C
ANCERPREVENTION RESEARCH □ REVIEW ARTICLE □
163 책임저자:조쌍구, 143-701, 서울시 광진구 화양동 1
건국대학교 동물생명공학과 Tel: 02-450-4207, Fax: 02-455-1044 E-mail: ssangoo@konkuk.ac.kr
접수일:2007년 7월 30일, 게재승인일:2007년 9월 10일
Correspondence to:Ssang-Goo Cho
Departmeny of Animal Biotechnology and RCTCP, Konkuk University, 1, Hwayang-dong, Gwangjin-gu, Seoul 143-701, Korea
Tel: +82-2-450-4207, Fax: +82-2-455-1044 E-mail: ssangoo@konkuk.ac.kr
플라보노이드의 세포 신호전달 조절
건국대학교 동물생명공학과
이응룡ㆍ강근호ㆍ강용진ㆍ김우열ㆍ최혜연ㆍ김봉우ㆍ정효순ㆍ조쌍구
Regulation of Cellular Signal Transduction by Flavonoids
Eung-Ryoung Lee, Geun-Ho Kang, Yong-Jin Kang, Woo-Yeul Kim, Hye-Yeon Choi, Bong-Woo Kim, Hyo-Soon Jeong and Ssang-Goo Cho
Department of Animal Biotechnology and RCTCP, Konkuk Unitersity, Seoul 143-701, Korea
Many studies revealed the neuroprotective, cardioprotective, and chemopreventive actions of dietary flavonoids. The plausible mechanistic interpretation of the various effects of flavonoids was concentrated on the anti-oxidant or free radical-scavenging properties of these phytochemicals, both in model systems and under in vivo conditions. While there has been a major focus on the anti-oxidant properties, there is an emerging view that flavonoids and their in vivo metabolites, do not act as conventional hydrogen-donating anti-oxidants, but they may exert regulatory functions in cells through actions at protein kinase or lipid kinase signaling pathways. Flavonoids and more recently their metabolites, have been reported to act at phosphoinositide 3-kinase (PI 3-kinase), Akt/protein kinase B (Akt/PKB), protein kinase C (PKC), mitogen activated protein kinase (MAP kinase), and various tyrosine kinases signaling cascades.
Inhibitory or stimulatory actions at these pathways are likely to affect cellular function profoundly by altering the phosphorylation state of target molecules and by modulating gene expression. A clear understanding of the mechanisms of action of flavonoids, either as anti-oxidants or modulators of cellular signaling pathways, and the influence of their metabolism on these properties are key to the evaluation of these potent biomolecules as anti-cancer agents, cardio-protectants, and inhibitors of neurodegeneration.
(Cancer Prev Res 12, 163-173, 2007)
Key Words: Flavonoid, Anti-oxidant, MAP kinase, PI3-Kinase, Apoptosis, Flavonoid metabolite, Free radicals
서 론
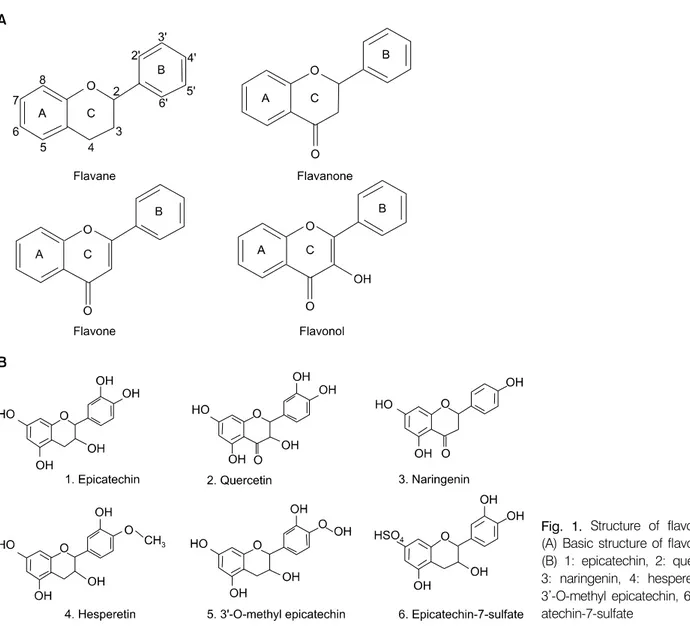
플라보노이드는 대부분 phenylalanine에서 합성되어지 는 광범위한 식물 색소의 구성 물질로, 최근 의학적 치료 의 목적으로 그 구조와 기능에 대한 많은 연구가 이루어 지고 있다. 산소 하나를 함유하는 Doublic Ring Compound 인 Benzopyran에 Phenyle과 산소 하나가 더 첨가되어 Ketons (C=O)를 형성하면 Flavone 구조를 이루게 된다.
여기에 다른 Function기가 결합된 것을 총체적으로 플라
보노이드(Flavonoid)라고 한다. 플라보노이드의 기본 구 조는 Fig. 1A에서 보는 바와 같다. 플라보노이드는 자연 계에서 식물의 색소에 많이 포함되어 있으며, 대부분은 UV light에 의해서 자극되어 밝은 형광을 나타낸다. 플라 보노이드는 일반적으로 식물에서 당과 결합된 형태로 존재하며, 잎, 꽃, 과실에서 orange, scarlet, blue색을 띄게 하는데 관여하며 다양한 과실과 야채 이외에도 wine (particularly red wine), tea, beer와 같은 음료와 씨앗, 땅콩, 곡류, 향신료, 약용 식물에서 발견된다. 플라보노이드는 암, 심장혈관 질환, 그리고 신경퇴화 방지에 효과가 있다
Fig. 1. Structure of flavonoids:
(A) Basic structure of flavonoids, (B) 1: epicatechin, 2: quercetin, 3: naringenin, 4: hesperetin, 5:
3’-O-methyl epicatechin, 6: epic- atechin-7-sulfate
고 보고되고 있는데, 이 물질의 생물학적 작용 중에 항산 화 작용이 중요하다고 알려져 있다. 하지만, 플라보노이 드가 나타내는 효과에 대한 인체 내에서의 작용 기작에 대한 정확한 분자 기전은 명백하게 밝혀져 있지 않은 실 정이다. 최근 플라보노이드가 양성자를 공여하여 잔재 산소를 환원시켜 항산화 효과를 나타낸다고 보고되고 있다.1) 이러한 보고들은 플라보노이드가 산화 환원 반응 에 중요한 변화들을 일으키며 체내에서 광범위하게 대 사 작용에 관여 한다는 것을 보여주고 있다. 생체 내에서 의 플라보노이드의 반응 형태들은 식물체 내에서 와는 다른 반응인 것으로 알려져 있는데, 예를 들어 신체 내 소장 흡수를 기반으로 한 플라보노이드의 반응 형태들 로부터 발생하는 대사 작용과 반응은 배당체나 배당체 의 가수 분해로 얻어지는 당 이외의 성분들에서의 식물 체 반응 형태에서는 찾아볼 수 없다고 알려져 있다.
Quercetin, hesperetin, naringenin, epicatechin 등과 같은 일반 적인 플라보노이드의 구조는 Fig. 1B에서 보는 바와 같 다. 대장 안에서 일어나는 현상에 대한 새로운 이론도 보고되고 있는데 대장 속에 존재하는 소화관 미생물의 효소가 플라보노이드를 simple phenolic acid (간에서 흡수 된 후 물질대사로 변화되어 작용하는 것)로 분해된다는 보고가 있다.2,3)
플라보노이드가 생체 내 대사를 통해 흡수되는 것은 세포의 특이성에 의존한다고 알려져 있는데,4) 이는 플라 보노이드가 단순하게 확산을 통해 세포 속으로 수동적 으로 흡수되기 보다는 많은 세포 간 대사 작용의 레벨들 과 빠른 전달 비율에 따라 특이적으로 흡수가 일어난 다 는 것이다. 일반적으로 플라보노이드들은 3가지 단계의 대사 작용을 거친다고 알려져 있다. 첫 번째로 글루타티 온(glutathione (GSH))과 결합을 형성하는 대사 작용을 거
이응룡 외 7인:Regulation of Cellular Signal Transduction by Flavonoids 165
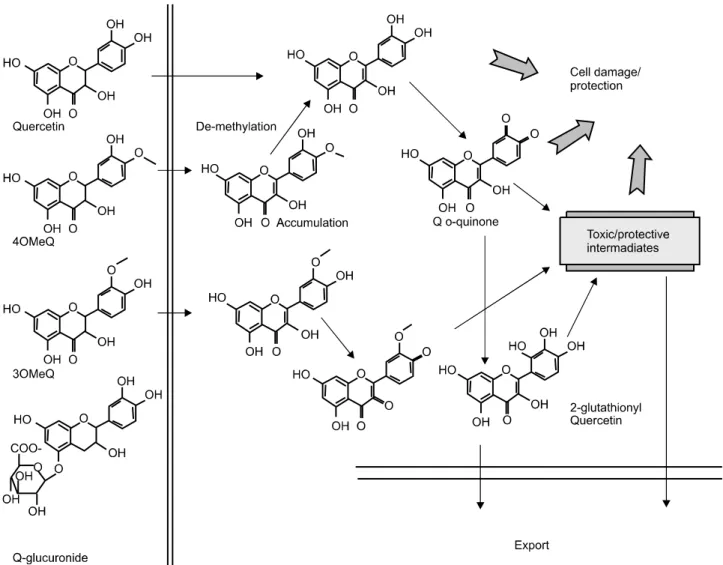
Fig. 2. Summary of the cellular metabolism of quercetin and its O-methylated metabolites. O-Methylated forms of quercetin associate with fibroblasts to a greater extent than quercetin, whereas the glucuronide does not enter. Quercetin and 3'-O-methyl quercetin undergo oxidative metabolism within the cells to yield as yet unidentified intermediates and a small amount of 4'-O-methyl quercetin is demethylated. Quercetin O-quinone reacts with GSH to form a 2'-glutathionyl conjugate. Both the GSH conjugate and oxidative products are exported from cells.15)
치고, 두 번째로 산화적 대사 작용, 그리고 세 번째는 P450과 관련된 대사 작용이다. 예를 들어 인간의 피부 섬유 세포 안에서 quercetin의 세포 상호적 대사 작용은 세포 간 산화적 생성물인 2’-glutathionyl quercetin (Fig. 2) 과 quercetin의 O-메틸화 형태의 de-methylation의 형성에 영향을 끼친다고 보고되었다. 반면에 epicatechin과 그것 의 O-메틸화 형태 대사물질은 세포 대사 작용에 크게 관 여 하지 않는다고 보고되고 있다.5) 2’-glutathionyl quer- cetin과 같은 플라보노이드에 의해 생성된 세포 대사 물 질들은 세포 안에서 유익하게 작용하거나 혹은 독성 작 용을 나타내는 것으로 연구되고 있으며, 플라보노이드 의 대사조정은 라디칼을 안정화시키기 위해서 hydrogen 을 줄 수 있는 B-ring catechol group (dihydroxylated B-ring)
의 존재로 나타나는 항산화 특성을 변화시킨다고 보고 되고 있다.6) 항산화 성질에서 중요한 구조적 특성은 C-ring에서 4-oxy-와 철이나 구리 같은 전이 금속 이온들 을 묶을 수 있는 기능적 능력이 있는 group들을 함유하 는 결합 내에서 2, 3번 위치에 불포화 화합물을 합성한다 고 보고되었다(Fig. 3, Quercetin의 예). Glucuronide와 O-메 틸화 형태, 그리고 플라보노이드-GSH 부가생성물(세포 간 대사물질) 같은 순환적 플라보노이드 대사물질들은 수소를 주는 능력이 감소된 특징을 가지고 있으며 또한 질소 생성물과 활성화 산소(reactive oxygen)를 효과적으 로 처리하기도 한다.7) 뇌 기관이나 원형질 내부 등의 생 체 내에서의 플라보노이드의 농도와 생체 내 축적된 그 들의 대사물질의 형태는8) ascorbic acid와 a-tocopherol 같은
Fig. 3. Flavonol: quercetin. Structure of a flavonol, quercetin shows features important in defining the classical antioxidant potential of flavonoids. Its most important feature is the catechol or dihydroxylated B-ring. Other important features include the presence of unsaturation in the C-ring and the presence of a 4-oxo function in the C-ring.
항산화 영양소 분자보다 낮은 단계의 대사물질 형태를 가지는 것으로 보고되었다. 플라보노이드는 유전자 발 현 억제뿐만 아니라 auxin indolyl acetic acid의 exocytosis를 통하여 식물성장의 많은 조절 기능에 영향을 끼치고 여 러 생물학적 기능을 조절한다. 그리고 플라보노이드는 reverse transcriptase와 protease와 같은 viral enzyme은 물론 이고 bacterial strain을 줄이거나 죽이는 기능도 한다. 이 것은 또한 특정 병원성 원생동물들을 죽인다고도 알려 져 있지만, 동물 세포에 대해 독성은 매우 낮다고 알려져 있다. 플라보노이드는 그들의 특정 효소를 특이적으로 저해하는 성질로 인하여 많은 질병의 치료에서 특정 호 르몬이나 신경전달물질을 자극시키거나 free radical을 제 거하는데 이용되어져 오고 있다.
플라보노이드의 작용: 항산화
플라보노이드는 활성화 산소종(reactive oxygen species (ROS))과 활성화 질소종(reactive nitrogen species (RNS))을 제거하는 항산화제로써 작용한다고 알려져 있다. 플라 보노이드의 기능에 대한 많은 연구에도 불구하고 플라 보노이드의 pro-oxidant/anti-oxidant 성질은 논쟁으로 남아 있으며, 또한 이들 효과에 대한 자세한 molecular mecha- nism은 잘 알려지지 않고 있다. 플라보노이드의 고리 구 조물에 부착되어 있는 phenolic hydroxyl group의 숫자와 배열에 따라 항산화제로써의 작용이 다르며 산화물에 전자를 공여함으로써 항산화 작용을 하는 플라보노이드 의 기능은 그들 스스로가 가지고 있는 라디칼의 환원 전 위와 라디칼에 대한 플라보노이드의 접근 가능성에 따 라 이뤄진다. 예를 들어 플라보노이드의 프록실기 라디 칼 제거과정에서 플라보노이드는 알킬 페록실기 라디칼
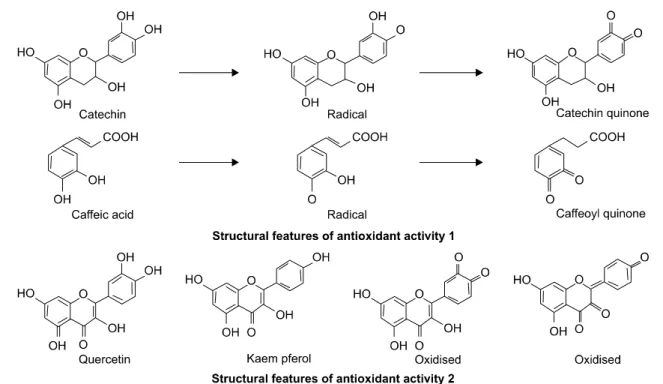
에 비례하여 알맞은 환원 전위를 유도할 수 있으며, 또한 플라보노이드는 산화 반응의 효과적인 억제인자가 될 수 있다. 플라보노이드의 알킬 페록실기 자유 라디칼을 감소시키는 특성의 기초가 되는 주된 구조적 특징은 B- 고리 형태 안에 존재하는 catechol (3’,4’-dihydroxy)이며, catechin과 caffeic acid의 환원 전위와 매우 비슷하다. 구조 적 특징을 좀 더 살펴보면 페닐기 링에 결합체가 연결되 고 플라보노이드의 라디칼 환원 전위는 낮은 환원 전위 의 B-고리 형태와 일치됨을 알 수 있다. 플라보노이드 구조 속에 페놀의 배열과 구조적 특성에 따른 항산화 작 용을 Fig. 4에서 나타냈다.
저밀도 리포 단백질의 산화가 동맥 경화 질병을 유발 한다는 사실을 알아낸 시점부터9) 많은 연구들은 저밀도 리포 단백질의 산화와 지질의 산화 반응을 막는 플라보 노이드의 성질을 감소시키는 것이 페록실기 라디칼이라 는 것이 연구 되었다.10) 산화반응에 대한 저밀도 리포 단 백질의 민감성을 감소시키는 플라보노이드의 기능에 대 해 활발한 실험이 이뤄지고 있다.
조직의 염증부위에서 iNOS 단백질이 생성되고 이러 한 NO 합성 효소(nitric oxide synthase)에 의한 nitric oxide (NO)의 합성에 대한 연구가 활발히 되고 있다. 플라보노 이드와 phenolic acid가 RNS를 제거하는 능력을 가지고 있 다는 것에 초점을 맞춘 몇몇의 연구들이 있다. RNS는 혈 관 내피 세포와 뉴런 안에서 nitric oxide synthase (NOS)의 작용에 의해 생성된다. 만성적인 염증 조직에서 동시에 발생하는 NO와 superoxide 라디칼의 생성물은 과산화아 질산염을 생성한다. 과산화아질산염은 유독한 병을 일 으키는 성분이며 superoxide 라디칼과 NO의 빠른 상호작 용에 의해 생성되는 nitrating species에 의한 것으로 보고 되었는데, peroxynitrous acid를 형성하는 pH에서 양자를 더해 생성 된다. 특히 페놀 성분은 조직에서 peroxynitrite- dependent reaction의 마커로써 요구되어 지는 3-nitrotyro- sine을 형성하며 peroxynitrite-dependent reaction에 적합한 성질을 갖는다고 보고되었다.11) 예를 들면 3-nitrotyrosine 는 증가한 NO 생성물과 깊은 연관을 가지고 있는 몇몇 질병에서 발견되고 있다. 하지만 nitrotyrosine이 이러한 질병들을 유발하는 인자인지는 확실치 않다. Nitrated protein은 면역성을 가지고 있으며, nitration은 그들 스스 로 자신의 기능과 안정성을 바꿀 수 있다고 알려져 있으 며, 단백질의 peroxynitrite-dependent nitration을 억제하는 능력은 RNS에 의해 생성되는 생분자의 해로움을 감소시 키는데 도움을 준다. 이러한 점에서 플라보노이드는 유 해 물질 억제 요소로 작용할 수 있는데, 몇몇 연구들에 의하면 Tyrosine의 peroxynitrite-dependent-nitration을 억제
이응룡 외 7인:Regulation of Cellular Signal Transduction by Flavonoids 167
Fig. 4. Oxidization of flavonoids. Oxidation of catechin, caffeic acid, quercetin, and kaempferol.
하는 플라보노이드의 작용은 그들의 구조에 의해 결정 된다고 조사되었다. 그런 이유에서 페놀기를 포함하고 있는 catechol은 전자의 제공을 통해 peroxynitrite를 제거 하고12) monophenolic B 고리 구조를 가지고 있는 mono- hydroxycinnamate와 플라보노이드는 tyrosine과 peroxynit- rite의 상호작용에 간섭함으로써 작용한다는 보고가 있 다. 몇몇 플라보노이드와 페놀 성분은 체내에서 nitrous acid-dependent nitration, DNA de-amination을 강하게 억제 할 수 있다고 보고되었다. 따라서 플라보노이드 성분을 가지고 있는 음식물들은 높은 수위의 RNS가 생성되는 corditois 상태에 위를 보호하는 효과를 제공한다고 알려 져 있으며, 하이드록실기를 가지고 있는 플라보노이드 와 페놀 성분은 chelate를 형성하는 transition metal ion과 상호작용할 수 있다고 보고되어지고 있다.13) 또한 많은 환경 오염물질에 의한 대기권 파괴는 유해한 자외선의 지표면 도달을 야기 시키고 있다. 이러한 자외선에 의해 피부세포는 많은 손상을 받고 있으며 이러한 자외선은 세포내에서 활성 산소종을 생성시키고 활성산소종에 의 해 피부세포의 세포사멸을 유도한다고 알려져 있다. 활 성산소종에 의한 피부손상을 플라보노이드의 한 종류인 eriodictyol이 활성산소를 저해시켜 자외선에 의한 피부세 포의 세포사멸을 저해한다는 보고가 되고 있다.14) 이러 한 플라보노이드의 항산화 기작은 생체 내에서 여러 가 지 자극에 의한 활성산소종을 제거해 건강한 삶을 영위
하게 할 수 있는 새로운 물질로 생각되어지고 있다.
플라보노이드의 세포 신호전달 조절 기전
플라보노이드는 phosphoinositide 3-kinase (PI 3-kinase), Akt/PKB, tyrosine kinases, protein kinase C (PKC), 그리고 MAP kinases 같은 여러 개의 인산화 효소(protein kinase)들 의 신호 전달을 선택적으로 조절하여 세포 안에서 항산 화 작용의 조절에 영향을 미칠 것이라고 보고되고 있 다.15) 이와 같은 신호 전달 체계의 억제나 촉진 작용은 표적 세포의 단백질 인산화 상태를 바꾸거나 유전자 발 현을 조절함으로써 세포 기능에 중대하게 영향을 미칠 가능성이 크다. 인산화 효소의 단계적 반응에서의 선택 적 억제 작용은 암이나 다양한 질병, 염증, 그리고 신경 퇴화를 예방하는데 기여할 수 있으며, 또한 선택적 억제 작용은 특히 미성숙 신경 세포의 성장기 동안 단백질과 lipid kinase가 신경연접형성(synaptogenesis)과 신경돌기 (neurite)의 성장을 조절할 때에도 작용할 수 있다. 성숙한 뇌 안에서의 신경세포는 MAP kinase와 시냅스 가소성(可 塑性-synaptic plasticity)과 기억의 구성(memory formation) 같은 중요한 기능에 작용하는 PI 3-kinase와의 단계적 반 응에 대해 보고되고 있으며,16) 이러한 이유로 위의 신호 전달 경로와 연계되는 플라보노이드의 상호작용은 예측 할 수 없는 결과를 초래할 수 도 있다. 그리고 신호 전달
경로와 연계되는 플라보노이드의 상호 작용은 세포의 종류에 따라 달라질 수 있을 것이라고 알려져 있다. 플라 보노이드의 세포에서의 효과는 궁극적으로 플라보노이 드가 세포질(cytosol)로 흡수되는 것과 플라보노이드가 막에서 어떤 상호 작용을 하느냐에 달려 있는데, 플라보 노이드의 흡수와 플라보노이드가 물질 순환과 세포 상 호 작용에 미치는 효과에 대한 정보는 새로운 관점으로 써 점점 더 중요해 지고 있다. 이러한 대사 물질들은 GSH을 함유하고 있는 산화적 대사 물질들을 포함하고 있다(Fig. 1). 상대적으로 몇몇 연구들은 신호 전달 기전 의 조절에 작용하는 대사 물질들의 능력에 초점을 맞추 고 있다. 플라보노이드는 미토콘드리아 ATPase,17) calcium plasma membrane ATPase,18) protein kinase A,19) protein kinase C, 그리고 topoisomerase20)을 포함하는 단백질의 수 많은 ATP-binding sites21)에 결합할 수 있는 가능성을 가진 것으로 보고되어 지며, 플라보노이드가 GABA-A 수용체 에 결합하는 benzodiazapine과 adenosine 수용체22)와 상호 작용한다는 것도 보고되었다. Resveratrol과 citrus flava- none, hesperetin, 그리고 naringenin은 몇몇 protein kinase에 서 억제 작용을 가지고 있다고 보고되었다.23) 아마도 이 러한 작용은 폴리페놀 결합 단계와 ATP binding site 결합 단계를 거치면서 억제 작용의 비(非)활동성을 유도하는 인산화효소 내에서의 3차원적 구조 변화를 일으키며 발 생 할 것으로 예상되어 진다. 또한 플라보노이드는 미토 콘드리아와 상호 작용하기도 하고 대사 작용 기전의 중 간 단계에 끼어들기도 하며, 유착 분자의 표현형을 조절 하기도 한다고 보고되어 지고 있다. 몇몇의 연구들은 플 라보노이드가 protein kinase C, tyrosine kinase의 주된 억제 제로 작용하는지 그렇지 않은지는 플라보노이드의 구조 에 달려 있을 수 있다고 보고되었다. LY294002는 querc- etin 구조의 표본으로 알려져 있다. LY294002와 quercetin 은 서로 무척 다른 정위(定位)에도 불구하고 효소 주머니 를 결합하는 ATP에 알맞게 작용한다고 보고되었다.24) B- 고리에 있는 수산기 그룹의 개수와 치환, 그리고 C2-C3
bond 불포화 화합물의 정도는 플라보노이드 생물활성의 중요한 결정원이 된다고 알려져 있다. 생체 내 대사물질 에서 quercetin이 pro-survival Akt/PKB signaling pathway를 방해한다는 점에서 무척 흥미로운데, 이처럼 플라보노 이드는 중요한 신호 기전 분자들과 상호 작용을 할 수 있다. 예를 들면 플라보노이드가 산화적 스트레스에 의 해 야기되는 세포사를 방지한다는 것은 c-jun amino- terminal kinase (JNK)의 활성화를 막음으로써 이루어진다 고 보고되었다. 그리고 신호를 JNK로 보내는 단백질을 활성화 시키는 많은 MAPKKK 중 하나에 영향을 미침으
로써 산화적 스트레스에 의한 세포사를 방지할수 있다 는 연구가 되고 있다. 플라보노이드가 MAPK 활성화에 매우 중요한 역할을 하는 calcium 항상성 유지에 중요하 다는 보고들도 있다.25) 또한 플라보노이드는 미토콘드리 아와 함께 상호 작용을 할 것이라고 알려져 있는데, 예를 들면, 세포사가 진행되는 동안에 플라보노이드는 cytoch- rome C (세포의 산화 환원에 작용 하는 색소 단백질)의 분비를 조절하는 mitochondrial transition pore (mPT)를 조 정하거나 DIABLO/smac 같은 미토콘드리아와 관련된 다 른 세포사 조절인자들을 조절함으로써 미토콘드리아와 상호 작용을 한다고 보고되어 지고 있다.26) 세포사가 진 행되는 동안에 transition pore는 기공의 개방과 cytochrome C의 분비를 조절하는 곳에 결합하는 benzodiazapine을 흡
수한다는22,23) 사실은 mPT와의 상호 작용 가능성을 제시
하고 있다. MAP kinase pathways는 ERK1/2, JNK 그리고 p38 MAP kinase로 구성 되어 있는데, 플라보노이드에 의 한 MAP kinase signalling pathways 조절시 ERK1/2와 JNK 가 growth-factor (T-세포분열, 세포분화, 세포사, 그리고 다양한 세포 가소성을 유도하는 것)와 연관 되어 있다고 보고되고 있다.27) ERK와 JNK는 일반적으로 신경 세포사 에 작용하는 것이라 알려져 있고,28) ERK1/2는 pro-survival signalling과 연관돼 있다고 알려져 있으며, 최근에는 항 암제에 의한 ERK의 활성이 세포사멸을 유도하고 플라보 노이드의 한 종류인 3,4‘-dihydroxyflavone이 ERK의 활성 을 저해해 세포사멸을 저해한다는 보고가 되고 있다.29) 이러한 보고들에 의해 플라보노이드가 세포 신호전달분 자의 저해제로의 충분한 가능성 뿐만 아니라 세포사멸 을 조절 할 수 있는 능력을 가졌다고 생각되어진다. Pro- survival signalling은 cyclic AMP regulatory binding protein (CREB)의 활성화, anti-apoptotic protein Bcl-2의 발현 증가, BAD의 비 (非)전사의 억제30)와 관련된 작용기전을 통하 여 이루어진다. JNK는 transcription-dependent apoptotic signalling과 관련돼 있으며, Transcription-dependent apopto- tic signalling은 c-Jun의 활성화, JunB, JunD, ATF-2를 포함 하는 AP-1 protein을 통하여 이루어진다. 산화 스트레스 는 세포 속 신호 기전에 다양한 영향을 미치며 특히 MAP kinase cascade에 다양한 영향을 주는데, 세포 산화 환원반응의 변화는 세포 속에서 세포사 기전을 시작하 게 하는 JNK같은 pro-apoptotic signalling proteins를 활성화 하는 결과를 초래한다고 알려져 있다. 산화 스트레스는 mitochondrial transition pore (mPT)와 cytochrome C의 분비 에 영향을 줌으로써 미토콘드리아에 작용할 것이라고도 보고되어 지고 있다.31) 발생적이거나 퇴행적인 death signalling에서 다양한 pro-apoptotic stimuli에 의해 초래되
이응룡 외 7인:Regulation of Cellular Signal Transduction by Flavonoids 169
는 신경 뉴런의 감소에 JNK의 활성화가 관련이 있다는 보고가 있다. 뉴런 속에 산화적 생성물의 생성 하에서 JNK는 dopamine과32) 4-HNE,30) SOD1의 발현 감소,33) hyd- rogen peroxide, oxLDL14)에 의해 활성화된다. 플라보노이 드는 위에서 설명한 활성화를 막는 것으로 알려져 있으 며 MAP kinase 이외에도 serine/threonine kinase인 Akt/PKB, PI 3-kinase, 그리고 pivotal kinase를 통한 신호전달의 조절 에서도 중요하다(Fig. 2). 또한 뉴런 안에서 Akt/PKB의 활 성화는 cell death machinery로 작용하는 단백질들을 억제 하는 것으로 보여진다. 즉, pro-apoptotic Bcl-2 family member BAD 와 poly (ADP-ribose) polymerase34)의 절편화 를 저해하고 세포사멸을 유도하는 caspase family들을 저 해한다. BAD는 serine의 잔재물인 Ser112와 Ser13635)의 인 산화반응에 의해 조절된다. 몇몇 연구들은 Serine 136 site 가 특별히 Akt/PKB에 의해 인산화반 응이 일어난다고 보고되고 있다.36) 여기서 플라보노이드는 위의 단계적 반응을 조절해서 세포사멸에 작용한다고 보고되고 있 다.
대표적인 플라보노이드인 Epigallocatechin Gallate (EGCG)의 세포 내 신호전달 조절 기작
플라보노이드 여러 그룹 중 플라보놀의 효과에 대해 많은 연구가 진행되고 있는데, 플라보놀의 예를 들면 epicatechin, epicatechin gallate (EGC)와 epigallocatechin gallate (EGCG) 등이 있다. 이들 폴리페놀의 세포 보호적 성질이 보고되고 있는데, 예를 들어 epicatechin과 체내 대사물질 들에 함유된 epicatechin 성분 중 하나와 3V-O-methyl epicatechin은 섬유아 세포와 뉴런 세포에서 산화적 스트 레스에 대해 강한 세포 보호적 효과를 유도한다고 보고 되고 있다.5,37,38) 이런 일련의 세포 보호적 작용을 하는 물질들은 caspase-3 활성화15,37)의 억제와 pro-apoptotic MAPK 단백질15)의 활성화와 관련돼 있는 것으로 밝혀졌 다. Epicatechin과 3V-O-methyl-epicatechin은 JNK의 oxLDL- induced activation과 c-jun, 그리고 pro-caspase-3에 대하여 뉴런을 보호하는 작용을 한다고 밝혀졌다. 플라보노이 드에 의한 JNK의 억제는 Bax, 미토콘드리아 기능장애, 그리고 caspases의 활성화와 같은 pro-apoptotic 단백질들 의 c-jun/AP-1-mediated regulation의 제거를 유도한다고 보 고되고 있다. 흥미롭게도 epicatechin과 epicatechin의 O-메 틸화 형태 대사물질은 세포 내 산화적 스트레스 안에서 oxLDL에 대해 별다른 영향을 미치지 않는다고 알려져 있다. 위의 연구 결과들은 catechol 구조가 mono-methyl- ation에 의해 막혀짐으로써 protection이 H-donating antio-
xidant processes에 의해 우선적으로 성립되는 것이 아니라 는 것을 알려준다. 다른 연구에서 보면 세포사멸을 유도 하는 산화적 스트레스에 대한 EGCG의 신경 세포 보호 적 메카니즘은 신호 단백질의 조절과 연관된 것으로 보 고되었다. EGCG는 PKC의 활성화와 Bax, Bad, Mdm2, Bcl-2, Bcl-w, 그리고 Bcl-x (L)와 같은 cell survival/cell cycle 관련 유전자들의 조절을 야기한다고 보고되었다.39) 여러 연구들에 의하면 세포 보호 작용은 어떤 경로를 통해, 그리고 어떠한 세포 상호 작용에 의해 발생하는지 정확 히 알 수 는 없지만, 세포 보호 작용은 부분적으로 신호 기전의 특별한 작용을 통하여 이루어지는 것을 알 수 있 다. 또한, 플라보노이드는 암세포의 증식과 관련된 성장 관련 신호 전달 경로에 어떤 영향을 미칠 것이라 보고되 었다.40) 이들 결과들로부터 플라보노이드의 작용은 다음 두 가지로 볼 수 있다. (1) 플라보노이드는 암세포의 세 포사멸을 유도할 수 있는 기능을 가졌고 반면에 보통 세 포에 대해서는 세포사를 유발시키지 않는다고 보고되었 다. (2) 플라보노이드는 발암현상의 증대에 관련된 신호 기전을 억제한다고 알려져 있다. 많은 연구들이 암세포 의 세포사를 유도하는 EGCG같은 플라보놀의 기능을 실 험했는데, 최근에 들어서는 플라보노이드가 어떠한 작 용 기전을 통해 세포사를 유도하는지에 대해 실마리가 풀리고 있다. 예를 들면 human leukaemia U937 세포의 세 포사를 EGCG가 유도하는 작용기전은 ASK1, MKK, 그리 고 JNK/p38 cascade와 관련되어 있다고 보고되었으며, 또 한 ERK의 활성화는 U937 세포의 세포사에 관여한다고 보고되었다.41) 또한 EGCG에 의한 JNK의 인산화 반응과 ERK의 억제는 arsenite-induced apoptosis의 억제에 중요한 역할을 한다고 보고되었다.42) EGCG와 녹차의 폴리페놀 성분은 암세포 성장의 억제와 AP-1 activity의 억제를 유 도한다 보고되었고,43) 많은 암세포 종류에서 발생하는 ras 유전자의 돌연변이는 성장 신호 전이 기전을 촉진시 키고 전사인자인 AP-1과 관련되어 있다고 보고되었다.44) EGCG의 항암효과는 PDGF-Rh tyrosine phosphorylation의 선택적인 억제에 의해 일어난다고 보고되어 있으며,45) 혹은 NF-κB inducing kinase (NIK) 조절의 감소, death- associated protein kinase 1, rhoB, 그리고 tyrosine-protein kinase gene과 retinoic acid receptor a1에 해당되는 유전자 조절을 증가시킴으로써 일어난다고 보고되었다.46) EGC 를 이용한 유방암 치료는 세포사멸에 대한 예를 보여주 고 있는데, 이는 anti-apoptotic protein Bcl-2의 감소를 유도 하고 pro-apoptotic인 Bax, caspase-8, 그리고 caspase-10을 증 가를 유도함으로써 세포사멸을 일으킨다고 보고되었다.
지난 연구들에서는 EGC가 일으키는 유방암 세포의 세
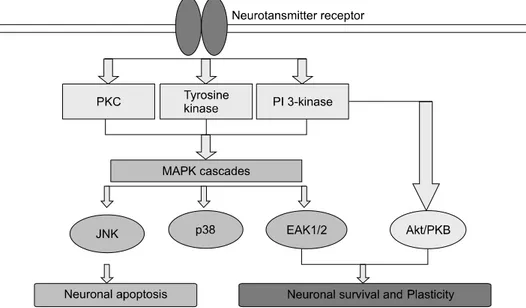
Fig. 5. Potential points of action of flavonoids within MAP kinase and Akt/PKB signalling cascades in neurons. Inhibitory actions within ERK1/2 and Akt/PKB pathways could initiate neuronal death, whereas inhibitory actions within JNK pathways are more likely to be neuroprotective.
포사는 Fas death receptor signaling과 관련이 있을 거라고 예상했었다. 다른 breast cancer cell line에서 EGCG는 cdc2 phosphorylation의 억제를 유도하고 cyclin A, cyclin B1, 그 리고 cdk protein의 발현을 조절하며, G2 세포주기에서 작 용을 멈추는 JNK/SAPK와 p38 MAP kinase의 phosphoryl- ation을 유도하는 것으로 나타났다. EGCG는 head and neck squamous cell carcinoma (HNSCC) 안에서 epidermal growth factor receptor (EGFR)와 연관 신호 기전의 활성화 를 억제하는 것으로 나타났다고 보고되었다.47) EGCG는 신호 전달 중에 EGFR과 연관된 기전과 HER-2/neu의 활 성화와 그 하위 신호전달 기작을 막는 것으로 추정된다 고 보고되고 있다.48) 이런 연구들을 통해 EGCG는 유방 암과 HNSCC의 치료에 유용한 물질이며, 암세포의 증식 과 성장을 유도하는 신호 기전을 막는데 효과가 있을 것 으로 생각 되어진다. 많은 연구들은 EGCG와 다른 플라 보놀 성분들이 자외선에 의해 발생하는 피부 손상을 막 는데도 효과가 있다고 제안하고 있다. 예를 들면 EGCG 는 자외선이 야기하는 산화적 스트레스에 의한 ERK1/2, JNK, 그리고 p38 MAP kinase의 인산화반응을 막는데 효 과적이라는 보고가 있다. 녹차의 폴리페놀 성분과 EGCG 는 자외선이 야기하는 산화적 스트레스에 의한 ERK1/2, JNK, 그리고 p38 MAP kinase의 활성화를 감소시킨다는 사실이 쥐 피부 실험을 통해 증명되었다.49) 흥미로운 사 실은 플라보노이드 성분을 구강을 통해 섭취하는 것보 다 주사기를 이용해 국소적 형태로 주입하는게 더 효과 적이라고 알려져 있다. 위와 같은 연구결과가 도출됨으 로써 아직까지 플라보노이드에 대한 효과의 정확한 증 명과 작용기전에 대한 명확한 이해가 없음에도 불구하
고 하루에도 수많은 플라보노이드 추출물을 포함한 기 능성 화장품이 출시되고 있다. EGCG는 전사인자인 NF- κB 신호전달 분자의 조절을 통해 자외선에 의해 발생하 는 부작용을 막는 것으로 보고되었다.50) NF-κB 기전은 다양한 생물학적 기능에 중요한 역할을 할 뿐 아니라 피 부암 발생에도 깊은 연관성을 가지고 있다. EGCG와 다 른 플라보놀 성분들은 산화적 스트레스와 MAPK가 유발 하는 photo-carcinogenesis 같은 피부 질환의 발생을 감소 시키는데도 효과가 있다고 알려져 있다. Epicatechin과 EGCG같은 플라보놀 성분들은 MAP kinase 기전과 다른 신호기전의 단계적 반응을 조절함으로써 때로는 유익한 효과를 내기도, 혹은 세포 독성적 작용을 하기도 한다.
Pro-survival MAP kinase 신호 전달 분자의 활성은 비교적 낮은 농도의 플라보노이드에서 일어나고 anti-oxidant response element (ARE)-mediated gene expression을 유도한 다. 반면에 높은 농도의 플라보노이드는 세포사의 전이 를 유도하는 JNK의 계속적인 활성화를 유도한다고 보 고 되었다.51)
결 론
플라보노이드는 항산화 기능을 통해서뿐만 아니라 단 백질 및 지질 인산화 효소들의 신호전달계를 조절하여 세포 사멸을 유도하거나, 억제하는 기능을 수행하는 것 으로 보고되고 있다. 즉 플라보노이드와 그들의 대사물 질은 체내에서 주된 항산화 작용을 하는 매개체가 아닐 가능성도 있으며, 수용체, 효소, 그리고 전사 요소에 직 접적인 상호작용을 하여 기능을 수행할 것으로 예상되
이응룡 외 7인:Regulation of Cellular Signal Transduction by Flavonoids 171
고 있다. 실제로 많은 연구들에서 플라보노이드가 여러 가지 단백질 인산화효소의 활성화 조절에 작용한다고 보고되고 있다.15,21∼24,38,39,45,47,51∼53)
이와 같은 플라보노이 드의 인산화 효소 조절은 효소의 ATP 결합 부위에 플라 보노이드가 결합함으로써 여러 작용의 선택성을 감소시 킬 수 있기 때문이다. 계속된 여러 연구들을 통하여 플라 보노이드의 upstream binding partner, regulatory kinase, 그 리고 receptor 등에 대해 조사하여 플라보노이드의 작용 구간을 정의하려고 했지만 아직까지 정확하게 밝혀지지 않고 있다. 예를 들면 Akt/PKB의 억제는 플라보노이드 가 PI 3-kinase에 작용하여 일어날 것으로 예상하며, ERK1/2의 작용은 플라보노이드가 조절하는 upstream regulatory kinase나 GTPase같은 MAP kinase 반응물의 상호 작용 분자들에 의해 일어난다. 이러한 작용들은 증식성 질병에는 유익한 영향을 미치지만, 신경세포 조직에는 해로운 영향을 미칠 것이다(Fig. 5). 그런 까닭에 세포 간 신호 전달 기전과 함께 상호적으로 작용하는 플라보노 이드는 예측할 수 없는 결과를 가져올 수도 있다. 플라보 노이드의 작용은 그 구조에 따라서 세포의 종류, 질병 등에 따라 다른 결과를 도출함을 알 수 있다.
감사의 글
This work was supported by the Korea Research Foundation grant funded by the Korean Government (MOEHRD) (KRF- 2006-312-C00627 and KRF-2005-070-C00095) and the Korea Science & Engineering Foundation (KOSEF) grant funded by the Korea Government (MOST) (No. R11-2002-100-01000-0).
참 고 문 헌
1) Rice-Evans, C. Flavonoid antioxidants. Curr Med Chem 8, 797- 807, 2001.
2) Spencer JP, Schroeter E, Rechner H, Rice-Evans AR.
Bioavailability of flavan-3-ols and procyanidins: gastrointes- tinal tract influences and their relevance to bioactive forms in vivo. Antioxid Redox Signal 3, 1023-1039, 2001.
3) Scheline, RR. Metabolism of oxygen heterocyclic compounds.
In: CRC Handbook of Mammalian Metabolism of Plant Compounds. Boca Raton, FL: CRC Press, pp 243–295, 1999.
4) Spencer JP, Abd El Mohsen MM, Rice-Evans C. Cellular uptake and metabolism of flavonoids and their metabolites:
implications for their bioactivity. Arch Biochem Biophys 423, 148-461, 2004.
5) Spencer JP, Schroeter H, Crossthwaithe AJ, Kuhnle G, Williams RJ. Rice-Evans, C. Contrasting influences of glucu-
ronidation and O-methylation of epicatechin on hydrogen peroxide-induced cell death in neurons and fibroblasts. Free Radic Biol Med 31, 1139-1146, 2001.
6) Rice-Evans CA, Miller NJ, Paganga G. Structure–antioxidant activity relationships of flavonoids and phenolic acids. Free Radic Biol Med 20, 933-956, 1996.
7) Miyake Y, Shimoi K, Kumazawa S, Yamamoto K, Kinae N, Osawa T. Identification and antioxidant activity of flavonoid metabolites in plasma and urine of eriocitrin-treated rats. J Agri Food Chem 48, 3217-3224, 2000.
8) Abd El Mohsen MM, Kuhnle G, Rechner AR, Schroeter H, Rose S, Jenner P, Rice-Evans CA. Uptake and metabolism of epicatechin and its access to the brain after oral ingestion. Free Radic Biol Med 33, 1693-1702, 2002.
9) De Whalley CV, Rankin SM, Hoult JRS, Jessup W, Leake DS. Flavonoids inhibit the oxidative modification of low density lipoproteins by macrophages. Biochem Pharmacol 39, 1743-1750, 1990.
10) Beckman J, Chen S, Ischiropoulos H. Cow J. Oxidative chemistry of peroxynitrite. Meth Enzymol 233, 229-240, 1994.
11) Pannala AS, Razaq R, Halliwell B, Singh S. Inhibition of peroxynitrite dependent tyrosine nitration by hydroxycin- namates: nitration or electron donation. Free Radic Biol Med 24, 1173-1178, 1998.
12) Morel I, Cillard P, Cillard J. flavonoid-metal interactions in biological system. In: eds, by Rice-Evans C, Parker L, 1st ed, Marcel Dekker Inc. Flavonoids in Health and Disease, pp 163-197, 1998.
13) Schroeter H, Spencer JP, Rice-Evans C, Williams RJ.
Flavonoids protect neurons from oxidized low-density-lipo- protein-induced apoptosis involving c-Jun N-terminal kinase (JNK), c-Jun and caspase-3. Biochem J 358, 547-557, 2001.
14) Lee ER, Kang YJ, Kim JH, Lee HT, Cho SG. Modulation of Apoptosis in HaCaT Keratinocytes via Differential Regulation of ERK Signaling Pathway by Flavonoids. J Biol Chem 280, 31498-31507, 2005.
15) Perkinton MS, Sihra TS, Williams RJ. Ca (2+)-permeable AMPA receptors induce phosphorylation of cAMP response element-binding protein through phosphatidylinositol 3-kinase- dependent stimulation of the mitogen-activated protein kinase signaling cascade in neurons. J Neurosci 19, 5861-5874, 1999.
16) Di Pietro A, Godinot C, Bouillant ML, Gautheron DC. Pig heart mitochondrial ATPase: properties of purified and mem- brane-bound enzyme. Biochimie 57, 959-967, 1975.
17) Barzilai A, Rahamimoff H. Inhibition of Ca2+-transport ATPase from synaptosomal vesicles by flavonoids. Biochim Biophys Acta 730, 245-254, 1983.
18) Revuelta MP, Cantabrana B, Hidalgo A. Depolarization- dependent effect of flavonoids in rat uterine smooth muscle contraction elicited by CaCl2. Gen Pharmacol 29, 847-857, 1997.
19) Boege F, Straub T, Kehr A, Boesenberg C, Christiansen K, Andersen A, Jakob F, Kohrle J. Selected novel flavones inhibit
the DNA binding or the DNA religation step of eukaryotic topoisomerase I. J Biol Chem 271, 262-227, 1996.
20) Conseil G, Baubichon-Cortay H, Dayan G, Jault JM, Barron D, Di Pietro A. flavonoids: class of modulators with bifunctional interactions at vicinal ATP-and steroid-binding sites on mouse P-glycoprotein. Proc Natl Acad Sci 95, 9831- 9836, 1998.
21) Medina JH, Viola H, Wolfman C, Marder M, Wasowski C, Calvo D, Paladini AC. Overview-flavonoids: new family of benzodiazepine receptor ligands. Neurochem. Res. JID-7613461.
22, 419-425, 1997.
22) Fischer PM, Lane DP. Inhibitors of cyclin-dependent kinases as anti-cancer therapeutics. Curr Med Chem 7, 1213-1245, 2000.
23) Walker EH, Pacold ME, Perisic O, Stephens L. Hawkins PT, Wymann MP. Williams, R. L. Structural determinants of phosphoinositide 3-kinase inhibition by wortmannin, LY- 294002, quercetin, myricetin, and staurosporine. Mol Cell 6, 909-919, 2000.
24) Enslen H, Tokumitsu H, Stork PJ, Davis RJ, Soderling TR.
Regulation of mitogen-activated protein kinases by calcium/
calmodulin-dependent protein kinase cascade. Proc Natl Acad Sci 93, 10803-10808, 1996.
25) Goyal L. Cell death inhibition: keeping caspases in check. Cell 104, 805-808, 2001.
26) Herdegen T, Skene P, Bahr M. The c-Jun transcription factor:
bipotential mediator of neuronal death, survival and regeneration. Trends Neurosci 20, 227-231, 1997.
27) Xia Z, Dickens M, Raingeaud J, Davis RJ, Greenberg ME.
Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 270, 1326-1331, 1995.
28) Anderson CN, Tolkovsky AM. Role for MAPK/ERK in sympathetic neuron survival: protection against p53-depen- dent, JNK-independent induction of apoptosis by cytosine arabinoside. J Neurosci 19, 664-673, 1999.
29) Lee ER, Kim JH, Kang YJ, Cho SG. The anti-apoptic and anti-oxidant effect of eriodictyol on UV-induced apoptosis of keratinocyte. Biol Pharm Bull 30, 32-37, 2007
30) Lemasters JJ, Qian T, He L, Kim JS, Elmore SP, Cascio WE, Brenner DA. Role of mitochondrial inner membrane permea- bilization in necrotic cell death, apoptosis, and autophagy.
Antioxid. Redox Signal 4, 769-781, 2002.
31) Luo Y, Umegaki H, Wang X, Abe R, Roth GS. Dopamine induces apoptosis through an oxidation-involved SAPK/JNK activation pathway. J Biol Chem 273, 3756-3764, 1998.
32) Wang L, Matsushita K, Araki I, Takeda M. Inhibition of c-Jun N-terminal kinase ameliorates apoptosis induced by hydrogen peroxide in the kidney tubule epithelial cells (NRK-52E). J Neurosci 91, 142-7, 2002.
33) Kennedy SG, Wagner AJ, Conzen SD, Jordan J, Bellacosa A, Tsichlis PN, Hay N. The PI 3-kinase/Akt signaling pathway delivers an anti-apoptotic signal. Breast Cancer Res Treat 11, 701-13, 1997.
34) Vergote D, Cren-Olive C, Chopin V, Toillon RA. Rolando, survival factor results in binding to 14-3-not BCL-X(L). Cell 87, 619-628, 1996.
35) del Peso L, Gonzalez-Garcia M, Page C, Herrera R, Nunez G. Interleukin-3-induced phosphorylation of BAD through protein kinase Akt. Science 278, 687–689, 1997.
36) Spencer JPE, Schroeter H, Kuhnle G, Srai SK, Tyrrell RM, Hahn U, Rice-Evans C. Epicatechin and its in vivo metabolite, 3-VO-methyl epicatechin, protect human fibroblasts from oxidative-stress-induced cell death involving caspase-3 activa- tion. Biochem J 354, 493-500, 2001.
37) Schroeter H, Williams RJ, Matin R, Iversen L, Rice-Evans CA.
Phenolic antioxidants attenuate neuronal cell death following uptake of oxidized low-density lipoprotein. Free Radic Biol Med 29, 1222-1233, 2000.
38) Levites Y, Amit T, Youdim MB, Mandel S. Involvement of protein kinase C activation and cell survival/cell cycle genes in green tea polyphenol (-)-epigallocatechin 3-gallate neuro- protective action. J Biol Chem 277, 30574-30580, 2002.
39) Bode AM, Dong Z. Signal transduction pathways: targets for chemoprevention of skin cancer. Lancet Oncol 1, 181-188, 2000.
40) Choi YJ, Lim SY, Woo JH, Kim YH, Kwon YK, Suh S, Lee SH, Choi WY, Kim J G, Lee IS, Park JW, Kwon TK. Sodium orthovanadate potentiates EGCG-induced apoptosis that is dependent on the ERK pathway. Bioche Biophys Res Commun 305, 176-185, 2003.
41) Chen NY, Ma WY, Yang CS, Dong Z. Inhibition of arsenite-induced apoptosis and AP-1 activity by epigallo- catechin-3-gallate and theaflavins. J Environ Pathol Toxicol Oncol 19, 287-295, 2000.
42) Chung JY, Huang C, Meng X, Dong Z, Yang CS. Inhibition of activator protein 1 activity and cell growth by purified green tea and black tea polyphenols in H-ras-transformed cells: structure-activity relationship and mechanism involved.
Cancer Res 59, 4610-4617, 1999.
43) Zachos G, Spandidos DA. Expression of ras proto-oncogenes:
regulation and implications in the development of human tumors. Crit Rev Oncol Hematol 26, 65-75, 1997.
44) Ahn HY, Hadizad, KR, Seul C, Yun YP, Vetter H, Sachinidis A. Epigallocathechin-3 gallate selectively inhibits the PDGF- BB-induced intracellular signaling transduction pathway in vascular smooth muscle cells and inhibits transformation of sis-transfected NIH 3T3 fibroblasts and human gliblasttoma cells (A172). Mol Biol Cell 10, 1093-1104, 1999.
45) Okabe S, Fuhimoto N, Sueoka N, Suganuma M, Fujiki H.
Modulation of gene expression by (-)-epigallocatechin gallate in PC-9 cells using a cDNA expression array. Biol Pharm Bull 24, 883-886, 2001.
46) Masuda M, Suzui M, Weinstein IB. Effects of epigallo- catechin-3-gallate on growth, epidermal growth factor receptor signaling pathways, gene expression, and chemosen- sitivity in human head and neck squamous cell cercinoma cell
이응룡 외 7인:Regulation of Cellular Signal Transduction by Flavonoids 173
lines. Clin Cancer Res 7, 4220-4229, 2001.
47) Masuda M, Suzui M, Lim JT, Deguchi A, Soh JW, Weinstein IB. Epigallocatechin-3-gallate decreases VEGF production in head and neck and breast carcinoma cells by inhibiting EGFR-related pathways of signal transduction. J Exp Ther Oncol 2, 350-359, 2002.
48) Vayalil PK, Elmets CA, Katiyar SK. Treatment of green tea polyphenols in hydrophilic cream prevents UVB-induced oxidation of lipids and proteins, depletion of antioxidant enzymes and phosphorylation of MAPK proteins in SKH- hairless mouse skin. Carcinogenesis 24, 927-936, 2003.
49) Afaq F, Adhami VM, Ahmad N, Mukhtar H. Inhibition of ultraviolet B-mediated activation of nuclear factor kappaB in normal human epidermal keratinocytes by green tea consti- tuent(-)-epigallocatechin-3-gallate. Oncogene 22, 1035-1044, 2003.
50) Chen C, Yu R, Owuor ED, Kong AN. Activation of
antioxidant-response element (ARE), mitogen-activated protein kinases (MAPKs) and caspases by major green tea polyphenol components during cell survival and death. Arch.
Pharm. Res 23, 605-612, 2000.
51) Schroeter H, Boyd C, Spencer JPE, Williams RJ, Cadenas E, Rice-Evans C. MAPK signaling in neurodegeneration: influ- ences of flavonoids and of nitric oxide. Neurobiol Aging 23, 861- 880, 2002.
52) Ishikawa Y, Kitamura M. Anti-apoptotic effect of quercetin:
intervention in the JNK-and ERK-mediated apoptotic path- ways. Kidney Int 58, 1078-1087, 2000.
53) Chung JY, Park JO, Phyu H, Dong Z, Yang CS. Mechanisms of inhibition of the Ras-MAP Kinase signaling pathway in 30.
7b Ras 12 cells by tea polyphenols (-)-epigallocatechin-3- gallate and theaflavin-3, 3'-digallate. FASEB J 15, 2022- 2024, 2001.