대한소화기학회지 2000;36:493 - 503
3)
접수: 2000년 5월 16일, 승인: 2000년 10월 6일 연락처: 박영숙, 139-230, 서울시 노원구 하계동 280-7
노원을지병원
Tel: (02) 970-8207, Fax: (02) 970-8429 E-mail: [email protected]
※ 본 논문은 1998년도 을지대학교 의과대학 교수연구비 의 보조로 이루어졌음.
서 론
궤양성 대장염과 크론병으로 대표되는 염증성 장 질환의 병태생리는 장내세균, 세균산물, 독소, 약제,
염증성 장질환 환자의 점막 고유층 T 림프구에서 C D4 C D11b 표면항원 발현 양상
을지대학교 의과대학 내과학교실, Cedars-Sinai Inflammatory Bowel Disease Center*
박영숙・S tephan R.Targan*
E x p r e s s i o n o f CD 4 CD 1 1 b An t i g e n o n La m i n a P r o p ri a T c e ll s i n P a t i e n t s w i t h In fla m m a t o r y B o w e l D i s e a s e
Yo u n g S oo k P a r k , M.D. a n d S t e p h an R. Ta r g an , M.D.*
Department of Internal Medicine, Eulji University School of Medicine, Seoul, Korea;
Inflammatory Bowel Disease Center*, Cedars-Sinai Medical Center, Los Angeles, U.S.A.
Background/Aims: CD11b/CD18, a member of β2 integrin family, is mainly expressed on
monocytes, neutrophils, NK cells and some T cells. We investigated the expression of CD4+CD11b+in lamina propria (LP) T cell in patients with inflammatory bowel disease (IBD) and tried to document their function by detection of intracellular cytokine expression. Methods: Intestinal specimen was obtained from 16 patients with IBD and 12 patients with colon cancer. After isolation of LP T cells, immunofluorescent staining was performed using monoclonal antibodies and flow cytometry. Results: The expression of CD4+CD11b+ on LP T cells was significantly higher in IBD patients than in control (UC:4.42±1.2%, CD:3.56±0.7% control:1.57±0.8%, p=0.03). On 7 to 10 day culture, the CD4+CD11b+ expression was increased up to 10% in all patients. Intracellular cytokine expression of interferon-γ, tumor necrosis factor-α showed no significant difference between CD11b+ and CD11b- T cells. Interleukin (IL)-10 was expressed highly on some of CD11b- T cells and showed only borderline brightness on 10% of CD11b+ T cells. Conclusions: In patients with IBD, the expression rates of CD4+CD11b+ on LP T cells were higher than those in normal controls. However, intracellular cytokine expression was not specific between CD11b+ and CD11b- T cells. (Kor J Gastroenterol 2000;36:493 - 503)
Key Words: Lamina propria T cells, CD11b/CD18, INF-γ, TNF-α, IL-10
494 대한소화기학회지 : 제 36 권 제 4 호 2000
음식 내 항원 등에 의한 면역반응이 개시, 지속 및 증폭되어 비특이적 염증반응과 조직 손상을 일으키 리라 생각되고 있다.1 면역반응의 지속 및 증폭은 면역조절 기능의 결함이 그 원인으로 크론병에서는 Th1림프구의 활성화반응이 궤양성 대장염에서는 Th2림프구의 활성화가 우세함이 보고되었다.2 면역 조절 기능의 장애는 전염증 분자(pro-inflammatory molecule)와 항염증 분자(anti-inflammatory mole- cule) 사이의 불균형을 의미하며, interleukin (IL)-10 과 같은 항염증 분자에 의한 적절한 염증반응 감쇄 가 병적 면역반응을 억제할 수 있음이 밝혀지면서,3 IL-10을 다량 분비하는 소위 조절 T림프구(regula- tory T cell)라 불리우는 장내 세포군을 찾고자 하는 연구가 진행되고 있다. 그러므로 특정 표면항원을 발현하는 세포군과 이들 세포가 분비하는 사이토카 인을 규명하여 특정 세포군의 장내 역할을 밝혀보 고자 한다.
CD11b/CD18은 일명 Mac-1으로 불리우며, β2백 혈구 접합분자군(leukocyte integrin family) 가운데 하나로, 단핵구와 중성구를 내피세포에 부착시키는 일에 관여하는 세포막단백이다.4 CD11b/CD18은 주 로 단핵구, 중성구 및 자연살해세포(natural killer cell)에서 발현되고,5 CD8+ T림프구의 50%에서, CD4+ T림프구에서는 5% 미만의 제한된 발현을 보 인다.3 사람 말초혈액의 CD4+ T림프구에서 CD28- 림프군이 CD11b를 발현한다고 보고되었고,6 CD4+
CD28-CD11b+ T림프군에서는, CD28+ T림프군과 는 달리 CD3 단독 자극에 의해서도 활성화되며 CD3에 의해 2차 자극을 해도 anergy를 보이지 않 고,7 아포토시스(apoptosis)에도 저항을 보이는 림프 군으로 보고하였다.8 염증성 장질환 환자의 혈액 내 단핵구 및 과립세포에서 CD11b/CD18의 발현이 증가 되어 있다는 보고와,9 2,4,6-trinitrobenzene sulfonic acid (TNBS)에 의해 유도된 장염에 CD11b/CD18에 대한 항체를 투여하여 효과적 관해가 이루어졌음이 보고되었다.10 그러나 염증성 장질환 환자의 대장점막 고유층의 T림프구에서 CD11b 표면항원 발현 및 이 들 세포의 역할에 대한 연구는 아직 보고된 바 없다.
저자 등은 염증성 장질환 환자의 점막 고유층 T 림프구에서 CD11b의 발현 양상 및 이들 세포의 세
포 내 사이토카인 염색을 통하여 CD11b+ 림프구의 염증성 장질환의 병태생리에 있어서의 역할을 규명 해 보고자 본 연구를 시행하였다.
연구 대상
대장 조직은 UCLA Cedars-Sinai Medical Center 에서 1998년 9월부터 1999년 8월까지 소장 및 대장 절제술을 받은 28예(남자; 22예, 여자; 6예)의 환자 로부터 얻었다. 이들은 대장암 12예와 크론병 9예 및 궤양성 대장염 환자 7예였다. 대장암 환자의 평 균 연령은 67.2세(49-82)이고, 염증성 장질환 환자의 평균 연령은 41.2세(27-56)였다. 본 실험은 Cedars- Sinai Medical Center산하 Institutional Review Board 로부터 승인을 받은 후 시행하였다.
연구 방법
1. 대장점막 고유층 단핵세포의 분리
정상 대조군으로 대장암 환자의 수술 조직 중 대 장암 이환 부위로부터 10 cm 이상 떨어진 정상 조 직을 취하였으며, 궤양성 대장염 환자에서는 이환된 조직을, 크론병에서는 이환된 조직과 이환되지 않은 조직을 각각 취하였다.
점막 고유층 단핵세포의 분리는 Bull과 Bookman 이 처음 기술하고,11 본 교실에서 수정하여 사용하고 있는 방법을 이용하였다.12 요약하면 대장 조직은 Hank' s balanced salt solution (HBSS)으로 세척한 다 음, 점막을 분리하였다. 분리한 점막은 칼슘과 마그 네슘이 없는 HBSS에 담그고, 1 mM EDTA, 50 μg/
ml gentamycin, 100 U/ml penicillin, 100 μg/ml streptomycin, and 50 μg/ml fungizone을 첨가한 후 상피세포가 제거되기까지 30분 간격으로 용액을 교 환하며 진탕 수조(100 rpm)에서 세척하였다. 부유액 을 제거하고 남은 점막 고유층은 1-2 mm의 작은 조 각으로 분쇄한 후 10% 우태아혈청, 0.5 mg/ml collagenase B (Boehringer Mannheim, Indianapolis, IN), 1 mg/ml hyaluronidase (Sigma, St. Louis, MO), 0.1 mg/ml DNase I (Sigma, St. Louis, MO), 50 μg/
ml gentamycin, 100 U/ml penicillin, 100 μg/ml
박영숙 외 1인. 염증성 장질환 환자의 점막 고유층 T림프구에서 CD4CD11b 표면항원 발현양상 495
streptomycin 및 50 μg/ml fungizone을 포함한 RPMI 1640 (Irvine Scientific, Santa Ana, CA) 용액에 담궈 진탕 수조에서 10분간 소화시켰다. 부유액을 수집하 여 11 μm nylon mesh (Spectrum Laboratory Pro- ducts, Houston, TX)로 거른 후, 500 Xg에서 5분간 원심분리하였다. 세포환(cell pellet)으로부터 남은 상피세포와 기타 대세포 등을 제거하기 위해 용액 15 mL에 다시 부유시킨 후 30 Xg에서 5분간 원심 분리를 하였다. 부유액은 제거하고, 림프구는 Ficoll- Hypaque에 의한 비중 차이로(gradients) 분리하였다.
림프구는 HBSS 용액으로 3차례 씻은 후 10% 우태 아혈청을 첨가한 RPMI 1640에 부유시켰다.
2. 세포 배양
세포는 10% 우태아혈청과 10 U/mL의 recom- binant human interleukin-2 (rIL-2; R&D Systems, Minneapolis, MN)를 첨가한 RPMI 1640에 부유시켜 37℃, 5% CO2 조건에서 배양하였다.
3. 단클론항체
anti-CD11b FITC, anti-CD28 Phycoerythrin (PE), anti-CD3 Tricolor (TC), anti-CD4 TC, anti-CD3 PE, anti-CD25 (IL-2 receptor) PE는 Caltag사(Burlin- game, CA)에서 구입하였고, Anti-IL10 PE, purified anti-IL-10, anti-CD11b PE, anti-CD18 PE, anti-IFN- γ PE 항체는 Becton Dickinson사(San Jose, CA)로 부터 구입하였다. anti-TNF-α PE 항체는 Olympus Immunochemicals (Lake Success, NY)에서 Anti- CD45RO 항체는 Dako corporation (Carpenteria, CA)에서 구입하였다. anti-T112와 anti-T113 (anti- CD2)는 Reinherz E.L (Laboratory of Immunobio- logy, Depatment of Pathology, Harvard medical school, Boston, MA)가 보내주었고, 9.3 (anti-CD28) 및 G19.4 (anti-CD3) 항체는 Ledbetter J.A (Onco- gen, Seattle, WA)가 친절하게 제공하였다.
4. 면역형광염색(Immunofluorescent s taining) 표면항원 염색은 분리한 점막 고유층 림프구 5×
105개를 2% 우태아혈청과 0.1% sodium azide를 첨 가한 용액(PBS-FCS) 100 μl에 부유시킨 후 형광이
부착된 단클론항체를 첨가하고 4℃에서 30분간 염 색하였다.
5. 세포 내 사이토카인 염색
염색하고자 하는 림프구를 골지체(Golgi' s body) 의 기능을 억제하는 Brefeldin A (Golgi plug, Sigma, 1 μg/mL) 혹은 Monensin (Golgi Stop, Pharmingen, 2 μM/mL)을 첨가한 배양용액에서 16내지 18시간 동안 배양시킨 후, 수집하여 phosphate buffered saline-fetal calf serum (PBS-FCS)용액에 세척하고, CD3, CD4, CD28, CD11b 등의 표면항원 염색을 시 행하였다. 표면항원 염색이 끝난 세포들은 PBS-FCS 용액에 세척 후 원심분리하여 세포환을 만들고, 100 μl의 고정완충액(Fix and Perm solution, Caltag, Reagent A)에 부유시켜 15분간 실온에서 고정시켰 다. 세포들을 다시 세척하고, 100 μl의 투과완충액 (Reagent B)에 부유시킨 후 0.5 μg의 형광이 부착된 INF-γ, TNF-α, IL-10의 단클론항체와 함께 4℃에 서 1시간 동안 배양하였다. 배양 후 PBS-FCS 용액 으로 2회 세척하고, 1% paraformaldehyde를 첨가한 PBS 용액에서 고정한 후, FACScan flow cytometer (Becton Dickinson)를 이용하여 림프구 아군과 세포 내 사이토카인을 분석하였다.
6. 통계 분석 방법
통계 처리는 chi-squre와 Fisher' s exact test로 처 리하였으며 p값이 0.05 이하일 때 통계적 유의성을 인정하였다.
결 과
1. 정상 대조군 및 염증성 장질환 환자군의 점막 고유층 T 림프구에서 CD4+ CD11b+
표면항원의 발현
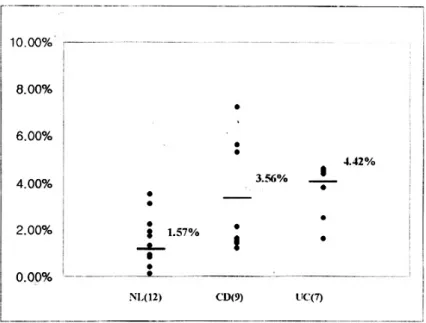
수술 조직에서 분리한 직후 점막 고유층 T세포의 CD4+CD11b+ 발현율은 정상 대조군(12예)에서 1.57±0.8%, 크론병 환자군(9예)에서 3.56±0.7%, 궤양성 대장염 환자군(7예)에서 4.42±1.2%로서 크 론병 및 궤양성 대장염 환자군이 정상 대조군에 비 해 유의한 발현율 증가를 보였다(Fig. 1, p=0.03). 대
496 The Korean Journal of Gastroenterology : Vol. 36, No. 4, 2000
장 및 소장점막 고유층 T세포에 있어 CD28 발현과 CD11b 발현은 상호 연관성을 보이지 않아, CD4+
CD28-는 2.5% CD4+CD28-CD11b+ 0.4%로 CD11b 발현군과 CD28의 비발현군이 동일하지 않음을 시
사하였다(Fig. 2).
2. C D11b 표면항원 발현 T 림프구의 특징 CD3+CD4+CD11b+ T세포의 표면항원 특징은
Fig. 2. Co-expression of CD11b and CD28. The co-expression rate of CD11b+ and CD28- on LP T cells was very low, which meant CD11b+ T cells was not counter population of CD28+ T cells on laminar propria.
Fig. 1. Expression of CD4+CD11b+ on LP T cells. CD11b surface antigen expression on LP T cells in patients with IBD is significantly higher than normal (p=0.03).
Park, et al. CD4 CD11b Antigen on Lamina Propria T Cells of Inflammatory Bowel Disease 497
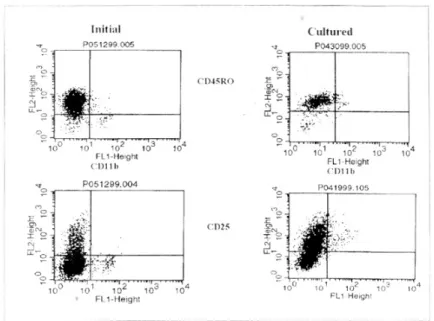
수술 조직에서 분리 직후 CD11b+인 모든 세포에서 CD11b/CD18 (Mac-1)을 보이고, CD45RO는 일부의 세포에서 발현하며, IL-2 Receptor (CD25, IL-2R)는 발현하지 않았으나, 7-10일간의 배양 후 모든 CD11b+
T세포에서 CD11b/CD18 (Mac-1), CD45RO 및 IL- 2R을 발현하였다(Fig. 3).
3. 7- 10일간 배양 후의 CD11b 표면항원 발현율의 변화
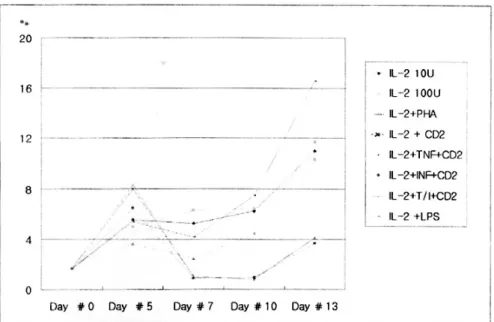
IL-2를 첨가한 배지에서 7-10일 동안 배양 후 CD11b 표면항원의 발현율은 평균 11.8%로 증가하 였으며, 정상 대조군에서 13.4%, 크론병군에서 11.7%,
Fig. 3. The superficial antigen characteristics of CD11b+ T Cell. Initially few cells of CD11b+ T cells showed CD25 (IL-2R) and CD45RO, on cultured state almost CD11b+ T cells expressed CD25 and CD45RO.
Fig. 4. Increased expression of CD4+CD11b on LP T cells after culture with rIL-2 10 U/ml. On 7 to 10 days, the mean rate of CD11b expression was 11.8%. However, there was no significant difference between IBD and normal.
498 대한소화기학회지 : 제 36 권 제 4 호 2000
궤양성 대장염군에서 10.3%로 각 군 간의 유의한 차이는 없었다(Fig. 4).
Fig. 5에서 보이는 바, 다양한 자극제를 배양액에 첨가하였으나 IL-2 단독 첨가군과, IL-2와 Lipo- polysaccharide (LPS, E coli 055:B, Sigma) 첨가군이 IL-2와 PHA, CD2, TNF-α, INF-γ, TNF-α+INF-γ 복합 첨가군에 비해 높은 CD11b 발현을 보였다. 13 일 이상 배양시에는 배양 개시일에 비해 20% 미만 의 세포만이 생존하고 이들의 경우 CD11b 의 발현 율은 배양 7일째에 비교하여 2배 이상 증가하였다.
4. CD4+CD11b+ T 림프구의 세포 내 사이토카인 발현 양상
1) IN F - γ와 T NF - α의 세포 내 발현
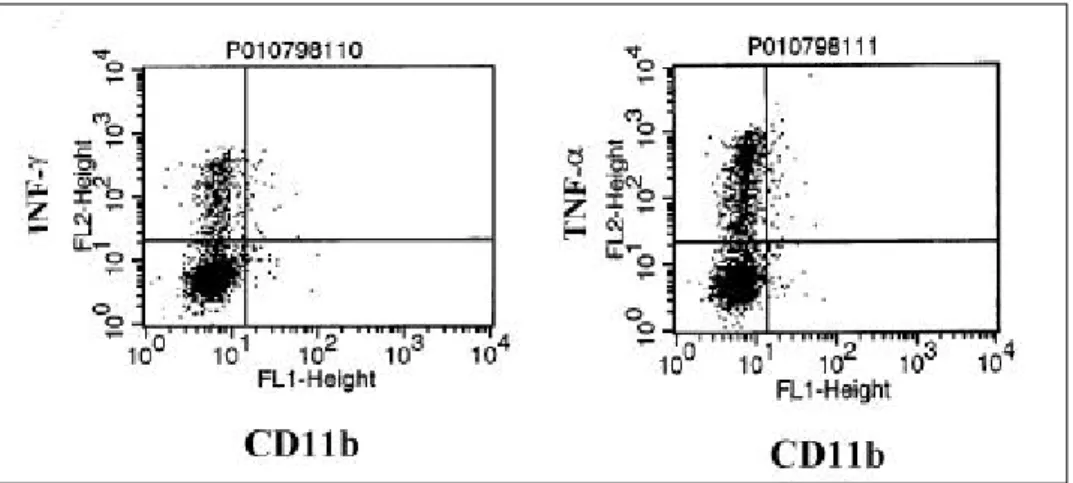
세포 내 INF-γ와 TNF-α는 CD11b 발현군의 50-60%에서 flow cytometer상에 매우 밝게 발현되 었으나, CD11b 발현군과 비발현군 사이에 유의한 차이는 없었다(Fig. 6).
2) IL- 10의 세포 내 발현
IL-10의 세포 내 발현을 얻기 위해서는 적절한
배양 조건이 필요하였다. 한 가지 조건은 CD11b의 발현율을 높이기 위해 IL-2 (10-100 U/ml)를 첨가한 배양액에서 7-10일간 배양 후에 40시간 동안 CD2 와 CD28으로 자극하고, 이 기간의 마지막 16-18시 간 동안 Monensin (2 μM/ml)을 첨가한 후 IL-10의 염색을 시행하였다. 다른 조건으로는 rIL-2 (10-100 U/ml)와 PHA (1 μg/ml)를 첨가하여 7일간 배양 후, 2일간 IL-2 (10-20 U/ml)만을 첨가한 배지에서 휴식 시킨 후 PMA (25 ng/ml)와 Ionomycin (0.5 μg/ml) 및 Monensin을 첨가하여 6시간 동안 자극 후 IL-10 의 염색을 시행하였다.
염색 결과 CD11b 비발현군의 10-15%에서 IL-10 이 밝게 발현되었으나, CD11b 발현군의 약 10%에 서 약한 발현만을 보였다(Fig. 7).
고 찰
염증성 장질환 환자의 말초혈액 T세포에는 활성 화된 림프구의 표지자의 발현이 증가되어 있고 이 는 질환의 활동도와 비례함이 알려져 있다. 성인의 CD11b+ T세포의 분포를 보면 말초혈액에서 20%,
Fig. 5. The change of CD4+CD11b+ expression on LP T cells by variable culture condition. In CD11b expression, IL-2 only and IL-2 with LPS added culture condition are superior to PHA, CD2, TNF-α and INF-γ.
박영숙 외 1인. 염증성 장질환 환자의 점막 고유층 T림프구에서 CD4CD11b 표면항원 발현양상 499
간에서 32%, 비장에서 16%로 비교적 높은 분포를 보이고, 림프절, 갑상선, 대장, 폐에서는 매우 드물 게 관찰되며 연령이 높아질수록 점차 많아진다.6 CD11b 발현과 CD28의 발현이 서로 상반되게 발현 됨이 보고되었고,7,13 정상인의 말초혈액에서 95%의 CD4+ T세포 및 50%의 CD8 T세포가 CD28+
CD11b-의 표현형을 가지며, CD4+ T세포의 1-5%
CD8+ T세포의 50%가 CD28-CD11b+을 보인다.6,14 T림프구의 활성화를 위해 T세포 수용체와 CD28의 공동 자극이 필요하고 T세포 수용체 단독 자극은 T
세포의 anergy 혹은 아포토시스를 유발하지만, CD4+CD28-CD11b+ T세포군에서는 CD3 단독 자극 에 의해서도 활성화되고, HLA-DRB1*0401과 DR1 을 가진 사람이 높은 CD28-CD11b+ 발현을 보여 자 가면역과 연관이 있을 것으로 주장하였다.7 류마치 스양 관절염 환자의 말초혈액에서 CD4+CD28- T세 포의 발현율은 비슷한 연령의 정상인에 비해 증가 되어 있고,15 이들 세포군은 IL-2를 중단해도 아포토 시스가 일어나지 않으며, 자가반응을 보이므로 질환 의 병태생리에 영향을 미칠 것으로 보고하였다.8 그 Fig. 7. Intracellular anti-inflammatory cytokine expression of CD4+CD11b+ T cells. Anti-inflammatory
cytokine IL-10 was expressed brightly on some of CD11b- T cells and expressed only borderline brightness on CD11b+ T cells. Blocking was obtained by using purified IL-10 antibodies.
Fig. 6. Intracellular cytokine expression of CD4+CD11b+ T cells. Intracellular expression of INF-γand TNF-α was not specific between CD11b+ and CD11b- T cells.
500 The Korean Journal of Gastroenterology : Vol. 36, No. 4, 2000
러나 염증성 장질환 환자의 말초혈액에서의 CD28- CD11b+ T림프구는 15-20%이고, 이 가운데 95% 이 상의 T세포가 CD8+CD28-CD11b+이고 CD4+CD28- CD11b+ T림프구는 1-5%로 정상인과 유사하였다.
염증성 장질환 환자의 점막 고유층 T림프구에서 CD4+CD11b+는 정상 점막 고유층 T림프구보다 높 은 발현을 보였으나, CD11b+군과 CD28-군 사이의 상관관계가 없어 점막 고유층의 CD4+CD11b+ T림 프군이 CD4+CD28- T림프군과 동일하지 않은 림프 군임을 시사하였다. CD11b는 수술 조직에서 분리한 직후보다 배양 후에 발현율이 매우 증가하였고, 이 들 세포 모두가 CD45RO를 발현하는 기억세포이며, IL-2R을 발현하는 활성화된 림프구였다. CD11b를 발현하는 T세포가 배양 후에 증가하는 것은 CD11b 의 발현이 림프구의 활성화와 동반된다는 의미를 가지며 2주 이상 배양 후 20% 미만의 세포만이 생 존하는 상황에서 CD11b를 발현하는 세포는 2배 이 상 증가를 보이는 것은 이들 세포가 아포토시스에 내성을 가진 세포군이라는 사실을 암시한다.
말초혈액에서 CD11b+ T세포의 역할은 아직 충 분히 밝혀져 있지 않으나 억제 T세포 활성도 (suppressor T cell activity)를 보이고,16,17 lympho- cytic choriomeningitis virus (LCMV) 감염시 세포독 성 T림프구 활성화의 표지자이며,18 LCMV 감염 후 지연과민반응에서 중요한 역할을 하고, 염증 부위로 T세포 유도(homing)의 역할을 함이 발표되었다.19 CD11b/CD18이 혈관내피세포에 단핵구 및 중성구 를 부착시키는 데 관여함이 알려져 있으며 염증성 장질환 환자의 혈액 내 단핵구 및 호중구에서 CD11b/
CD18의 발현이 증가되어 있고,9 CD11b/CD18에 대 한 항체를 투여하여 TNBS에 의한 장염치료에 효과 적임을 보고하여,10 염증성 장질환의 병태생리에 중 요 부분인 염증 조직으로 단핵구와 중성구의 유입 에 CD11b/CD18가 관여함을 알 수 있다.
ICAM-1, iC3b, fibrinogen 및 응고인자 X 등이 CD11b의 자연 배위자(natural ligands)이다.5 ICAM- 1은 당단백으로 원래 제한된 세포에서만 약하게 발 현되고, IL-1, TNF, INF와 같은 염증성 사이토카인 에 의해 유도되며, 중성구의 혈관내피세포 부착, T 세포의 항원제시세포 부착, T세포와 T세포 및 T세
포와 B세포의 상호작용에 관여한다.20,21 염증성 장 질환 환자에서 혈관내피세포 및 점막 고유층 단핵 세포가 ICAM-1을 발현하고, 활동성 크론병에서 정 상에 비해 높은 ICAM-1 농도를 보이며, 스테로이드 치료 후 농도가 떨어진다고 보고되었다.20,21 또한 면 역화학염색으로 염증성 장질환 환자의 대장에서 국 소적으로도 ICAM-1이 상향 조절되어 있음이 증명 되었다.20,21 이는 CD11b+ T림프구의 발현이 염증성 장질환 환자의 점막 고유층에서 증가하는 것과도 연관성이 있음을 시사한다. 세포접합분자로서 CD11/
CD18는 식세포 수용체로 그람음성 세균의 LPS 성 분에 대한 세포반응에 관여하고, 내독소혈증 때 사 이토카인의 유도뿐 아니라 식작용, 세포 내 저류 및 침입 세균 사멸에도 관여함이 밝혀졌다.22 염증성 장 질환 환자의 혈중 호중구를 LPS로 자극하면 정상인 이나 급성 장염 환자에 비해 높은 전염증 사이토카 인의 분비를 보이고 염증성 장질환의 활동성과도 밀접한 관련이 있음을 볼 때,23 LPS 자극에 의해 점 막 고유층 T림프구의 CD11b 발현이 증가되는 것은 CD11b+ T림프구의 기능이 LPS와 연관될 것으로 생각된다.
IL-10은 대표적인 항염증 분자로 대식세포,24 단 핵구,25 T림프구26 및 호중구23로부터 분비되는 전염 증 사이토카인의 분비를 억제함이 밝혀졌다. IL-10 이 Th1의 증식을 억제하여 IL-2 및 INF-γ의 생성 을 억제한다고 보고되었고,27-29 이후 Th2 림프구에 의한 IL-4와 IL-5의 발현을 억제함이 밝혀졌다.30 IL-10은 또한 자연살해세포에 의한 IFN-γ와 TNF- α의 분비뿐 아니라 대식세포에 의한 growth factor, chemokine (MIP-1α)도 억제함이 보고되었다.31 사람 에서는 활성화된 림프구와 대식세포가 IL-10의 중 요 분비원으로서,25 고농도의 IL-10이 CD4+CD45RO+인 기억 T세포에 의해 분비되나 세포독성 T세포, 비만 세포, B세포, 각질세포, 호산구 및 다양한 종양세포 에 의해서도 분비된다.31 활성화된 점막 고유층 T림 프구는 활성화된 말초혈액 T림프구에 비해 상대적 으로 많은 양의 IL-10을 분비한다. 이러한 증강 효 과는 점막 고유층이 CD45RO+기억 T세포의 분포가 많을 뿐 아니라 CD2 자극에 의한 감수성이 높은 것 과도 연관된다.32 IL-10은 전염증 사이토카인의 전사
Park, et al. CD4 CD11b Antigen on Lamina Propria T Cells of Inflammatory Bowel Disease 501
후 비교적 늦게 나타나는데, 이는 전염증 사이토카 인이 IL-10 생성을 위한 신호임을 시사한다.31 염증 성 자극이 전염증 사이토카인의 분비를 유도하는 한편 IL-10의 생성을 자극하여 진행되고 있는 면역 세포들의 활성화와 염증반응을 소멸시키므로 면역 계의 균형이 유지되고 있음을 암시한다. 그러므로 IL-10의 결핍은 염증반응을 지속 및 증폭시킬 수 있 으며, 이는 IL-10 결실 실험쥐에서 빈혈, wasting syndrome 및 궤양성 대장염과 유사한 염증성 장질 환이 발생한 것으로 증명되었다.33 국내에서도 궤양 성 대장염 환자군에서 대조군에 비해 혈청 IL-10의 농도가 유의하게 높았음이 보고되었다.34 IL-10 존재 하에 사람의 CD4+ T세포를 반복 자극하면, 저증식 성을 갖고 고농도 IL-10을 생성하며 IL-4는 생성하 지 않고 IL-2만 소량 생성하는 CD4+ T세포군, 즉 조절 T세포(T regulatory cells)라 불리는 CD4+ T세 포 아군을 유도하였는데, 이들은 항원 특이적인 면 역반응을 억제하고, 병적 면역반응을 적극적으로 하 향 조절하는 역할을 함이 보고되었다.3 CD4+CD11b+
T세포의 세포 내 사이토카인의 발현을 통해 이 세 포들의 기능을 규명하고자 하였으며, 전염증 사이토 카인인 TNF-α 및 INF-γ는 유세포 분석 결과 CD11b+군의 50%에서 매우 밝게 발현되었으나 CD11b-인 군과 의미 있는 차이를 발견할 수 없었 고, 항염증 사이토카인인 IL-10에 있어서는 CD11b+
군의 10%에서 약한 발현만을 보여 조절 T세포의 가능성은 매우 적다고 생각된다.
그러나 말초혈액의 T림프구는 E. coli의 내막단백 과 세포질단백에 대해 모두 증식하는 데 반해, 장점 막의 T림프구는 오직 순수하게 정제된 항원에만 반 응하고, 이들의 조합에는 반응하지 않는 특성을 보 이므로,35 장점막 고유층의 CD11b+ T림프구가 고농 도의 IL-10을 발현할 수 있는 적절한 조건을 만족하 지 못하여 약한 발현만을 보였을 가능성도 배제할 수 없어 향후 CD11b+ T림프구의 자극에 대한 연구 가 계속되어야 할 것으로 기대한다.
요 약
목적: CD11b/CD18은 β2 세포접합분자로 주로
단핵구, 중성구 및 자연살해세포에서 발현되고, 림 프구에서는 CD8+ T림프구의 50%에서, CD4+T림프 구에서는 5% 미만의 제한된 발현을 보인다. 말초혈 액의 CD4+CD28-림프군이 CD11b를 발현하며, CD3 단독 자극에 의해서도 활성화되고 CD3에 의해 2차 자극을 해도 anergy를 보이지 않고, 아포토시스에도 저항을 보이는 림프군으로 보고하였다. 염증성 장질 환 환자의 혈액 내 단핵구 및 과립세포에서 CD11b/
CD18의 발현이 증가되어 있음이 보고되었다. 염증 성 장질환 환자의 점막 고유층 T림프구에서 CD11b 의 발현 양상 및 세포 내 사이토카인 염색을 통하여 CD11b 양성 림프구의 기능을 규명해 보고자 본 연 구를 시행하였다. 대상 및 방법: 대장 조직은 UCLA Cedars-Sinai Medical Center에서 소장 및 대 장절제술을 받은 28예(남자; 22예, 여자; 6예)의 환 자로부터 얻었으며, 대장암 12예와 크론병 9예 및 궤양성 대장염 7예였다. 대장암 환자의 평균 연령은 67.2세(49-82)이고, 염증성 장질환 환자의 평균 연령 은 41.2세(27-56)였다. 수술 조직으로부터 점막 고유 층 T림프구를 분리하고, 단클론항체를 첨가한 후 면 역형광염색을 시행하여 CD4+CD11b+ T림프구의 발현 양상을 관찰하고, TNF-α, INF-γ, IL-10 등의 세포 내 사이토카인 염색을 시행하였다. 결과: 점막 고유층 T세포의 CD4+CD11b+ 발현율은 정상 대조 군(12예)에서 1.57±0.8%, 크론병 환자군(9예)에서 3.56±0.7%, 궤양성 대장염 환자군(7예)에서 4.42±
1.2%로서 염증성 장질환 환자군이 대조군에 비해 유의한 발현율 증가를 보였다(p=0.03). IL-2를 첨가 한 배지에서 7-10일 동안 배양 후 CD11b 표면항원 의 발현율은 평균 11.8%로 증가하였으며, 정상 대 조군에서 13.4% 크론병 환자군에서 11.7%, 궤양성 대장염 환자군에서 10.3%로 각 군 간의 유의한 차 이는 없었다. INF-γ, TNF-α 등의 세포 내 전염증 사이토카인의 발현은 CD11b+군과 CD11b-군 사이 에 유의한 차이가 없었으며, 항염증 사이토카인인 IL-10의 세포 내 발현은 CD11b-군의 10-15%에서 IL-10이 밝게 발현되었으나, CD11b+군의 약 10%에 서 약한 발현만을 보였다. 결론: 대장점막 고유층 T 세포의 CD4+CD11b+의 발현율은 정상 점막에 비해 염증성 장질환 환자에서 높게 발현되었으나, 세포
502 대한소화기학회지 : 제 36 권 제 4 호 2000
내 사이토카인 염색 결과는 CD4+CD11b+ T 림프구 에서 특이한 소견을 찾지못하였다.
색인단어: 점막 고유층 T세포, CD11b/CD18, INF- γ, TNF-α, IL-10
감사의 글
본 연구가 진행되는 동안 아낌 없는 조언을 해주 신 Cedars-Sinai Inflammatory Bowel Disease Center 의 John Prehn, Richard Deem, Carol Landers외 연구 원 여러분께 진심으로 감사드립니다.
참 고 문 헌
1. Sartor RB. Current concepts of the etiology and pathogenesis of ulcerative colitis and Crohn' s disease. Gastroenterol Clin North Am 1995;24:475- 507.
2. Papadakis KA, Targan SR. Current theories on the causes of inflammatory bowel disease. Gastroen- terol Clin North Am 1999;28:283-296.
3. Groux H, Garra AO, Bigler M, et al. A CD4+ T cell subset inhibits antigen specific T cell responses and prevents colitis. Nature 1997;389:737-742.
4. Ingalls RR, Arnaut MA, Golenbock DT. Outside-in signaling by lipopolysaccharide through a tailless integrin. J Immunol 1997;159:433-438.
5. Springer TA, Adhesion receptors of the immune system. Nature 1990;346:425-430.
6. Yamada H, Martin PJ, Bearn MP, et al. Mono- clonal antibody 9.3 and anti-CD11 antibodies define reciprocal subsets of lymphocytes. Eur J Immunol 1985;15:1168-1175.
7. Chapman A, Stewart SJ, Nepom GT, et al.
CD11b+CD28-CD4+ Human T cells. J Immunol 1996;157:4771-4780.
8. Schirmer M, Vallejo AN,Weyand CM, Goronzy JJ.
Resistance to apoptosis and elevated expression of bcl-2 in clonally expanded CD4+CD28- T cells from Rheumatoid Arthritis patients. J Immunol
1998;161:1018-1025.
9. Collins CE, Davis C, Macey MG, McCarthy DA, Rampton DS. Increased expression of circulating leucocyte adhesion molecules CD11b and CD18 in inflammatory bowel disease. Gastroenterology 1996;
110(abstr):A885.
10. Palmen MJ, Dijkstra CD, van der Ende MB, Pena AS, van Rees EP. Anti-CD11b/CD18 antibodies reduce inflammation in acute colitis in rats. Clin Exp Immunol 1995;101:351-356.
11. Bull DM, Bookman MA. Isolation and functional characterization of human intestinal mucosal lym- phoid cells. J Clin Invest 1977;59:966-970.
12. Shanahan F, Brogan M, Targan S. Human mucosal cytotoxic effector cells. Gastroenterology 1987;92:
1951-1959.
13. Hoshino T, Yamada A, Honda J., et al. Tissue- specific distribution and age-dependent increase of human CD11b+ T cells. J Immunol 1993;151:
2237-2246.
14. Damle NK, Mohagheghpour N, Hasen JA, Engleman EG. Alloantigen-specific cytotoxic and suppressor T lymphocytes are derived from phenotypically distinct precursors. J Immunol 1983;
131:2296-2302.
15. Schmidt D, Goronzy JJ, Weyand CM. CD4+
CD7-CD28- T cells are expanded in rheumatoid arthritis and are characterized by autoreactivity. J Clin Invest 1996;97:2027-2037.
16. Li SG, Ottenhoff THM, van den Elsen P, et al.
Human suppressor T cell clones lack CD28. Eur J Immunol 1990;20:1281-1292.
17. Koide J, Engleman EG. Differences in surface phenotype and mechanism of action between alloantigen-specific CD8+ cytotoxix and suppressor T cell clones. J Immunol 1990;144:32-40.
18. Macfarland HI, Nahill SR, Maciaszek JW, Welsh RM. CD11b (Mac-1): a marker for CD8+ cytotoxic T cell activation and memory in virus infection. J Immunol 1992:149:1326-1333.
19. Nielsen HV, Christensen JP, Anderson EC, Marker O, Thomsen AR. Expression of tpye 3 complement
박영숙 외 1인. 염증성 장질환 환자의 점막 고유층 T림프구에서 CD4CD11b 표면항원 발현양상 503
receptor on activated CD8+ T cells facilitate homing to inflammatory sites. J Immunol 1994;153:
2021-2028.
20. Sands BE. Biologic therapy for inflammatory bowel disease. Inflamm Bowel Dis 1997;3:95-113.
21. Jones SC, Banks RE, Haidar A, et al. Adhesion molecules in inflammatory bowel disease. Gut 1995;36:724-730.
22. Flaherty SF, Golenbock DT, Milham FH, Ingalls RR. CD11/CD18 leukocyte integrins: new signaling receptors for bacterial endotoxin. J Surg Res 1997;
73:85-89.
23. Nikolaus S, Bauditz J, Gionchetti P, Witt C, Losch H, Schreiber S. Increased secretion of pro-inflam- matory cytokines by circulating polymorphonuclear neutrophils and regulation by interleukin 10 during intestinal inflammation. Gut 1998;42: 470-476.
24. Fiorentino DF, Zlontnik A, Timothy RM, Howard M, O' garra A. IL-10 inhibits cytokine production by activated macrophages. J Immunol 1991;147:
3815-3822.
25. de Waal Malefyt R, Arbrams J, Bennet B, Figdor CG, de Vries JE. IL-10 inhibits cytokine synthesis by human monocytes: An autoregulatory role of IL-10 produced by monocytes. J Exp Med 1991;
174:1209-1220.
26. Ralph P, Nakoinz I, Sampson JA, et al. IL-10, T lymphocyte inhibitor of human blood cell produc- tion of IL-1 and tumor necrosis factor. J Immunol 1992;148:808-814.
27. de Waal Malefyt R, Haanen J, Spits H, et al. IL-10 and viral IL-10 strongly reduce antigen specific human T cell proliferation by diminishing the
antigen presenting capacity of monocytes via downregulation of class II major histocompatibility complex expression. J Exp Med 1991;174:915-924.
28. Taga K, Tosato G. IL-10 inhibits human T cell proliferation and IL-2 production. J Immunol 1992;148:1143-1148.
29. Ding L, Shevach EM. IL-10 inhibits mitogen induced T cell proliferation by selectively inhibiting machrophage costimulatory function. J Immunol 1992;148:3133-3139.
30. Prete GD, Carli MD, Almerigogna F, Giudizi MG, Biagiotti R, Romagnani S. Human IL-10 is produced by both type 1 helper and type 2 helper T cell clones and inhibits their antigen specific proliferation and cytokine production. J Immunol 1993;150:353-360.
31. Borrish L. IL-10: Evolving concepts. J Allergy Clin Immunol 1998;10:293-297.
32. Braunstein J, Qiao L, Autschbach F, Schurmann G, Meuer S. T cells of the human intestinal laminar propria are high producers of IL-10. Gut 1997;41:
215-220.
33. Kuhn R, Lohler J, Rennick D, Rajewsky K, Muller W. IL-10 deficient mice develop chronic entero- colitis. Cell 1993;75:263-274.
34. 민상운, 김태연, 유상두 등. 궤양성 대장염 환자에서 혈청 Interleukin-10과 Interleukin-12 측정에 의한 Th1/Th2 반응의 평가. 대한소화기학회지 2000;35:
32-38.
35. Khoo UY, Proctor IE, Macpherson AJS. CD4+ T cell down regulation in human intestinal mucosa. J Immunol 1997;158:3626-3634.