358 책임저자:박동일, 614-052, 부산시 진구 양정동 산 45번지
동의대학교 한의과대학 내과학교실 Tel: 051-850-8650, Fax: 051-853-4036 E-mail: [email protected]
접수일:2011년 11월 30일, 1차 수정일:2011년 12월 5일, 2차 수정일:2011년 12월 8일, 게재승인일:2011년 12월 12일
Correspondence to:Dong Il Park
Department of Internal Oriental Medicine, Dongeui University College of Oriental Medicine, San 45, Yangjung-dong, Busanjin-gu, Busan 614-052, Korea
Tel: +82-51-850-8650, Fax: +82-51-853-4036 E-mail: [email protected]
보중익기가미길경탕에 의한 A549 인체 폐암세포의 Apoptosis 유도
동의대학교 한의과대학 1생화학교실 및 2내과학교실,
3대학원 바이오물질제어학과 및 블루바이오소재개발센터
배종훈1ㆍ신동역1ㆍ박 철1ㆍ최영현1,3ㆍ박동일2
Induction of Apoptosis by Bojungikgigilkyung-tang in Human Lung Cancer A549 Cells
Jong Hoon Bae1, Dong Yeok Shin1, Cheol Park1, Yung Hyun Choi1,3 and Dong Il Park2
Departments of 1Biochemistry and 2Internal Oriental Medicine, Dongeui University College of Oriental Medicine, Busan 614-052, 3Department of Biomaterial Control (BK 21 Program), Graduate School and Blue-Bio Industry
Regional Innovation Center, Dongeui University, Busan 614-714, Korea
Bojungikkigamikilkyung-tang (BJIKTP) is a traditional oriental herbal formulation consisting of crude ingredients extracted from nine herbs. This prescription has been widely used for patients suffering from a lung disease in Oriental medicine. In the present study, it was examined the biochemical mechanisms of apoptosis induced by BJIKTP in human non-small-cell lung cancer (NSCLC) cells. It was found that exposure to BJIKTP was found to result in a concentration-dependent growth inhibition by induction of apoptosis, as evidenced by chromatin chondensation, DNA fragmentation and the accumulation of sub-G1 phase, in NSCLC A549 cells, but not in NCI-H460 cells. Apoptosis induction of A549 cells by BJIKTP was connected with a modulation of Bcl-2 and inhibitors of apoptosis (IAP) family members, and up-regulation of death-receptor related members. BJIKTP treatment also induced proteolytic activation of caspase-3, -8 and -9, and degradation of caspase-3 substrate proteins including poly (ADP-ribose) polymerase and phospholipase C γ-1. In addition, BJIKTP-induced apoptosis is associated with the reduction of mitochondrial membrane potential (MMP) and cleavage of Bid proteins. Taken together, the results indicate that BJIKTP may be a potential chemotherapeutic agent for use in the control of human lung cancer and that further studies are needed for the identification of the active compounds. (Cancer Prev Res 16, 358-370, 2011)
Key Words: BJIKTP, A549, Apoptosis, Caspase, Bid
서 론
보중익기탕(補中益氣湯, Bojungikki-tang)은 한의학에 서 체력을 보강해주는 대표적인 처방 중의 하나이다. 특
히 과로나 소화 불량에 의한 발열이나 땀이 많이 나는 것을 억제하여 기를 상승시켜주는 처방으로 널리 이용 되고 있다.1~3) 보중익기탕은 1첩 기준으로 황기(黃耆) 6 g, 인삼(人蔘) 4 g, 백출(白朮) 4 g, 감초(甘草) 4 g, 당귀(當 歸) 2 g, 진피(陳皮) 2 g과 술에 절인 시호(柴胡) 및 승마
(升摩)가 각 1.2 g으로 구성되어 있다. 최근 보중익기탕에 길경(桔梗)이 함유된 보중익기가미길경탕(Bojungikkiga- mikilkyung-tang, BJIKTP)이 인체 폐암세포의 이동과 침윤 을 억제할 수 있음을 보고된바 있는데, 이는 matrix metal- loproteinase 활성의 감소와 tight junctions의 기능 강화와 연관성이 있었다.4) 길경(桔梗)은 초롱꽃과(Campanula- ceae)에 속하는 다년생 초본인 도라지(Platycodon grandi- florum)의 뿌리를 건조한 것으로서 맛은 쓰고 매우며, 폐 에 작용하여 가래가 많고 호흡이 불편한 증상을 치료할 뿐만 아니라 폐를 맑게 하고 답답한 가슴을 풀어주며 뱃 속의 찬 기운을 풀어주어 기침을 멈추게 하는 것으로 알 려져 있다.5,6) 또한 길경은 모든 약재의 상승작용을 유발 할 뿐만 아니라 인후통, 감기로 인한 기침, 가래, 코막힘, 천식, 기관지염증, 흉막염, 두통, 오한 및 편도선염 등에 약효가 있는 것으로 보고되고 있다.7) 따라서 보중익기가 미길경탕은 소화기계 및 폐의 기운을 증가시켜 면역력 을 강화시키는 원래의 처방인 보중익기탕에 염증이나 종양 등을 억제하는 길경을 첨가하여 약효를 증가시킨 처방으로서 면역력 저하로 인한 상부의 제반 증상에 효 능이 우수할 것으로 기대되어지는 처방이다.
정상적인 세포는 세포내 조절기능에 의해서 분열하며 성장하고 사멸하면서 개체 수준에서 세포 수의 항상성 을 유지한다. 이러한 세포의 증식과 암의 발생 측면에서 세포주기 조절의 교란 및 세포의 사멸 기전 이상에 의하 여 세포가 끊임없이 증식되는 것을 암세포라고 할 수 있 다.8) 세포의 여러 가지 활동 중 세포사멸은 크게 apoptosis 와 necrosis로 구분될 수 있으며, 이는 세포의 형태적 및 생화학적 특성에 의하여 구분된다. Necrosis는 세포 주위 의 환경이 급격하게 변화함으로써 세포의 팽창, 이온 농 도의 변화 및 물의 유입 등과 같은 조절되지 않는 상태 로 유발되는 세포 죽음의 한 형태로서 에너지 비의존적 이며, necrosis가 유발된 세포의 내용물이 유출 또는 방출 되어 인접한 세포에 영향을 미치게 되며 염증성 반응을 촉진하기도 한다.9,10) 이와 반대로 apoptosis는 programmed cell death라고 불리는 생리학적 과정이며, apoptosis의 특 징으로는 mitochondria의 기능 변화, 세포의 수축, 염색질 응축, 핵의 단편화, 세포막의 수포화 현상, cysteine-related protease인 caspases의 활성화, 세포 표면에 phosphatidylser- ine의 발현 및 apoptotic body의 형성 등과 같은 형태적 또 는 생화학적인 변화를 동반하고 있으며, 다양한 유전자 발현의 증가 또는 감소 등과 같은 상호작용에 의해 유발 되는 것으로 알려져 있다.11,12) 또한 apoptosis는 대부분의 조직에서 손상을 입었거나 감염된 세포들을 제거하는 중요한 과정으로서 이러한 apoptosis 과정이 실패하게 되
면 암과 같은 여러 가지 질병의 원인이 되기 때문에 apoptosis는 암화 과정의 여러 단계에서 암을 치료하는 중 요한 표적이 되고 있다.
Apoptosis를 통한 세포의 죽음에는 세포의 분해에 관여 하는 효소를 활성화시키는 caspase라고 불리는 effector protease의 활성이 필요하다. 이러한 과정은 extrinsic path- way라고 하는 death receptor pathway와 intrinsic pathway라 고 하는 mitochondrial pathway와 같은 과정에 의해 유발된 다.13) Extrinsic pathway는 mitochondria 비의존적인 apopto- sis 과정으로서 apoptotic ligand가 세포 막에 존재하는 death receptor와의 결합으로 시작된다.14) Ligand가 death receptor와 결합하게 되면 intracellular plasma membrane에 death-inducing signaling complex (DISC)를 형성하게 되고, 이러한 pro-apoptotic 신호에 의하여 initiator caspase인 cas- pase-8 및 -10을 활성화시켜 effector caspase인 caspase-3, -6 및 -7의 활성화와 그에 따른 DNA의 단편화 현상을 유발
시킨다.15~17) Mitochondria 의존적 apoptosis 과정인 intrinsic
pathway는 mitochondrial membrane의 permeability 변화에 의하여 여러 종류의 inter-mitochondrial apoptotic molecule 들이 세포질로 방출됨으로서 시작된다.14) 가장 중요한 pro-apoptotic molecule 중 하나인 cytochrome c가 mitochon- dria에서 세포질로 방출되면 pro-caspase-9 및 Apaf-1 (apoptosis protease activating factor-1)이 apoptosome이라는 중합체를 형성하여 caspase-9 및 -3 등을 활성화시킨
다.18,19) 즉 mitochondria는 여러 세포에서 apoptosis가 유발
되는 동안 손상되지 않고 남아서 initiator 작용을 하는 반 면에 necrosis가 유발이 되면 팽창되어 파괴되므로 apop- tosis 유발 과정에 있어서 매우 중요한 역할을 하고 있 다.20,21)
본 연구에서는 BJIKTP가 인체 폐암세포의 apoptosis 유 발에 미치는 생화학적 기전의 해석을 위하여 apoptosis 과 정 중 extrinsic 및 intrinsic pathway에 관여하는 몇 가지 중 요한 유전자들의 발현 변화를 조사하여 유의적인 결과 를 얻었기에 이를 보고하는 바이다.
재료 및 방법 1. 암세포의 배양
본 실험에 사용한 A549 및 NCI-H460 인체폐암세포는 American Type Culture Collection (Rockville, MD, USA)에서 분주 받아 사용하였으며, 암세포의 배양을 위해 90%의 RPMI-1640 배지(Gibco BRL, Grand Island, NY, USA), 10%
의 우태아혈청(fetal bovine serum, FBS), 1%의 penicillin 및 streptomycin (Biofluids, Rockville, MD, USA)이 포함된 성장
배지를 사용하여 배양하였다. 세포는 37oC, 5% CO2 조건 하에서 배양하였으며, 성장배지의 교환은 매 48시간마 다 해주었고, 세포 수의 증식에 따른 과밀도 현상을 해소 하기 위하여 0.05% trypsin-EDTA (Gibco BRL)를 처리하여 세포를 부유시킨 다음 세포배양용 페트리 접시로 옮겨 배양하였다.
2. BJIKTP의 추출 및 처리
본 연구에 사용된 BJIKTP는 부산광역시 소재 화평한 의원에서 제공 받았으며, 100 g을 1,000 ml의 증류수에 3시간 이상 끓인 후, 3,000 rpm으로 20분간 원심 분리시 켜 침전물을 제거하였다. 이를 다시 0.45μm의 여과지를 이용하여 부유 성분을 걸러낸 후 수용성분을 동결 건조 하여 사용하였다. BJIKTP의 처리를 위하여 세포를 0.05% trypsin-EDTA를 이용하여 세포배양용 페트리 접시 로부터 부유시킨 다음 새로운 세포배양용 페트리 접시 에 분주하여 24시간 동안 안정화 시켰다. BJIKTP를 세포 에 처리하기 직전 적정 농도로 성장배지에 첨가하여 녹 인 다음, 0.22μm의 pore size를 가진 주사기용 필터유닛 을 사용하거나 1회용 펌프 필터 유닛을 사용하여 미생물 및 불순물을 걸러낸 다음, 세포의 성장배지를 갈아주면 서 직접 처리하였다.
3. MTT assay를 통한 세포 생존율의 측정
세포 배양용 6 well plate에 A549 및 NCI-H460 세포를 1×104개/ml로 분주하여 안정화시킨 다음 BJIKTP를 배지 에 희석하여 각 well 당 적정농도(0∼4 mg/ml)로 처리하 였다. 일정 시간 경과 후 0.5 mg/ml 농도의 tetrazolium bromide salt (MTT, Amresco, Solon, OH, USA)를 2 ml씩 분 주하고 2시간 동안 CO2 incubator에서 배양시킨 다음 MTT 시약을 제거하고 DMSO를 1 ml씩 분주하여 well에 생성된 formazin을 모두 녹인 후 96 well plate에 200μl씩 옮겨서 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)로 540 nm에서 흡광도를 측정하였다. 측정은 모두 세 번을 하였으며, 그에 대한 평균값과 표준 오차를 Microsoft EXCEL program을 사용하여 분석하였다.
4. 세포형태 관찰 및 DAPI 염색에 의한 핵의 형태 관찰
BJIKTP 처리에 의한 암세포의 전체적인 형태 변화 관 찰을 위하여 암세포를 24시간 동안 안정화시킨 후, 다양 한 농도의 BJIKTP가 희석된 배지에서 48시간 동안 배양 된 암세포를 위상차 현미경을 이용하여 200배의 배율로 각 처리 농도에 따른 형태의 변화를 조사하였다. BJIKTP 처리에 의한 암세포의 apoptosis 유발 여부를 확인하기 위
한 핵의 형태적 변화 관찰을 위하여 동일 조건에서 배양 된 세포를 모은 다음, 37% formaldehyde 용액과 PBS를 1:9의 비율로 섞은 fixing solution을 모아진 세포에 500 μl 첨가하여 충분히 섞은 후, 상온에서 10분 동안 고정 하였다. 2,000 rpm으로 5분간 원심 분리한 후 상층액을 제거하고 PBS 200μl를 넣어서 충분히 섞은 후, slide glass 위에 80μl 정도 떨어뜨려 1,000 rpm에서 5분간 cytospin 하였다. PBS로 2~3회 washing하고 PBS가 다 마르기 전에 0.2%의 Triton X-100 (Amresco)을 첨가하여 상온에서 10 분간 고정하였다. 그 후 다시 PBS로 washing하고 4',6-dia- midino-2-phenylindole (DAPI, Sigma, St. Luis, MO, USA) 용 액을 세포가 고정된 slide glass 위에 적당량을 떨어뜨린 후 빛을 차단하고 상온에서 염색시켰다. 15분 정도 염색 시킨 후, PBS로 DAPI 용액을 충분하게 세척하고 증류수 로 재빨리 세척한 다음 absolute alcohol을 이용하여 탈수 과정을 거친 slide glass 위에 mounting solution을 처리한 후 형광 현미경(Carl Zeiss, Germany)을 이용하여 400배의 배율로 각 농도에 따른 암세포 핵의 형태 변화를 관찰하 였다.
5. DNA fragmentation 분석
특정 세포에 apoptosis가 유발되었을 때 관찰할 수 있는 DNA fragmentation의 분석을 위하여 위와 동일한 조건으 로 배양된 세포를 모아 lysis buffer [5 mM Tris-HCl (pH 7.5), 5 mM EDTA, 0.5% Triton X-100]를 4oC에서 30분간 처리한 다음, 그 상층액에 proteinase K (Sigma)를 0.5 mg/ml의 농도로 처리한 다음 50oC에서 3 시간 동안 반응 시켰다. 그 후 phenol:chloroform:isoamyl alcohol 혼합용 액(25:24:1, Sigma)을 첨가하여 30 분간 inverting한 뒤 원심분리하여 따로 분리한 상층액에 적정량의 isopro- panal (Sigma)과 5 M NaCl를 첨가한 다음 4oC에서 over- night시켰다. 이후 원심분리하여 분리된 DNA pellet을 70% ethanol로 충분히 세척하고 RNase A가 적당량 들어 있는 DEPC water에 녹여 gel loading dye (Bioneer, Daejeon, Korea)를 혼합한 후 1.5% agarose gel에 1 시간 정도 50 V 로 전기영동하였다. 이후 ethidium bromide (EtBr, Sigma)로 염색하여 ultra violet (UV) 하에서 관찰하였다.
6. Flow cytometry 분석
정상 및 BJIKTP가 함유된 배지에서 48시간 동안 배양 시킨 A549 세포를 씻어 내고 0.05% trypsin-EDTA를 처리 하여 부유시킨 다음 2,000 rpm으로 5분간 원심 분리하여 상층액을 버리고 세포들만 모았다. 여기에 다시 PBS를 첨가하여 충분히 씻은 다음 2,000 rpm으로 5분간 원심분
Table 1. Sequence of primers used for RT-PCR
Gene name Sequence
GAPDH Sense
Antisense
5'-CGG AGT CAA CGG ATT TGG TCG TAT-3' 5'-AGC CTT CTC CAT GGT GGT GAA GAC-3'
TRAIL Sense
Antisense
5'-ATG GCT ATG ATG GAG TCC AG-3' 5'-TTG TCC TGC ATC TGC TTC AGC-3'
DR4 Sense
Antisense
5'-CAG AAC GTC CTG GAG CCT GTA AC-3' 5'-ATG TCC ATT GCC TGA TTC TTT GTG-3'
DR5 Sense
Antisense
5'-GGG AAG AAG ATT CTC CTG AGA TGT G-3' 5'-ACA TTG TCC TCA GCC CCA GGT CG-3'
Fas Sense
Antisense
5'-TCT AAC TTG GGG TGG CTT TGT CTT C-3' 5'-GTG TCA TAC GCT TTC TTT CCA T-3'
FasL Sense
Antisense
5'-GGA TTG GGC CTG GGG ATG TTT CA-3' 5'-AGC CCA GTT TCA TTG ATC ACA AGG-3'
Bcl-2 Sense
Antisense
5'-CAG CTG CAC CTG ACG-3' 5'-ATG CAC CTA CCC AGC-3'
Bcl-XL Sense
Antisense
5'-CGG GCA TTC AGT GAC CTG AC-3' 5'-TCA GGA ACC AGC GGT TGA AG-3'
Bax Sense
Antisense
5'-ATG GAC GGG TCC GGG GAG-3' 5'-TCA GCC CAT CTT CTT CCA-3'
XIAP Sense
Antisense
5'-GAA GAC CCT TGG GAA CAA CA-3' 5'-CGC CTT AGC TGC TCT CTT CAG T-3'
cIAP-1 Sense
Antisense
5'-TGA GCA TGC AGA CAC ATG C-3' 5'-TGA CGG ATG AAC TCC TGT CC-3'
cIAP-2 Sense
Antisense
5'-CAG AAT TGG CAA GAG CTG G-3' 5'-CAC TTG CAA GCT GCT CAG G-3'
Survivin Sense
Antisense
5'-GCA TGG GTG CCC CGA CGT TG-3' 5'-GCT CCG GCC AGA GGC CTC AA-3' 리 한 후 상층액만 버리고 남은 세포에 CycleTEST PLUS
DNA REAGENT Kit (Becton Dickinson, San Jose, CA, USA) 를 이용하여 고정 및 염색을 하여 4oC, 암실에서 30분 동 안 반응을 시킨 다음 DNA flow cytometer (Becton Dickin- son)에 적용시켜 형광반응에 따른 histogram을 ModiFit LT (Becton Dickinson) 프로그램으로 분석하였다.
7. Mitochondrial membrane potential (MMP, Δ ψm)의 분석
BJIKTP가 처리된 A549 세포의 MMP 변화 정도를 측정 하기 위하여 적정시간 동안 BJIKTP가 처리된 세포들을 모은 다음 500μl의 PBS를 첨가하여 충분히 섞은 후, 10 μM의 5,5′, 6,6′-tetrachloro-1,1′,3,3′-tetraethyl-imidacarbocya- nine iodide (JC-1, Sigma) 용액을 처리하여 20분 동안 37oC 에서 반응시켰다. 반응시킨 세포를 2,000 rpm으로 원심 분리하여 상층액을 버리고 다시 500μl의 차가운 PBS를 첨가하고 35 mm mesh를 이용하여 단일세포로 분리한 후 DNA flow cytometer에 적용시켜 BJIKTP 처리에 따른 MMP의 변화 정도를 분석하였다.
8. Reverse transcription-polymerase chain re- action (RT-PCR)에 의한 mRNA 발현 변화 분석
동일한 조건에서 배양된 A549 세포들에 TRIzol reagent (Invitrogen Co., Carlsbad, CA)를 4oC에서 1시간 동안 처리 하여 RNA를 분리하였다. 분리된 RNA를 정량한 후, 각각 의 primer (Table 1), DEPC water 그리고 ONE-STEP RT-PCR PreMix Kit (Intron, Korea)를 넣고 Mastercycler gra- dient (Eppendorf, Hamburg, Germany)를 이용하여 증폭하 였다. 각 PCR 산물들의 양적 차이를 확인하기 위하여 1×TAE buffer로 1% agarose gel을 만들고 well 당 각각의 primer에 해당하는 PCR 산물에 DNA gel loading solution을 섞어서 loading한 후 50 V에서 전기영동을 행하였다. 전 기영동으로 DNA 분리가 끝난 gel을 EtBr을 이용하여 염 색한 후 UV 하에서 발현의 차이를 확인하였으며, house- keeping 유전자인 glyceraldehyde-3-phosphate dehydrogenase (GAPDH)를 internal control로 사용하였다.
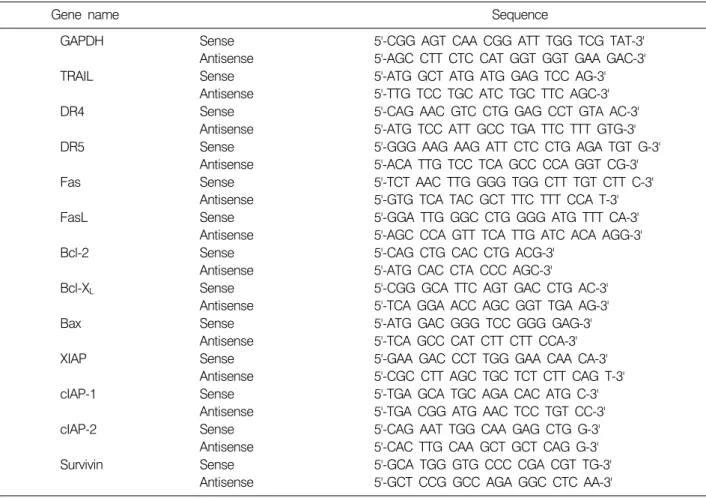
Fig. 1. Effects of BJIKTP on the viability of non-small-cell lung cancer (NSCLC) NCI-H460 and A549 cells. Cells were plated at 1×104 cells per 60-mm plate, and incubated for 24 h. The cells were treated with variable concentrations of BJIKTP for 48 h and the cell viability was measured by the metabolic- dye-based MTT assay. Results are expressed as the means±
S.D. of three independent experiments. Significance was de- termined using a Student’s t-test (*p<0.05 vs. untreated control).
9. Western blot analysis에 의한 단백질 발현 분석
정상 및 BJIKTP가 처리된 배지에서 자란 세포들을 PBS로 씻어 내고 0.05% trypsin-EDTA를 처리하여 부유시 킨 다음 원심분리를 하여 세포를 모았다. 이렇게 모아진 세포에 적당량의 lysis buffer [25 mM Tris-Cl (pH 7.5), 250 mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenymethyl- sulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]를 첨가 하여 4oC에서 1시간 동안 반응시킨 후, 14,000 rpm으로 30분간 원심분리하여 그 상층액을 취하였다. 상층액의 단백질 농도는 Bio-Rad 단백질 정량 시약(Bio-Rad, Her- cules, CA, USA)과 그 사용방법에 따라 정량 한 다음 동량 의 Laemmli sample buffer (Bio-Rad)를 섞어서 sample을 만들 었다. 이렇게 만든 동량의 단백질을 sodium dodecyl sul- phate (SDS)-polyacrylamide gel을 이용하여 전기영동으로 분리하였다. 분리된 단백질을 함유한 acrylamide gel을 ni- trocellulose membrane (Schleicher and Schuell, Keene, NH, USA)으로 electroblotting에 의해 전이시킨 후, 5% skim milk를 함유한 PBS-T (0.1% Tween 20 in PBS)에 담구어 상온에서 1시간 정도 incubation하여 비특이적인 단백질 들에 대한 blocking을 실시하고 PBS-T로 15분(5분간 3번) 정도 세척하였다. 준비된 membrane에 1차 항체를 처리하 여 상온에서 2시간 이상 또는 4oC에서 over night 시킨 다 음 PBS-T로 세척(15분간 1번, 5분간 5번)하고 처리된 1차 항체에 맞는 2차 항체(PBS-T로 1:1,500으로 희석하여 사용)를 사용하여 상온에서 1시간 정도 반응시켰다. 다 시 PBS-T로 세척(10분간 4번)하고 Enhanced Chemilumi- noesence (ECL) 용액(Amersham Life Science Corp., Arlington Heights, IL, USA)을 적용시킨 다음 암실에서 X-ray film에 감광시켜 특정단백질의 양을 분석하였다.
10. In vitro caspase-3, -8 및 -9의 활성 변화 측정
BJIKTP 처리에 의한 caspases의 활성화 정도를 알아보 기 위하여 정상 및 BJIKTP가 처리된 배지에서 48시간 배양된 세포를 모은 뒤 단백질을 추출하고 정량하여 각 각 150μg의 단백질을 fluorogenic peptide 기질 100μM이 함유된 extraction buffer [40 mM HEPES (pH 7.4), 20% glyc- erol (v/v), 1 mM EDTA, 0.2% NP-40 and 10 mM DL-DTT]
50μl에 혼합하였으며, microtiter plate에 다시 extraction buffer에 희석하여 각 sample 당 총 volume이 100μl가 되 게 하였다. 실험에 사용된 기질은 caspase-3의 경우에는 Asp-Glu-Val-Asp (DEVD)-p-nitroaniline (pNA)이었고 cas- pase-8의 경우에는 Ile-Glu-Thr-Asp (IETD)-pNA이었으며, caspase-9는 Leu-Glu-His-Asp (LEHD)-pNA였다. 준비된 plate
를 37oC에서 3시간 동안 incubation시킨 후 ELISA reader를 이용하여 405 nm의 흡광도를 이용하여 반응의 정도를 측 정하였다.
11. 통계 처리
모든 실험결과는 평균±표준편차로 표시하였고 Sigma- Plot을 이용하여 Student t-test를 이용하여 통계적 유의성 을 얻었다.
결 과
1. 폐암세포의 생존에 미치는 BJIKTP의 영향
BJIKTP 처리에 의한 인체 폐암세포의 생존율 변화를 측정하기 위하여 MTT assay를 실시하였으며, 이를 위하 여 두 종류의 폐암세포(NCI-H460 및 A549)에 BJIKTP를 1∼4 mg/ml의 농도로 48시간 처리 한 후 얻어진 결과는 Fig. 1에 나타낸 바와 같다. 제시된 결과에서 알 수 있듯 이, A549 세포의 경우 BJIKTP 처리 농도의 증가에 따른 생존율의 저하가 매우 크게 관찰되었는데, 2 mg/ml 처리 군에서 약 25% 정도의 생존율 저하가 관찰되었으며, 3 mg/ml 및 4 mg/ml 처리군에서는 대조군에 비하여 생존 율이 약 50% 및 65% 정도 억제되었음을 확인할 수 있었 다. 그러나 NCI-H460 세포의 경우에서는 A549 세포에 비하여 BJIKTP에 대한 감수성이 상대적으로 낮게 나타
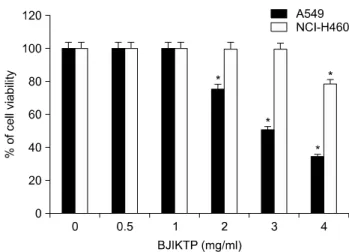
Fig. 2. Effects of BJIKTP on the cellular and nuclear morphological changes in A549 cells. (A) A549 cells were treated with the indicated concentrations of BJIKTP. After incubation for 48 h, the cells were sampled and examined using an inverted microscope (magnification, ×200). (B) The cells grown under the same conditions as (A) were sampled, fixed, and then stained with DAPI.
The stained nuclei were observed under a fluorescent microscope using a blue filter (magnification, ×400).
나, 4 mg/ml 처리군에서 약 22% 정도의 생존율 저하를 보여 주었다. 따라서 BJIKTP에 대한 항암 감수성이 높을 것으로 예상되는 A549 세포를 대상으로 BJIKTP 처리에 따른 증식억제와 연관된 apoptosis 유발 기전에 대한 연구 를 실시하였다.
2. 세포의 형태 및 핵의 형태에 미치는 BJIKTP의 영향
BJIKTP 처리에 따른 세포 생존율의 감소에 따른 A549 폐암세포의 형태적 변화를 관찰한 결과는 Fig. 2A에 나 타낸 바와 같다. 결과에서 알 수 있듯이 전체 세포의 밀 도 감소와 함께 부유 세포의 증가 및 다양한 형태적 변 형을 관찰할 수 있었다. 다음은 BJIKTP 처리에 의한 A549 세포의 증식억제가 apoptosis 유발과 연관성이 있는 지 알아보기 위하여 정상 및 BJIKTP가 처리된 배지에서 배양된 세포를 고정시킨 후 DAPI 염색을 실시하여 핵의 형태변화를 형광현미경하에서 관찰하였다. Fig. 2B에 나 타낸 바와 같이 정상배지에서 배양된 세포의 경우 핵의 형태에 아무런 변화가 나타나지 않았지만 BJIKTPS를 처 리한 경우, 처리 농도가 높아짐에 따라 apoptosis가 일어 난 세포에서 전형적으로 관찰되는 염색질 응축에 의한 apoptotic body의 출현이 증가되었다. 이는 BJIKTP 처리 에 의한 A549 세포의 증식억제가 apoptosis 유발에 의한 것임을 의미하는 결과이다.
3. BJIKTP 처리에 의한 A549 세포의 apoptosis 유발 다음은 apoptosis 유발의 또 다른 직접적인 증거가 되
는 DNA의 단편화 유도 여부를 A549 세포를 대상으로 확인하였다. Fig. 3A에서 나타낸 바와 같이 BJIKTP 처리 농도의 증가에 따라 DNA 단편화의 정도가 매우 증가되 었으며, 이러한 BJIKTP 처리에 의한 apoptosis 유발의 정 도를 정량적으로 비교 평가하기 위하여 상기와 동일한 조건으로 배양된 세포들을 대상으로 DNA flow cytometry 분석을 통한 세포주기의 sub-G1기에 해당되는 세포들의 빈도를 조사하였다. Fig. 3B의 결과에서 알 수 있듯이, BJIKTP 처리 농도의 증가에 따라 sub-G1기에 해당하는 세 포의 빈도가 점차 증가되어 3 mg/ml 및 4 mg/ml BJIKTP 처리군에서 대조군에 비하여 약 2.9 및 3.7배로 높아졌다.
이상의 결과에서 BJIKTP 처리에 의한 A549 세포의 생존 율 감소는 apoptosis 유발과 직접적인 관련이 있음을 알 수 있었다.
4. Mitochondrial membrane potential (MMP)에 미치는 BJIKTP의 영향
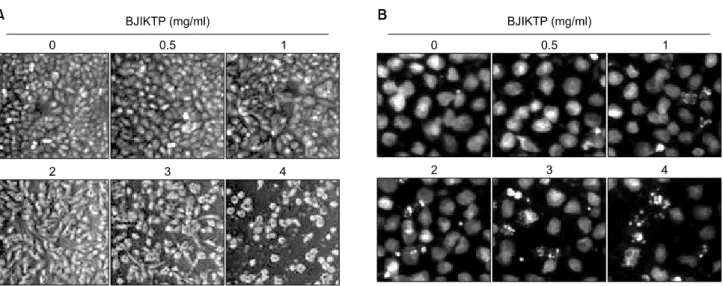
BJIKTP 처리에 따른 A549 세포의 apoptosis 유발에 관 한 기전 해석을 위하여 apoptosis 조절에 중심이 되는 mi- tochondria와의 연관성을 먼저 조사하였다. 이를 위하여 BJIKTP가 처리 A549 세포를 대상으로 MMP의 손실 정도 를 조사하기 위하여 dual-emission fluorescent dye인 JC-1을 적용시켜 형광반응에 따른 histogram을 분석하였다. Fig.
4의 결과에서 알 수 있듯이 BJIKTP 처리에 따른 MMP의 소실이 BJIKTP의 농도가 증가함에 점차 증가되어 최고 처리 농도인 4 mg/ml에서 약 50%의 세포에서 MMP 소실
Fig. 3. Induction of apoptosis by BJIKTP in A549 cells. (A) Cells were incubated with the indicated concentrations of BJIKTP for 48 h. For the analysis of DNA fragmentation, genomic DNA from cells was extracted, separated by 1.5%
agarose gel electrophoresis, and visualized under UV light after staining with EtBr. Marker indicates a size marker of the DNA ladder. The results shown are from one representative experiment of two experiments that showed similar patterns.
(B) To quantify the degree of apoptotic cells, cells were treated with the indicated concentrations of BJIKTP for 48 h. Then the cells were collected and stained with PI for flow cytometry analysis. The profile represents the increase of sub-G1 population and each point represents the mean of two independent experiments.
을 보여 주었다. 이는 BJIKTP에 의한 A549 세포의 apop- tosis 유도에 mitochondria의 기능 손상이 연관되어 있음을 의미하는 결과이다.
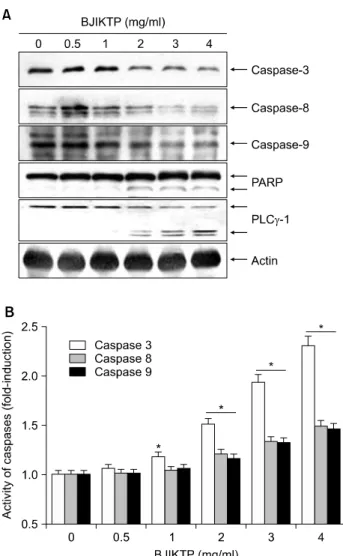
5. BJIKTP 처리에 의한 caspase의 활성 증가
다음은 BJIKTP 처리에 의한 apoptosis 유도가 extrinsic 및 intrinsic pathway 중 어느 경로를 활성화 시키는지를 조사하기 위하여 두 경로의 대표적인 initiator caspase에 해당되는 caspase-8 및 -9와, 그들에 의하여 활성이 증대 되는 caspase-3의 발현에 미치는 BJIKTP의 영향을 조사하 였다. Fig. 5A에 나타난 바와 같이 BJIKTP의 처리에 의하 여 initiator caspase에 해당되는 caspase-8과 -9 뿐만 아니라 effector caspase인 caspase-3의 비활성형 단백질의 발현이 BJIKTP 처리농도 의존적으로 현저히 감소되는 것으로 관찰되었으며, 활성형 caspase-3의 기질단백질에 해당되 는 poly (ADP-ribose) polymerase (PARP) 및 phospholipase C γ-1 (PLCγ-1)의 단편화 현상이 뚜렷하게 관찰되었다.
이상의 단백질 수준에서의 결과를 재확인하기 caspase의 활성 정도를 in vitro assay에 의하여 직접 분석한 결과, BJIKTP 처리 농도 증가에 따라 caspase-3의 활성이 현저 히 증가되었고 caspase-8 및 -9의 활성도 다소 증가되었음 을 알 수 있었다(Fig. 5B). 이상의 결과는 BJIKTP에 의한 A549 세포의 apoptosis 유도에는 extrinsic 및 intrinsic path- way가 동시에 관여할 가능성이 매우 높음을 의미한다.
6. Bcl-2 family의 발현에 미치는 BJIKTP의 영향
다음으로 Bcl-2 family에 속하는 인자들이 BJIKTP에 의 해 유발되는 apoptosis에 있어서 어떠한 역할을 하는지를 조사하였다. 이를 위하여 동일 조건에서 배양된 A549 세 포를 대상으로 몇 가지 주요 Bcl-2 family 인자들의 발현 변화를 전사 및 번역 수준에서 비교하였다. Fig. 6에 나타 낸 결과에서 알 수 있듯이, BJIKTP 처리에 의하여 대표 적임 pro-apoptotic 인자인 Bax의 발현이 mRNA 및 단백질 수분에서 모두 증가하였으며, anti-apoptotic 인자에 해당 되는 Bcl-2 및 Bcl-xL의 발현은 BJIKTP 처리 농도 증가에 따라 전사 및 번역 수준에서 현저하게 감소되었다. 이러 한 Bcl-2 family 인자들의 발현 변화는 MMP의 소실과 연 관된 intrinsic pathway의 활성이 BJIKTP에 의한 apoptosis 유도에 관여하였을 가능성이 높음을 보여주는 결과이 다. 아울러 Bid 단백질의 발현이 BJIKTP 처리에 따라 점 차 감소되어 truncated Bid (tBid)로의 전환이 증가하였을 것으로 추정된다. 이상의 결과는 BJIKTP 처리에 의한 pro-apoptotic Bax의 발현이 상대적으로 증가되어 MMP의 소실이 촉진되었을 것이며, 아울러 BJIKTP 처리에 의한
Fig. 4. Loss of MMP values by BJIKTP in A549 cells. (A) After incubation with various concentrations of BJIKTP for 48 h, cells were stained with JC-1 and then incubated at 37oC for 20 min, after which the mean JC-1 fluorescence intensity was detected with a flow cytometer. (B) Data represent the mean±SD of the representative experiments performed at least three times. The significance was determined by a Student’s t-test (*p<0.05 vs. untreated control).
caspase-8이 활성 증가가 tBid로의 전환을 증대시켜 ex- trinsic 및 intrinsic pathway의 연결을 통한 apoptosis 유도 증대를 가져왔을 것으로 추정된다.
7. IAP family의 발현에 미치는 BJIKTP의 영향
Inhibitors of apoptosis (IAP) family에 속하는 인자들은 caspase화의 직ㆍ간접적인 결합을 통하여 그들의 활성을 억제하는 역할을 하기 때문에 그들 인자들의 발현에 미 치는 BJIKTP의 영향을 RT-PCR 및 Western blotting 방법 으로 조사하였으며, 그 결과는 Fig. 7에 나타낸 바와 같 다. 제시된 결과에서 알 수 있듯이 A549 세포에서 BJIKTP 처리에 의하여 IAP family에 속하는 모든 인자들 의 발현이 전사 및 번역 수준에서 감소되었다. 따라서 BJIKTP 처리에 따른 IAP family 인자들의 발현 감소는 결 국 caspase의 활성 증대와 직접적인 연관성이 있음을 의 미한다.
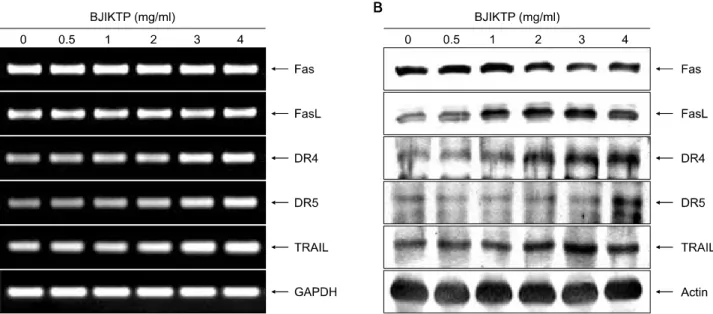
8. Death receptor 연관 인자들의 발현에 미치는 BJIKTP의 영향
BJIKTP 처리에 의한 A549 세포의 apoptosis 유발에 death receptor 관련 경로가 관여하는지를 확인하기 위하 여 RT-PCR 및 Western blot analysis 방법으로 주요 death receptor 연관 인자들의 발현 변화를 조사하였다. Fig. 8의 결과에서 알 수 있듯이 BJIKTP 처리에 의하여 death re- ceptor에 속하는 여러 유전자 중 death receptor 4 (DR4) 및 DR5가 전사와 번역수준에서 BJIKTP 처리 농도 의존적
으로 발현이 매우 증가하였으며, death ligand에 해당되는 tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) 및 Fas ligand (FasL)의 발현도 특히 전사 수준에서 크게 증가되었다. 이는 BJIKTP의 처리에 의한 A549 세 포에서의 apoptosis 유발에 extrinsic pathway가 개시 신호 로서 작용하였을 가능성이 매우 높음을 의미하는 결과 이다.
고 찰
본 연구에서는 보중익기가미길경탕(BJIKTP)의 항암활 성에 관한 효능을 인체폐암세포의 증식억제와 연관된 apoptosis 유발 관련 기전 측면에서 조사하였다. 이를 위 하여 A549 및 NCI-H460 폐암세포를 대상으로 BJIKTP의 항암활성 감수성을 조사한 결과, A549 세포에서 BJIKTP 에 대한 감수성이 상대적으로 높게 나타났다. 이러한 BJIKTP 처리에 따른 A549 세포의 증식억제가 apoptosis 유도 현상에 의한 것임을 전형적인 apoptosis가 유발된 세 포에서 관찰되는 3가지 주요 지표, 즉 염색질의 응축, DNA의 단편화 및 sub-G1 세포 빈도의 증가로 제시하였 다.
일반적으로 apoptosis 유발 경로는 intrinsic 및 extrinsic pathway로 두 가지로 대별되어지는데, extrinsic pathway의 경우 TRAIL 또는 FasL 등이 death receptor와 결합을 함으 로서 활성화된다. 그 중, Apo-2 ligand로 알려진 TRAIL은 정상세포에서는 세포독성을 나타내지 않지만,17,22) 암세
Fig. 5. Activation of caspases by BJIKTP in A549 cells. (A) Cells were treated with the indicated conditions of BJIKTP for 48 h. The cells were lysed, cellular proteins were separated by SDS-polyacrylamide gels and then transferred onto nitro- cellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (B) After 84 h incubation with the indicated concentrations of BJIKTP, the cells were lysed and aliquots (50μg protein) were assayed for in vitro caspase-3, -8, and -9 activity using DEVD-pNA, IETD-pNA, and LEHD-pNA as substrates, respec- tively, at 37oC for 1 h. The released fluorescent products were measured. Data are expressed as mean±SD of three inde- pendent experiments. Significance was determined using a Student’s t-test (*p<0.05 vs. untreated control).
포를 포함한 형질전환된 세포에서 두 가지 death do- main-containing agonistic receptor인 DR4 및 DR5와 결합함 으로서 caspase-8의 활성을 유도하는 apoptosis 개시 신호 를 자극한다.23,24) 그러나 TRAIL은 osteoprotegerin, DcR1 (TRAIL-R3) 및 DcR2 (TRAIL-R4)와 같은 세 종류의 decoy receptor와 결합을 하게 되면 TRAIL signaling이 억제된
다.24) 그리고 TNF receptor superfamily에 속하는 Fas (CD95/
Apo-1)에 Fas ligand (FasL/CD95L) 또는 agonistic anti-Fas an- tibody가 결합하면 Fas clustering, Fas-associated death do- main protein (FADD) 및 pro-caspase-8이 death-inducing sig- naling complex (DISC)를 형성하게 되어 caspase-8이 활성화 되고, 이에 의한 caspase cascade에 의하여 apoptosis를 유발 하게 된다.25,26) 즉 caspase-8은 death receptor 연관 apoptosis 신호 개시의 중요한 initiator caspase로 작용한다.27∼29) 본 연구의 결과에 의하면 A549 세포에서 BJIKTP의 처리에 의하여 FasL, DR4, DR5 및 TAIL을 포함한 extrinsic path- way 개시 신호에 관여하는 주요 death receptor 연관 인자 들의 발현이 매우 높게 나타났으며(Fig. 8), caspase-8의 활 성 또한 BJIKTP의 처리 농도 증가에 따라 서서히 증가되 었다(Fig. 5). 따라서 BJIKTP에 의한 A549 세포의 apopto- sis 유발에 extrinsic pathway의 활성화가 초기 apoptosis 유 도 신호로서 작용하였음을 의미한다.
Caspase는 apoptosis 유발에 핵심적으로 작용하는 효소군 으로서 핵과 mitochondria의 외막에 불활성 상태인 pro- enzyme 형태로 존재한다.30,31) 다양한caspases 중 extrinsic pathway의 initiator caspase의 하나인 caspase-8은 DISC에 의 해 활성화되어 effector caspase인 caspase-3 및 -7을 활성화 시킨다. 그리고 intrinsic pathway의 대표적인 initiator cas- pase인 caspase-9의 활성화는 mitochondria에서 유리된 cy- tochrome c에 의해서 형성된 apoptosome에 의하여 유발되 며, 활성화된 caspase-9는 caspase-3 및 -7을 또한 활성화시
킨다.21,32) 이렇게 활성화된 caspase-3은 세포 내 많은 표적
단백질들을 분해하면서 apoptosis를 유발한다. 따라서 다 양한 caspase의 활성화와 caspase-3 및 -7의 표적 단백질의 단편화는 apoptosis 유발에 대한 또 다른 증거가 될 수 있 다. Caspase-3 및 -7의 대표적인 표적 단백질 중 하나인 PARP는 DNA repair 동안 DNA의 결함을 인지하는 F1 및 F2라고 하는 두 개의 zinc finger motif를 가지고 있으며 molecular nick sensor 기능을 하는 N-terminal DNA binding domain과 NAD+가 결합하는 C-terminal catalytic domain으 로 구성되어 있다.33,34) PARP는 caspase-3 및 -7의 활성화 에 의하여 Asp214와 Gly215 사이에 분해가 일어남으로서 116 kDa의 분자량을 가진 PARP 단백질이 85 및 24 kDa 크기의 단편으로 잘리게 된다.35,36) 또 다른 이들 caspase 의 표적 단백질인 PLC-γ1은 조혈세포에만 존재하는 PLC-γ2와는 달리 체내 조직 모든 세포에 넓게 분포하고 있으며, phosphatidylinositol 4,5-bisphosphate (PI 4, 5-P2)를 protein kinase C (PKC) activator인 diacylglycerol 및 세포 내 Ca2+ 조절에 중요한 역할을 하는 inositol 1,4,5-trispho- sphate (IP3)로 가수분해된다.37,38) PLC-γ1 또한 대표적인
Fig. 6. Effects of BJIKTP on the Bcl-2 family members in A549 cells. (A) Cells were treated with the indicated concentrations of BJIKTP for 48 h. Total RNAs were isolated and reverse-transcribed. Resulting cDNAs were then subjected to PCR and the reaction products were subjected to electrophoresis in a 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
Fig. 7. Effects of BJIKTP on the IAP-2 family members in A549 cells. (A) Cells were treated with the indicated concentrations of BJIKTP for 48 h. Total RNAs were isolated and reverse-transcribed. Resulting cDNAs were then subjected to PCR and the reaction products were subjected to electrophoresis in a 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
caspase-3의 기질 단백질 중의 하나이므로, PARP와 함께 caspase 활성 의존적 apoptosis 유발의 생화학적 표식자로 널리 사용이 되고 있다.39,40) 본 연구의 결과에 의하면 BJIKTP가 처리된 A549 세포에서 이들 두 표적 단백질의 분해가 BJIKTP 처리 농도의 증가에 따라 매우 증가되어 BJIKTP에 의한 apoptosis 유발은 casapse의 활성화가 중요 한 역할을 하고 있음을 알 수 있다(Fig. 5).
Caspases 의존적인 apoptosis는 여러 단계로 조절되는데 그 중 IAP family에 속하는 몇몇 단백질들은 caspase에 직ㆍ 간접적으로 작용하여 그들 효소 활성을 억제하는 것으 로 알려져 있다.41∼43) 현재까지 밝혀진 여덟 종류의 hu- man IAPs는 한 개 이상의 baculovirus IAP repeat (BIR) do- main을 가진다. Human IAPs 중 일부는 caspase의 ubiquiti- nation 및 degradation을 조절하는 RING finger domain을 가
Fig. 8. Effects of BJIKTP on the death receptor-related members in A549 cells. (A) Cells were treated with the indicated concentrations of BJIKTP for 48 h. Total RNAs were isolated and reverse-transcribed. Resulting cDNAs were then subjected to PCR and the reaction products were subjected to electrophoresis in a 1% agarose gel and visualized by EtBr staining. GAPDH was used as an internal control. (B) The cells were lysed and then cellular proteins were separated by SDS-polyacrylamide gels and transferred onto nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control.
지며, 또 다른 일부는 protein–protein interaction 기능을 하 는 caspase-recruitment domain (CARD)을 가지는 것으로 알 려져 있다.44) IAPs의 중요한 기능은 BIR domain 의존적인 상호작용 및 caspase-3, -7 및 -9의 억제를 통한 apoptosis의 억제이다. 가장 강력한 IAP로 알려진 X-linked IAP (XIAP) 는 활성화된 caspase와 높은 친화력을 가지며 apoptosis를 억제한다. XIAP의 BIR3 domain은 caspase-9와 결합하는 반면에 BIR1 및 BIR2 domain은 활성형 caspase-3 및 -7과 결합하여 apoptosis를 억제한다.45∼48) XIAP 외에도 cIAP-1 및 cIAP-2 등도 caspase와 결합하여 apoptosis를 억제하는 것으로 알려져 있는데,41,49) IAPs의 발현은 전사 조절인자 인 nuclear factor kappa B (NF-κB)에 의해 조절 받는 것으 로 밝혀져 apoptosis의 신호전달 연구에서 있어서 매우 중
요하다.50,51) 본 실험의 결과에 의하면 BJIKTP가 함유된
배지에서 배양된 A549 세포에서는 조사된 모든 IAP fam- ily 인자들의 발현이 매우 저하되었으며(Fig. 7), 이러한 IAP family 인자들의 발현 감소는 BJIKTP 처리에 의한 caspase의 활성화에 기여하였음을 알 수 있었다.
한편 mitochondria를 중심으로 일어나는 intrinsic path- way의 활성화에는 Bcl-2 family 인자들의 발현 변화가 중 요한 역할을 하는데, Bcl-2 family에 속하는 단백질들은 네 가지의 Bcl homology (BH) domains (BH1–BH4) 중 최소 한 한 개의 domain을 포함하고 있으며, mitochondria 보존
과 mitochondria에 의해 유도되는 apoptosis를 조절하는 중 요한 조절자이다. 이 단백질들은 Bcl-2, Bcl-xL, Bcl-w 및 Mcl-1과 같이 apoptosis를 억제하는 anti-apoptotic member 와 Bax, Bad, Bak, Bid 및 Bcl-xS와 같이 apoptosis를 유발하 는 pro-apoptotic member로 구성되어 있다.52,53) 결국 이러 한 anti-apoptotic 및 pro-apoptotic member들 사이의 상대적 발현 변화의 차이가 특정 생리적 및 병리적인 조건 하에 서 세포의 생존 여부를 결정하게 된다.54) 즉 pro-apoptotic member의 발현 증가로 이 member들 간의 균형이 바뀐다 면 mitochondria로부터 cytosol로 cytochrome c가 방출되어 caspases, 종양억제 유전자인 p53, DNA의 단편화와 연관 된 endonuclease 등의 활성이 증가되어 apoptosis가 유발되 는 것으로 알려져 있다.55,56) 따라서 BJIKTP 처리에 따른 apoptosis 유발에 intrinsic pathway가 연관되어 있는지의 여 부를 조사한 결과, BJIKTP가 처리된 A549 세포에서 BJIKTP 처리 농도 증가에 따라 MMP의 소실이 급속히 증가되었다(Fig. 4). 아울러 Bcl-2 family에 속하는 인자들 중, pro-apoptotic Bax의 발현이 BJIKTP 처리에 의하여 매 우 증가되었으며, anti-apoptotic Bcl-2 및 Bcl-xL의 발현은 현저히 감소되었다(Fig. 6). 이러한 결과는 BJIKTP에 의 한 A549 세포의 apoptosis 유도에 intrinsic pathway의 활성 화가 관여하고 있음을 의미한다. 아울러 Bcl-2 family에 속하는 또 다른 인자인 Bid의 발현이 BJIKTP가 함유된
배지에서 배양된 A594 세포에서 매우 감소되었는데, Bid 의 발현이 감소되었다는 것은 tBid로의 전환이 많이 되 었음을 의미하는 것으로 BJIKTP 처리에 따라 Bid의 trun- cation의 비율이 증대되었을 것으로 추정된다(Fig. 6). Bid 의 tBid로의 전환은 활성화된 caspase-8에 의한 것이며, tBid는 mitochondria로 이동되어 intrinsic pathway를 증폭시 키는 작용에 관여한다.52,53) 따라서 A549 세포에서 BJI- KTP의 처리에 의하여 pro-apoptotic Bax의 발현이 상대적 으로 증가되어 MMP의 소실이 촉진되었을 것이며, 아울 러 BJIKTP 처리에 의한 caspase-8이 활성 증가가 tBid로의 전환을 증대시켜 extrinsic 및 intrinsic pathway의 연결을 통 한 apoptosis 유도 증대를 가져왔을 것으로 추정된다.
결 론
이상의 결과를 종합하여 보면, BJIKTP에 의한 A549 폐암세포의 증식억제는 apoptosis 유발과 밀접한 관련이 있었으며, 이러한 apoptosis의 유도에는 intrinsic 및 ex- trinsic pathway가 동시에 관여하며, 특히 Bid의 tBid로의 전환이 두 경로를 연결하는 매개체 역할을 하고 있음을 알 수 있었다. 이러한 연구 결과는 BJIKTP의 생화학적 항암기전 해석을 이해하고 향후 지속적인 연구를 위한 귀중한 자료로 사용될 것이라 생각된다.
감사의 글
본 논문은 2011년도 동의대학교 교내연구비(2011- AA113) 지원으로 이루어졌음.
참 고 문 헌
1) 황도연. 증맥 방약합편. 서울, 남산당, pp 142-145, 1989.
2) 허 준. 동의보감 잡병편. 서울, 여강출판사, pp 1561- 1562, 1994.
3) 전국한의과대학 방제학 교수 공편저. 방제학. 서울, 영 림사, pp 168-169, 1994.
4) BaeJ H, Shin DY, Park C, Park SE, Hong SH, Park DI, Choi YH. Inhibition of cell invasion by Bojungikgigilkyung-tang in human lung cancer A549 cells. Cancer Prev Res 16, 238-248, 2011.
5) 전국한의과대학 본초학 교수 공편저. 본초학. 서울, 영 림사, pp 460-461, 1994.
6) 황도연. 증맥 방약합편. 서울, 남산당, pp 124, 1989.
7) 허 준. 동의보감 탕약 침구편. 서울, 여강출판사, pp 2704, 1994.
8) Weinberg J. Oncogenes and tumor suppressor genes. CA
Cancer J Clin 44, 160-170, 1994.
9) Leist M, Jäättelä M. Four deaths and a funeral: from caspases to alternative mechanisms. Nat Rev Mol Cell Biol 2, 589-598, 2001.
10) Vilches Troya J. Understanding cell death: a challenge for biomedicine. An R Acad Nac Med 122, 631-656, 2005.
11) Huerta S, Goulet EJ, Huerta-Yepez S, Livingston EH.
Screening and detection of apoptosis. J Surg Res 139, 143-156, 2007.
12) Huerta S, Goulet EJ, Livingston EH. Colon cancer and apoptosis. Am J Surg 191, 517-526, 2006.
13) Los M, Stroh C, Jänicke RU, Engels IH, Schulze-Osthoff K.
Caspases: more than just killers? Trends Immunol 22, 31-34, 2001.
14) Schultz DR, Harrington WJ Jr. Apoptosis: programmed cell death at a molecular level. Semin Arthritis Rheum 32, 345-369, 2003.
15) Zamzami N, Susin SA, Marchetti P, Hirsch T, Gómez- Monterrey I, Castedo M, Kroemer G. Mitochondrial control of nuclear apoptosis. J Exp Med 183, 1533-1544, 1996.
16) Kischkel FC, Lawrence DA, Chuntharapai A, Schow P, Kim KJ, Ashkenazi A. Apo2L/TRAIL-dependent recruitment of endogenous FADD and caspase-8 to death receptors 4 and 5. Immunity 12, 611-620, 2000.
17) Walczak H, Krammer PH. The CD95 (APO-1/Fas) and the TRAIL (APO-2L) apoptosis systems. Exp Cell Res 256, 58-66, 2000.
18) Susin SA, Zamzami N, Castedo M, Daugas E, Wang HG, Geley S, Fassy F, Reed, JC, Kroemer G. The central execu- tioner of apoptosis: multiple connections between protease activation and mitochondria in Fas/APO-1/CD95- and cer- amide-induced apoptosis. J Exp Med 186, 25-37, 1997.
19) Baliga B, Kumar S. Apaf-1/cytochrome c apoptosome: an es- sential initiator of caspase activation or just a sideshow? Cell Death Differ 10, 16-18, 2003.
20) Liu X, Kim CN, Yang J, Jemmerson R, Wang X. Induction of apoptotic program in cell-free extracts: requirement for dATP and cytochrome c. Cell 86, 147-157, 1996.
21) Green DR, Reed JC. Mitochondra and apoptosis. Science 281, 1309-1312, 1998.
22) Belka C, Jendrossek V, Pruschy M, Vink S, Verheij M, Budach W. Apoptosis-modulating agents in combination with radiotherapy-current status and outlook. Int J Radiat Oncol Biol Phys 58, 542-554, 2004.
23) Wang S, El-Deiry WS. TRAIL and apoptosis induction by TNF-family death receptors. Oncogene 22, 8628-8633, 2003.
24) Kimberley FC, Screaton GR. Following a TRAIL: update on a ligand and its five receptors. Cell Res 14, 359-372, 2004.
25) Kischkel FC, Hellbardt S, Behrmann I, Germer M, Pawlita M, Krammer PH, Peter ME. Cytotoxicity-dependent APO-1 (Fas/CD95)-associated proteins form a death-inducing sig- naling complex (DISC) with the receptor. EMBO J 14, 5579-5588, 1995.
26) Ashkenazi A. Targeting death and decoy receptors of the tumour-necrosis factor superfamily. Nat Rev Cancer 2, 420- 430, 2002.
27) Budihardjo I, Oliver H, Lutter M, Luo X, Wang X. Bio- chemical pathways of caspase activation during apoptosis.
Annu Rev Cell Dev Biol 15, 269-290, 1999.
28) Peták I, Houghton JA. Shared pathways: death receptors and cytotoxic drugs in cancer therapy. Pathol Oncol Res 7, 95-106, 2001.
29) Chen M, Wan J. Initiator caspases in apoptosis signaling pathways. Apoptosis 7, 313-319, 2002.
30) Nagata S. Apoptosis by death factor. Cell 88, 355-365, 1997.
31) Allen RT, Cluck MW, Agrawal DK. Mechanisms controlling cellular suicide: role of Bcl-2 and caspases. Cell Mol Life Sci 54, 427-445, 1998.
32) Chai F, Truong-Tran AQ, Ho LH, Zalewski PD. Regulation of caspase activation and apoptosis by cellular zinc fluxes and zinc deprivation: A review. Immunol Cell Biol 77, 272-278, 1999.
33) de Murcia G, Ménissier de Murcia J. Poly (ADP-ribose) poly- merase: a molecular nick-sensor. Trends Biochem Sci 19, 172-176, 1994.
34) Muller S, Briand JP, Barakat S, Lagueux J, Poirier GG, De Murcia G, Isenberg DA. Autoantibodies reacting with poly (ADP-ribose) and with a zinc-finger functional domain of poly (ADP-ribose) polymerase involved in the recognition of damaged DNA. Clin Immunol Immunopathol 73, 187-196, 1994.
35) Lazebnik YA, Kaufmann SH, Desnoyers S, Poirier GG, Earnshaw WC. Cleavage of poly (ADP-ribose) polymerase by a proteinase with properties like ICE. Nature 371, 346-347, 1994.
36) Kaufmann SH, Desnoyers S, Ottaviano Y, Davidson NE, Poirier GG. Specific proteolytic cleavage of poly (ADP-ribose) polymerase: an early marker of chemotherapy-induced apop- tosis. Cancer Res 53, 3976-3985, 1993.
37) Nishizuka Y, Kikkawa U. Early studies of protein kinase C:
a historical perspective. Methods Mol Biol 233, 9-18, 2003.
38) Berridge MJ, Irvine RF. Inositol trisphosphate, a novel second messenger in cellular signal transduction. Nature 312, 315- 321, 1984.
39) Myklebust JH, Blomhoff HK, Rusten LS, Stokke T, Smeland EB. Activation of phosphatidylinositol 3-kinase is important for erythropoietin-induced erythropoiesis from CD34(+) he- matopoietic progenitor cells. Exp Hematol 30, 990-1000, 2002.
40) Bae SS, Perry DK, Oh YS, Choi JH, Galadari SH, Ghayur T, Ryu SH, Hannun YA, Suh PG. Proteolytic cleavage of phospholipase C-gamma1 during apoptosis in Molt-4 cells.
FASEB J 14, 1083-1092, 2000.
41) Deveraux QL, Reed JC. IAP family proteins-suppressors of apoptosis. Genes Dev 13, 239-252, 1999.
42) Deveraux QL, Roy N, Stennicke HR, Van Arsdale T, Zhou Q, Srinivasula SM, Alnemri ES, Salvesen GS, Reed JC. IAPs block apoptotic events induced by caspase-8 and cytochrome c by direct inhibition of distinct caspases. EMBO J 17, 2215- 2223, 1998.
43) Salvesen GS, Duckett CS. IAP proteins: blocking the road to death's door. Nat Rev Mol Cell Biol 3, 401-410, 2002.
44) Checinska A, Hoogeland BS, Rodriguez JA, Giaccone G, Kruyt FA. Role of XIAP in inhibiting cisplatin-induced caspase activation in non-small cell lung cancer cells: a small molecule Smac mimic sensitizes for chemotherapy-induced apoptosis by enhancing caspase-3 activation. Exp Cell Res 313, 1215-1224, 2007.
45) Scott FL, Denault JB, Riedl SJ, Shin H, Renatus M, Salvesen GS. XIAP inhibits caspase-3 and -7 using two binding sites:
evolutionarily conserved mechanism of IAPs. EMBO J 24, 645-655, 2005.
46) Shiozaki EN, Chai J, Rigotti DJ, Riedl SJ, Li P, Srinivasula SM, Alnemri ES, Fairman, R, Shi Y. Mechanism of XIAP- mediated inhibition of caspase-9. Mol Cell 11, 519-527, 2003.
47) Chai J, Shiozaki E, Srinivasula SM, Wu Q, Datta P, Alnemri ES, Shi Y. Structural basis of caspase-7 inhibition by XIAP.
Cell 104, 769-780, 2001.
48) Huang Y, Park YC, Rich RL, Segal D, Myszka DG, Wu H.
Structural basis of caspase inhibition by XIAP: differential roles of the linker versus the BIR domain. Cell 104, 781-790, 2001.
49) Roy N, Deveraux QL, Takahashi R, Salvesen GS, Reed JC.
The c-IAP-1 and c-IAP-2 proteins are direct inhibitors of specific caspases. EMBO J 16, 6914-6925, 1997.
50) Cheng JQ, Jiang X, Fraser M, Li M, Dan HC, Sun M, Tsang BK. Role of X-linked inhibitor of apoptosis protein in che- moresistance in ovarian cancer: possible involvement of the phosphoinositide-3 kinase/Akt pathway. Drug Resist Updat 5, 131-146, 2002.
51) LaCasse EC, Baird S, Korneluk RG, MacKenzie AE. The inhibitors of apoptosis (IAPs) and their emerging role in cancer. Oncogene 17, 3247-3259, 1998.
52) Reed JC. Bcl-2 family proteins. Oncogene 17, 3225-3236, 1998.
53) Du J, Chen GG, Vlantis AC, Chan PK, Tsang RK, van Hasselt CA. Resistance to apoptosis of HPV 16-infected laryngeal cancer cells is associated with decreased Bak and increased Bcl-2 expression. Cancer Lett 205, 85-88, 2004.
54) Kroemer G. The proto-oncogene Bcl-2 and its role in regulating apoptosis. Nat Med 3, 614-620, 1997.
55) Donovan M, Cotter TG. Control of mitochondrial integrity by Bcl-2 family members and caspase-independent cell death.
Biochim Biophys Acta 1644, 133-147, 2004.
56) Rosse T, Olivier R, Monney L, Rager M, Conus S, Fellay I, Jansen B, Borner C. Bcl-2 prolongs cell survival after Bax- induced release of cytochrome c. Nature 391, 496-499, 1998.