는 유전자 군이다. bcl-2, bcl-xL은 cytochrome c의 유리를 막아 세포자멸사를 저해하고 bax, bad, bak는 cyto- chrome c 유리를 촉진하여 세포자멸사를 유도한다. 본 실험에서 는 bcl-2 family 중에서 가장 대표적인 세포사멸 억제신호인 bcl-xL와 촉진신호인 bax를 분석하였다. 인테그린 α2와 EGFR 차단항체를 처리 후 시간이 지남에 따라 bax는 증가했고, 이와 길항 작용을 하는 bcl-xL 또한 증가한 것으로 보아 bax를 통해서 세포자멸사가 일어나지만 bcl-xL과는 독립적으로 세포자멸사가 유도된다는 것을 알 수 있었다[31]. 전형적인 세포자멸사를 유도 하는 조절인자인 caspase는 핵과 사립체 외막에 proenzyme 형태로 존재하다 세포 내ㆍ외의 신호에 의해서 활성화되어 표적 단백질들의 분해를 유발한다. 많은 선행연구에서 caspase의 활성 화 자체가 세포자멸사 유발에 대한 증거가 될 수 있다고 보고되어 본 연구에서는 caspase-3, -7, -9의 발현 및 활성에 대해 조사하였 다. Caspase-3, -7, -9의 발현이 시간이 지남에 따라 활성화되고 있음을 알 수 있었다. 또한 caspase-3의 활성으로 인해, 세포의 DNA를 회복시키는 기능을 수행하는 PARP와 DNase인 DFF40 을 억제하고 있는 DFF45 (ICAD)가 시간이 지남에 따라 파괴됨으 로써 세포자멸사가 촉진됨을 알 수 있었다[32,33]. Caspase-6와 관련되어 세포핵의 골격을 담당하는 lamin A/C가 감소된 것을 관찰할 수 있어 핵의 수축이 진행되어 세포자멸사가 진행됨을 알 수 있었다.
세포자멸사에 들어간 세포는 세포 내 탈수로 크기가 작아지면 서 염색질이 농축되고 세포 소기관들이 뭉쳐서 세포자멸사소체 (apototic body)가 형성되어 탐식된다[34]. 이때, 분절화가 일어 나는 DNA에 형광을 나타내는 물질인 PI로 염색한 후 세포자멸사 세포와 세포주기정지를 조사하였다. 세포가 증식하기 위해서 G1
기를 거쳐 DNA 복제(S기), 그리고 G2기를 거쳐 세포분열(M기) 을 하는 일련의 단계를 거치는데 세포손상이 발생하게 되면 이를 회복하기 위한 방어체계의 하나로 세포주기정지를 한다. 세포자 멸사에서는 DNA가 분절되어 PI 발현이 매우 적은 sub-G1 세포가 증가되게 된다. 인테그린 α2와 EGFR 차단항체를 처리한 SCC25 세포의 유세포분석기를 이용한 분석에서 sub-G1 세포가 시간이 지남에 따라 점점 증가하는 것을 관찰할 수 있었으며, 이로, 세포 주기의 S기에서 세포주기정지가 진행되었음을 알 수 있었다. 이것
으로 세포주기가 정지되고 지속적인 세포자멸사가 일어났음을 알 수 있었다.
세포주기에 관련된 단백질의 변화를 분석했을 때 인테그린 α2와 EGFR 차단항체를 처리한 SCC25 세포에서 세포주기 G1기 에 관련된 cyclin D1, CDK 4가 감소되었으며, S기와 G2/M기가 관련된 cyclin A, cyclin E, CDK 2도 시간이 지남에 따라 발현이 감소되었다[35-37]. CDK 저해 단백질 합성을 유도를 하고 CDK 활성을 억제하는 것으로 알려져 있는 p27이 감소된 것을 확인할 수 있었다. p27은 암억제유전자인 p53에 의해서 활성화되어 G1
기를 유도하는 cyclin D1-CDK 복합체에 결합해 CDK 4/6의 kinase 활성을 저해하여 망막아종단백질(retinoblastoma pro- tein [RB protein])의 인산화를 막는다. 이에 p27과는 독립적으로 일어나는 세포주기정지를 양상을 보이며 전반적인 세포주기 정지 에 의해서 p27의 감소가 일어난 것으로 볼 수 있다[38]. 유세포분 석기의 결과에서는 S기에서 세포주기가 정지하는 것으로 나왔지 만, 세포주기와 관련된 단백질들의 발현은 S기에만 국한되지 않 고, 대부분의 단백질이 감소하는 것으로 볼 때, 세포주기가 전반적 으로 정지되었음을 알 수 있었다. 그리고 유도된 세포자멸사 현상 또한 세포주기정지와는 관련이 없을 것으로 여겨진다.
세포자멸사가 일어나면 세포의 형태가 변화하게 된다. 섬유형 액틴은 세포의 기계적인 지지를 제공하고 세포의 정교한 모양을 유지시키는 역할을 한다. 세포자멸사가 일어나면 세포가 수축하 면서 섬유형 액틴의 붕괴가 일어나며 세포 부착의 파괴가 일어나 게 된다. 인테그린 α2와 EGFR 차단항체를 처리했을 때, 처리하 지 않은 대조군에 비해 focal adhesion이 파괴되고, 세포골격인 섬유형액틴들이 붕괴되어 세포가 심하게 수축된 것이 관찰된다.
위의 결과를 종합해서 살펴보면 구강편평상피암 세포에서 인테 그린 α2와 EGFR에 대한 차단항체를 사용하였을 때, 세포의 생존 율 감소가 나타났으며 이 생존율 감소가 세포자멸사 양상으로 나타났음을 확인할 수 있었다.
이에 SCC25 세포의 표적인자로 확인된 인테그린 α2와 EGFR 을 이용하여 좀 더 효과적인 암세포 사멸을 유도하기 위해서 인테그린 α2와 EGFR 차단항체를 인체에 독성이 없는 GNP에 결합시켜 각각의 중합체(GNP-Integrin α2, GNP-EGFR)를 만들 고 난 후, 저온 플라즈마를 조사하였다. 본 실험에서 구강편평상피 암세포에서 플라즈마를 단독 처리한 경우보다 GNP-Integrin α2, GNP-EGFR 중합체를 만들어 적용한 후 플라즈마를 처리한 경우 약 3배 증가된 세포사멸률을 나타내었으며 GNP-EGFR 중합체를 처리한 경우에서 GNP-Integrin α2 중합체를 처리한 경우보다 세포사멸율이 약간 높게 나타났다. 정상 섬유모세포에서는 플라 즈마를 처리한 군과 GNP-Integrin α2, GNP-EGFR 중합체를 적용하고 플라즈마를 처리한 군과의 차이가 나타나지 않았다.
이는 중합체와 플라즈마 처리 시 구강편평상피암세포에서 정상세 포보다 훨씬 효과적인 암세포 제거가 일어난다는 것을 알 수
있었다. GNP-Integrin α2와 GNP-EGFR 중합체를 처리한 경우 에 구조적으로 암세포와 잘 결합하여 항체 고유의 암세포 사멸효 과에 더하여 플라즈마에 의해 자극된 GNP가 표적단백질의 활성 을 억제함으로써 구강편평상피암 세포에 대한 선택적 사멸이 더 효력을 발휘한 것으로 추정된다[39,40].
이상의 결과를 종합해 보면 인테그린 α2와 EGFR을 구강편평 상피암 세포의 표적인자로 사용할 수 있음을 확인하였으며 이에 대한 항체 처리 시 그 사멸 양상이 세포자멸사로 일어남을 알 수 있었다. 또한 항체와 GNP의 중합체를 만들어 플라즈마 처리 시 정상세포에 비해 구강편평상피암 세포의 사멸이 훨씬 효과적으 로 일어나 구강편평상피암의 치료 시 새로운 선택적 방법이 될 수 있음을 제시한다.
결 론
본 연구는 설암의 구강편평상피암세포인 SCC25에서 인테그린 α2와 EGFR의 발현 양을 western blot 분석법을 통해 정상 폐섬 유세포인 WI-38 세포와 비교 관찰하였다. 또한 인테그린 α2와 EGFR에 대한 항체를 사용하여 WST-1 법으로 세포생존율을 측정 하였고, Hemacolor 염색법, Immunoflorescence 염색법, FACS 분석법, 그리고 western blot 분석법을 실시하여 세포주기 정지 및 세포자멸사를 관찰하여 인테그린 α2와 EGFR의 항체를 통한 암세포의 사멸을 확인하였다.
그 후 인테그린 α2와 EGFR의 항체에 GNP를 적용한 중합체를 만들어 플라즈마를 처리하여 암세포의 선택적 제거가 유도되는 추가적인 결과를 얻었다.
1. 정상 세포인 WI-38 세포보다 암세포인 SCC25 세포에서 인테그린 α2와 EGFR이 훨씬 많이 발현이 되었다.
2. 인테그린 α2와 EGFR의 차단항체를 처리한 경우에 WI-38 세포에서는 생존율에 영향을 받지 않았으나, SCC25 세포는 시간 이 지남에 따라 생존율이 감소하였다.
3. 인테그린 α2와 EGFR의 차단항체를 처리한 경우 SCC25 세포에서는 핵의 모양과 세포막의 변화양상을 관찰하여 세포사멸 을 확인하였으며 AIF가 사립체에서 핵으로, cytochrome c가 사립체에서 세포질로 이동을 확인하였다.
4. 세포자멸사와 관련된 단백질과 관련해서는 bax의 증가, 전형적인 세포자멸사를 유도하는 조절인자인 caspase-3, cas- pase-7과 caspase-9의 활성화, PARP와 DNase인 DFF40을 억 제하고 있는 DFF45 (ICAD)가 감소하여 세포자멸사를 확인할 수 있었다. 또한 lamin A/C는 감소를 확인하였으며, bcl-xL는 증가를 확인하였다.
5. 유세포 분석에서는 세포자멸사의 지수를 나타내는 sub G1
의 증가와 S기의 증가를 확인하였다. 인테그린 α2와 EGFR의 차단항체 처리 후에 p27의 감소와 cyclin D1, cyclin A, cyclin
E, CDK 2 그리고 CDK 4 발현이 감소를 관찰하여 세포주기정지 를 유도하는 것을 확인하였다. 아울러 섬유형 액틴의 붕괴를 관찰 하여 세포가 수축된 것을 알 수 있었다.
6. 인테그린 α2와 EGFR에 대한 항체를 GNP에 결합시키고 저온 플라즈마를 조사를 한 결과 세포사멸률이 WI-38 세포보다 SCC25 세포에서 3배 정도 효과가 뛰어남을 관찰할 수 있었다.
위의 결과를 종합해 보면, SCC25 세포에서 인테그린 α2와 EGFR이 과잉발현되며 이에 대한 차단항체 처리 시 이들 단백질의 기능을 강력하게 저해하여, 세포자멸사 양상으로 세포생존율의 감소가 일어나는 것이 확인되었다. 아울러 이들 항체와 GNP 중합체를 만들어 플라즈마 조사한 경우 보다 효과적인 구강편편상 피암세포의 사멸이 일어나는 것으로 조사되었다. 향후 구강편평 상피암 치료 시 인테그린 α2와 EGFR을 표적인자로 사용하여 GNP와 플라즈마를 이용한 접근법이 새로운 선택적 방법이 될 수 있음을 제시한다.
References
1. Hughes AL. Evolution of the integrin alpha and beta protein families. J Mol Evol 2001;52:63-72.
2. Guo W, Giancotti FG. Integrin signalling during tumour progression. Nat Rev Mol Cell Biol 2004;5:816-26.
3. Guo L, Zhang F, Cai Y, Liu T. Expression profiling of integrins in lung cancer cells. Pathol Res Pract 2009;205:847-53.
4. Bargmann CI, Hung MC, Weinberg RA. The neu oncogene encodes an epidermal growth factor receptor-related protein.
Nature 1986;319:226-30.
5. Herbst RS. Review of epidermal growth factor receptor biology. Int J Radiat Oncol Biol Phys 2004;59(2 Suppl):21-6.
6. Ishitoya J, Toriyama M, Oguchi N, et al. Gene amplification and overexpression of EGF receptor in squamous cell carci- nomas of the head and neck. Br J Cancer 1989;59:559-62.
7. Neve RM, Lane HA, Hynes NE. The role of overexpressed HER2 in transformation. Ann Oncol 2001;12 Suppl 1:S9-13.
8. Ullrich A, Schlessinger J. Signal transduction by receptors with tyrosine kinase activity. Cell 1990;61:203-12.
9. Young NR, Liu J, Pierce C, et al. Molecular phenotype predicts sensitivity of squamous cell carcinoma of the head and neck to epidermal growth factor receptor inhibition. Mol Oncol 2012.
pii: S1574-7891(12)00121-4.
10. Markovic A, Chung CH. Current role of EGF receptor mono- clonal antibodies and tyrosine kinase inhibitors in the man- agement of head and neck squamous cell carcinoma. Expert Rev Anticancer Ther 2012;12:1149-59.
11. Kim KW, Kim MJ. Expression of the epidermal growth fac- tor receptor and cell cycle analysis in the head and neck squamous cell carcinomas. J Korean Assoc Oral Maxillofac Surg 2000;26:154-63.
12. Moon BC, Han SJ, Jeong D, Kim KW. Epidermal growth factor receptor overexpression and K-ras mutation detection in the oral squamous cell carcinoma. J Korean Assoc Oral Maxillofac Surg 2011;37:396-402.
13. Fridman G, Fridman G, Gutsol A, Shekhter AB, Vasilets VN,
Fridman A. Applied plasma medicine. Plasma Process Polym 2008;5:503-33.
14. Fridman G, Shereshevsky A, Jost MM, et al. Floating elctrode dielectric barrier discharge plasma in air promoting apoptotic behavior in melanoma skin cancer cell lines. Plasma Chem Plasma Process 2007;27:163-76.
15. Kim CH, Kwon S, Bahn JH, et al. Effects of atmospheric nonthermal plasma on invasion of colorectal cancer cells.
Appl Phys Lett 2010;96:243701.
16. Kim D, Gweon B, Kim DB, Choe W, Shin JH. A feasibility study for the cancer therapy using cold plasma. ICBME Proc 2008;23:355-7.
17. Kim GC, Lee HJ, Shon H. The effects of a micro plasma on melanoma (G361) cancer cells. J Korean Phys Soc 2009;54:628-32.
18. Connor EE, Mwamuka J, Gole A, Murphy CJ, Wyatt MD.
Gold nanoparticles are taken up by human cells but do not cause acute cytotoxicity. Small 2005;1:325-7.
19. Thomas M, Klibanov AM. Conjugation to gold nanoparticles enhances polyethylenimine's transfer of plasmid DNA into mammalian cells. Proc Natl Acad Sci U S A 2003;100:9138-43.
20. Levy JM, Thorburn A. Targeting autophagy during cancer therapy to improve clinical outcomes. Pharmacol Ther 2011;
131:130-41.
21. Chen CS, Alonso JL, Ostuni E, Whitesides GM, Ingber DE.
Cell shape provides global control of focal adhesion assembly. Biochem Biophys Res Commun 2003;307:355-61.
22. Riveline D, Zamir E, Balaban NQ, et al. Focal contacts as mechanosensors: externally applied local mechanical force induces growth of focal contacts by an mDia1-dependent and ROCK-independent mechanism. J Cell Biol 2001;153:
1175-86.
23. Zamir E, Geiger B. Molecular complexity and dynamics of cell-matrix adhesions. J Cell Sci 2001;114:3583-90.
24. Ennis BW, Lippman ME, Dickson RB. The EGF receptor sys- tem as a target for antitumor therapy. Cancer Invest 1991;9:
553-62.
25. Mendelsohn J, Baselga J. Epidermal growth factor receptor targeting in cancer. Semin Oncol 2006;33:369-85.
26. Ghavami S, Hashemi M, Ande SR, et al. Apoptosis and cancer:
mutations within caspase genes. J Med Genet 2009;46:497-510.
27. Dejean LM, Martinez-Caballero S, Manon S, Kinnally KW.
Regulation of the mitochondrial apoptosis-induced channel, MAC, by BCL-2 family proteins. Biochim Biophys Acta 2006;
1762:191-201.
28. Susin SA, Lorenzo HK, Zamzami N, et al. Molecular charac- terization of mitochondrial apoptosis-inducing factor. Nature 1999;397:441-6.
29. Dejean LM, Martinez-Caballero S, Kinnally KW. Is MAC the knife that cuts cytochrome c from mitochondria during apoptosis? Cell Death Differ 2006;13:1387-95.
30. Alnemri ES, Livingston DJ, Nicholson DW, et al. Human ICE/CED-3 protease nomenclature. Cell 1996;87:171.
31. Murphy KM, Ranganathan V, Farnsworth ML, Kavallaris M, Lock RB. Bcl-2 inhibits Bax translocation from cytosol to mi- tochondria during drug-induced apoptosis of human tumor cells. Cell Death Differ 2000;7:102-11.
32. Yu SW, Andrabi SA, Wang H, et al. Apoptosis-inducing fac- tor mediates poly(ADP-ribose) (PAR) polymer-induced cell death. Proc Natl Acad Sci U S A 2006;103:18314-9.
33. Widłak P. The DFF40/CAD endonuclease and its role in apoptosis. Acta Biochim Pol 2000;47:1037-44.
34. Karam JA. Apoptosis in carcinogenesis and chemotherapy.
Netherlands: Springer; 2009.
35. Morgan DO. Principles of CDK regulation. Nature 1995;374:
131-4.
36. Sherr CJ. G1 phase progression: cycling on cue. Cell 1994;
79:551-5.
37. Pines J. Cyclins: wheels within wheels. Cell Growth Differ 1991;2:305-10.
38. Si X, Liu Z. Expression and significance of cell cycle-related proteins Cyclin DI, CDK4, p27, E2F-I and Ets-1 in chon- drosarcoma of the jaws. Oral Oncol 2001;37:431-6.
39. Kim GC, Kim GJ, Park SR, et al. Air plasma coupled with antibody-conjugated nanoparticles: a new weapon against cancer. J Phys D Appl Phys 2009;42:032005.
40. Choi BB, Choi YS, Lee HJ, Lee JK, Kim UK, Kim GC.
Nonthermal plasma-mediated cacer cell death: targeted can- cer treatment. J Therm Sci Technol 2012;7:399-404.