51 This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://
creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
J. Mushrooms 2018 March, 16(1):51-56 http://dx.doi.org/10.14480/JM.2018.16.1.51 Print ISSN 1738-0294, Online ISSN 2288-8853
© The Korean Society of Mushroom Science
*Corresponding author
E-mail : [email protected], [email protected] Tel : +82-43-261-2293
†These authors contributed equally to this work.
Received February 27, 2018 Revised March 13, 2018 Accepted March 23, 2018
표고버섯 품종 ‘산마루2호’를 구분할 수 있는 CAPS marker 개발
문수윤1,† · 이화용1,2,† · 가강현3 · 구창덕2,* · 류호진1,*
1충북대학교 생물학과
2충북대학교 산림학과
3국립산림과학원 화학미생물과
Development of a CAPS marker for the identification of the Lentinula edodes cultivar, ‘Sanmaru 2ho’
SuYun Moon1,†, Hwa-Yong Lee1,2,†, Kang-Hyeon Ka3, Chang-Duck Koo2,*, and HoJin Ryu1,*
1Department of Biology, Chungbuk National University, Cheongju 28644, Korea
2Department of Forest Science, Chungbuk National University, Cheongju 28644, Korea
3Division of Wood Chemistry and Microbiology, National Institute of Forest Science, Seoul 02455, Korea
ABSTRACT: In Korea, the oak mushroom (Lentinula edodes) is highly preferred by consumers in the food industry and makes up about 97.7% of the total forest mushroom production. This indicates that the oak mushroom is an important non-timber forest product in Korea. Recently, the breeding and development of new cultivars of L. edodes have been actively initiated, and the development of molecular markers that are able to identify and discriminate the new cultivars is crucial for protecting the breeder’s rights. This study was carried out to develop a cleaved amplified polymorphic sequence (CAPS) marker for the identification and discrimination of a new cultivar, Sanmaru 2ho from the 37 other oak mushroom cultivars. A single nucleotide polymorphism (SNP) was identified at the 1,803,483rd position of scaffold2 in the genome of Sanmaru 2ho. The amplified DNA containing the SNP of Sanmaru 2ho was uniquely not cleaved by the restriction enzyme, Hha I, and thus Sanmaru 2ho was successfully distinguished from the other oak mushroom cultivars.
KEYWORDS: CAPS marker, Lentinula edodes, Single nucleotide polymorphism, Cultivar identification
표고버섯은 Agaricales 목 Omphalotaceae과 Lentinula 속으로 (International Mycological Association, 2018) 산 림 내 쓰러진 참나무류 등의 활엽수에서 서식하는 백색부 후균이다. 표고는 한국, 중국, 일본 등 아시아 국가에서 일반적으로 재배되는 식용버섯으로(Bak et al, 2013), β-
glucan인 렌티난 등을 함유하고 있어 항암효과가 뛰어난 것으로 알려져 있다(Ng and Yap, 2002; Rop et al., 2009; Bisen et al., 2010).
표고버섯은 2010년 전세계 버섯생산 중 양송이(30%), 느타리(27%)에 이어 약 17%를 차지하여 세 번째로 많 이 생산된 버섯이었다(Royse, 2014). 국내의 경우 2016 년 생산된 단기소득임산물 중 건표고와 생표고를 포함한 표고버섯의 생산량은 약 23,470 톤으로, 전체 임산버섯 생산량인 약 24,014 톤의 약 97.7%를 차지하였다(Korea Forest Service, 2017). 우리나라 국민의 버섯 선호도는 표고버섯(20.6%), 큰느타리버섯(13.4%), 느타리버섯 (12.9%), 양송이(11.1%), 팽이(8.9%) 순으로, 표고버섯에 대한 국민 선호도가 매우 높았다(Korea Rural Economic Institute, 2016). 따라서 표고버섯은 중요한 단기소득임 산물이다.
국민의 선호도가 높아짐에 따라, 표고버섯은 생산량에 비하여 소비량이 증대되고 있다. 2016년 국내 표고버섯의 생산량은 건표고가 약 1,036 톤, 생표고가 약 22,433 톤
으로 총 약 23,469 톤 이며(Korea Forest Service, 2017), 수출량은 약 175 톤, 그리고 수입량은 약 17,165 톤이었 다(KATI, 2017). 특히 2003년 약 1,926 톤이었던 표고버 섯의 수입량은, 2016년 약 17,165 톤으로 약 9배 정도 증 가했으며, 수출량은 2003년 약 374 톤에서 2016년 약 175 톤으로 감소하였다(KATI, 2017). 국내에서 표고버섯 의 소비가 대폭 증가하였으며, 대부분 수입에 의존하는 것을 알 수 있다. 이에 따라 국내에서는 많은 품종들이 개 발되고 있고, 개발된 품종들에 대한 보호가 중요한 문제 로 대두되었다.
표고버섯은 2008년 밤나무, 느티나무, 벚나무, 대추, 감, 천마 등과 같이 ‘종자산업법’에 의하여 산림분야 품종보 호 대상 작물로 지정되었으며, 2010년 나고야 의정서가 채택됨에 따라 유전자원에 대한 보호가 강화되고 있다.
이에 개발되는 품종들은 각 품종들 간의 구별성에 대한 중요성이 증대되고 있다. 현재 개발된 품종에 대한 구별 성은 산림청에서 제작한 표고버섯 특성조사요령을 이용하 고 있다(Korea Forest Service, 2008). 이러한 형태를 이 용한 구별 이외에 분자마커를 이용한다면 더욱 효과적일 것이다. 표고버섯 품종 및 야생 균주의 구분, 다양성 등을 분석하기 위하여 개발, 이용된 분자마커로는 restriction fragment length polymorphism (RFLP) (Kulkarni, 1991), random amplified polymorphic DNA (RAPD)(Zhang and Molina, 1995; Fu et al., 2010; Wu et al., 2010), amplified fragment length polymorphism (AFLP)
(Terashima and Matsumoto, 2004), simple sequence repeat (SSR) (Xiao et al., 2010; Lee et al., 2017; Moon et al., 2017a), inter-simple sequence repeat (ISSR) (Fu et al., 2010; Zhang et al., 2007; Liu et al., 2015), sequence-characterized amplified region (SCAR) (Wu et al., 2010; Qin et al., 2006; Li et al., 2008; Liu et al., 2012), sequence-related amplified polymorphism (SRAP) (Fu et al., 2010; Liu et al., 2015), cleaved amplified polymorphic sequence (CAPS) (Moon et al., 2017b) 등이 있다. 이중 CAPS 마커는 DNA 염기서열 상에 생길 수 있 는 변이인 SNP (single nucleotide polymorphism)와 Indel (insertion and deletion)로 인해 생기거나 사라진 제한효소 의 인식부위를 이용하는 방법이다(Konieczny and Ausubel, 1993). Moon 등 (2017b)은 Shim 등 (2016)이 분석한 표고 버섯의 genome 정보를 이용하여 표고버섯 품종 ‘산마루1 호’와 ‘천장3호’를 구분할 수 있는 CAPS 마커를 개발하 였다.
최근 국립산림과학원에서 개발된 ‘산마루2호’는 고온성, 톱밥재배용 품종으로, 버섯은 평반구형으로 발생하며, 갓 의 직경은 약 64 mm, 두께는 약 23 mm, 그리고 발이온 도가 평균 21~29oC로 고온기에 재배가 가능한 품종이다 (National Institute of Forest Science, 2016). 이 연구에서 는 표고버섯의 genome정보를 기반으로 ‘산마루2호’를 구 분할 수 있는 CAPS마커를 개발하였다.
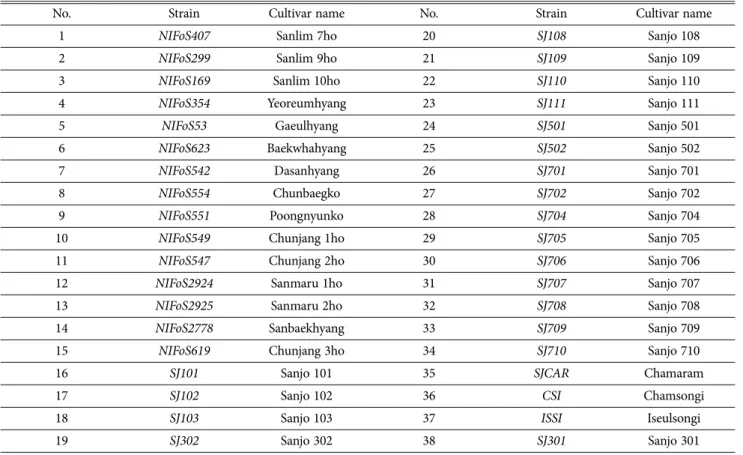
Table 1. List of Lentinula edodes cultivars for CAPS marker development
No. Strain Cultivar name No. Strain Cultivar name
1 NIFoS407 Sanlim 7ho 20 SJ108 Sanjo 108
2 NIFoS299 Sanlim 9ho 21 SJ109 Sanjo 109
3 NIFoS169 Sanlim 10ho 22 SJ110 Sanjo 110
4 NIFoS354 Yeoreumhyang 23 SJ111 Sanjo 111
5 NIFoS53 Gaeulhyang 24 SJ501 Sanjo 501
6 NIFoS623 Baekwhahyang 25 SJ502 Sanjo 502
7 NIFoS542 Dasanhyang 26 SJ701 Sanjo 701
8 NIFoS554 Chunbaegko 27 SJ702 Sanjo 702
9 NIFoS551 Poongnyunko 28 SJ704 Sanjo 704
10 NIFoS549 Chunjang 1ho 29 SJ705 Sanjo 705
11 NIFoS547 Chunjang 2ho 30 SJ706 Sanjo 706
12 NIFoS2924 Sanmaru 1ho 31 SJ707 Sanjo 707
13 NIFoS2925 Sanmaru 2ho 32 SJ708 Sanjo 708
14 NIFoS2778 Sanbaekhyang 33 SJ709 Sanjo 709
15 NIFoS619 Chunjang 3ho 34 SJ710 Sanjo 710
16 SJ101 Sanjo 101 35 SJCAR Chamaram
17 SJ102 Sanjo 102 36 CSI Chamsongi
18 SJ103 Sanjo 103 37 ISSI Iseulsongi
19 SJ302 Sanjo 302 38 SJ301 Sanjo 301
‘산마루2호’를 구분할 수 있는 CAPS마커 개발
‘산마루2호’를 구분할 수 있는 CAPS마커를 개발하기 위하여 국내에서 개발된 38 개 표고버섯 품종(Table 1)과 표고버섯 표준유전체 분석 균주인 B17 (Shim et al., 2016)을 이용하였다.
시험품종들은 potato dextrose broth (PDB) 배지에서 25oC, 암조건에서 약 2주간 110 rpm으로 진탕배양하였다.
진탕배양 된 균사체들은 미라클로스(miracloth)로 걸러 PBS buffer (135 mM NaCl, 2.7 mM KCl, 4.3 mM Na2HPO4및 1.4 mM KH2PO4)로 세척하고 물기를 제거하 였다. 물기가 제거된 균사 약 100 mg을 액체질소에 얼려 막자사발로 마쇄한 후, GenEx Plant kit (GeneAll, Seoul, Korea)를 이용하여 genomic DNA를 추출하였다. genomic DNA의 추출은 제조사의 프로토콜에 따라 추출하였다. 추 출한 DNA는 Micro-spectrophotometer K5600 (BioFuture Inc., Toyko, Japan)으로 정량 후 20 ng/μL로 희석하여 사 용하였다.
‘산마루2호’의 구분을 위한 CAPS 마커 개발에 적합한 SNP와 Indel의 동정은 Hiseq 2500 platform을 이용하여 분석한 시험품종들의 전장유전체 정보와 Shim 등 (2016) 이 분석한 표고버섯 유전체 정보를 비교하여 수행하였다.
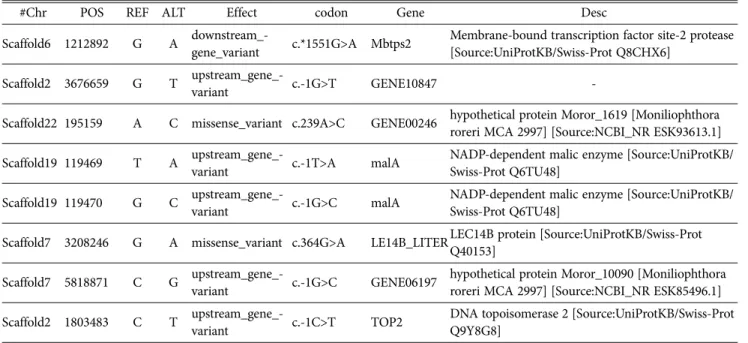
‘산마루2호’에만 존재하는 Indel은 동정이 되지 않았고 SNP는 8개가 동정되었다(Table 2).
동정된 SNP들을 중심으로 500 bp의 flanking sequence 를 추출하였고, Primer3Plus(http://biotools.umassmed.edu/
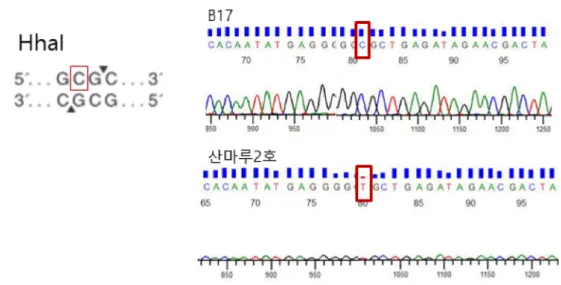
cgi-bin/primer3plus/primer3plus.cgi)를 이용하여 이 서열 들을 증폭할 수 있는 프라이머를 디자인하였다. 프라이머 디자인 후 표고버섯의 표준유전체 정보를 가지고 있는 B17과 본 연구에서 CAPS 마커를 개발하고자 하는 ‘산마 루2호’를 대상으로 genomic DNA 20 ng을 주형으로, 95oC에서 3분, 다시 95oC에서 30초, 58oC에서 30초, 72oC 에서 20초로 35사이클을 증폭한 후, 추가로 72oC에서 5분 간 반응시켰다. 중합효소연쇄반응 후 이 서열들을 sanger sequencing (Cosmogenetech, Seoul, Korea)을 수행하여 변이를 확인하였을 때, 표고버섯 표준유전체의 Scaffold 2 번 1803483의 염기서열 C가 ‘산마루2호’에서 T로 변한 SNP를 가장 적합한 서열로 판단하였다(Table 3). 이 서열 을 RL-LE-129로 명명하고 마커 서열로 이용하였다.
B17과 ‘산마루2호’에서 증폭한 마커서열 RL-LE-129에 존재하는 특이적인 제한효소를 dCAPS Finder 2.0 (http:/
/helix.wustl.edu/dcaps/dcaps.html)을 이용해 검색하였을 때, ‘산마루2호’에서는 제한효소 HhalⅠ의 인식부위가 존 Table 2. List of SNPs in Sanmaru 2ho
#Chr POS REF ALT Effect codon Gene Desc
Scaffold6 1212892 G A downstream_-
gene_variant c.*1551G>A Mbtps2 Membrane-bound transcription factor site-2 protease [Source:UniProtKB/Swiss-Prot Q8CHX6]
Scaffold2 3676659 G T upstream_gene_-
variant c.-1G>T GENE10847 -
Scaffold22 195159 A C missense_variant c.239A>C GENE00246 hypothetical protein Moror_1619 [Moniliophthora roreri MCA 2997] [Source:NCBI_NR ESK93613.1]
Scaffold19 119469 T A upstream_gene_-
variant c.-1T>A malA NADP-dependent malic enzyme [Source:UniProtKB/
Swiss-Prot Q6TU48]
Scaffold19 119470 G C upstream_gene_-
variant c.-1G>C malA NADP-dependent malic enzyme [Source:UniProtKB/
Swiss-Prot Q6TU48]
Scaffold7 3208246 G A missense_variant c.364G>A LE14B_LITERLEC14B protein [Source:UniProtKB/Swiss-Prot Q40153]
Scaffold7 5818871 C G upstream_gene_-
variant c.-1G>C GENE06197 hypothetical protein Moror_10090 [Moniliophthora roreri MCA 2997] [Source:NCBI_NR ESK85496.1]
Scaffold2 1803483 C T upstream_gene_-
variant c.-1C>T TOP2 DNA topoisomerase 2 [Source:UniProtKB/Swiss-Prot Q9Y8G8]
#Chr, number of chromosome; POS, position; REF, reference; ALT, alternative; Desc, description
Table 3. Characteristics of RL-LE-129 as marker for identification of Sanmaru 2ho
Primer name Primer sequence Target DNA sequence Size
(bp) RL-LE-129 F TGGGATTGCAGCATAACAAG TGGGATTGCAGCATAACAAGgactattcaaatgtctcaagcttgcgatacggacgcttaatgatc
atgactgatcaggtaagttgaatattgtcacaatatgagggggCgctgagatagaacgactaggatcacgacgga tctcatatcaaagggcttttaatcaatttcatcgaccacttttatccgtcactacttaaaatacccgacttcctcgttGA GTTCGTCACGCCGATTAT
RL-LE-129 R ATAATCGGCGTGACGAACTC 238
재하지 않았다(Fig. 1). 이를 이용하여, 38개 시험품종으 로 확대하여 검증하였을 때, ‘산마루2호’에서만 제한효소 HhalⅠ의 인식부위가 존재하지 않아 산마루2호의 PCR산 물만 절단되지 않는 것을 확인하였다(Fig. 2). 따라서 마 커서열인 RL-LE-129을 증폭하고, 제한효소 HhalⅠ을 처 리하여 ‘산마루2호’를 구분할 수 있는 CAPS 마커를 개발 하였다.
CAPS 마커는 공우성 마커로 비교적 간단한 분석을 통 해 일정한 결과를 안정적으로 얻어낼 수 있는 장점이 있 어(Konieczny and Ausubel, 1993; Kaundun and Matsumoto, 2003), 올리브, 배, 딸기와 같은 작물에서 품종을 판별하는데 사용되고 있다(Kunihisa et al., 2003; Reale et al., 2006; Moriya et al., 2007). 최근 표고버섯의 유전체 정보가 밝혀져, 이를 이용한 분자마 Fig. 1. Validation of SNP in monokaryon B17 and L. edodes cultivar Sanmaru 2ho using Sanger sequencing and, detection of restriction enzyme site for CAPS marker for identification of Sanmaru 2ho.
Fig. 2. Determination of Sanmaru 2ho by developed CAPS marker. Sequence of amplified RL-LE-129 in Sanmaru 2ho was not cleaved by restriction enzyme Hha I(box). The number shown in figure represented the given number of each cultivars listed in Table 2.
커의 개발이 증가할 것이라 생각된다, 본 연구에서는 표 고버섯의 전장유전체 정보를 바탕으로 국내에서 개발된 37개 표고버섯 품종으로부터 ‘산마루2호’의 구분이 가능 한 CAPS 마커를 개발하였다. 개발된 CAPS 마커는 산 마루2호를 다른 품종으로부터 보호하는 과학적 근거가 될 것이다.
적 요
우리나라에서 표고버섯은 소비자의 선호도가 매우 높고, 전체 임산버섯 생산량의 약 97.7%를 차지하고 있는 매우 중요한 단기소득임산물중 하나이다. 이러한 표고버섯은 최근 신품종의 개발이 활발히 이루어지고 있어, 육종가의 권리 보호를 위하여 품종을 구분할 수 있는 분자마커 개 발이 요구되고 있다. 본 연구에서는 신품종 ‘산마루2호’
를 37개의 표고버섯 품종으로부터 구분할 수 있는 CAPS 마커를 개발하였다. ‘산마루2호’의 유전체에서 Scaffold 2 번 1803483에 위치한 단일염기 다형성(SNP)이 확인되었 고, 이 SNP를 포함하여 증폭한 DNA는 제한효소 Hhal에 의하여 특이적으로 절단되지 않아 다른 표고버섯 품종들 과 구분되었다.
Acknowledgements
This study was supported by a grant Golden Seed Project (Center for Horticultural Seed Development, No.
213007-05-2-SBH20).
References
Bak WC, Park YA, Park JH. 2013. Present situation and future of oak mushroom industry: NIFos forest policy issue 11. p. 1.
Korea Forest Research Institute.
Bisen PS, Baghel RK, Sanodiya BS, Thakur GS, Prasad GB. 2010.
Lentinus edodes: A macrofungus with pharmacological activities. Curr Med Chem 17:2419-2430.
Fu LZ, Zhang HY, Wu XQ, Li HB, Wei HL, Wu QQ. 2010. Wang LA. Evaluation of genetic diversity in Lentinula edodes strains using RAPD, ISSR and SRAP markers. World J Microbl Biotech 26:709-716.
International Mycological Association. Mycobank Database Fungal Databases, Nomenclature and Species Banks.
Available online: http://www.mycobank.org/ (9 January 2018).
KATI. Available online: http://www.kati.net (12 January 2017). (in Korean)
Kaundun S, Matsumoto S. 2003. Development of CAPS markers based on three key genes of the phenylpropanoid pathway in tea, Camellia sinensis (L.) O. Kuntze, and differentiation between assamica and sinensis varieties. TAG Theoretical and Applied Genetics 106:375-383.
Konieczny A, Ausubel FM. 1993. A procedure for mapping Arabidopsis mutations using co‐dominant ecotype‐specific
PCR‐based markers. Plant J 4:403-410.
Korea Forest Service. 2008. Test guidelines for Lentinula edodes.
p. 1-29. Korea Forest Service. (in Korean)
Korea Forest Service. 2017. The Statistical Yearbook of Forestry 2017. p. 292-331. Korea Forest Service. (in Korean)
Korea Rural Economic Institute. 2016. Forestry observation for Lentinula edodes, November. p. 1-6. Korea Rural Economic Institute. (in Korean)
Kulkarni RK. 1991. DNA Polymorphisms in Lentinula edodes, the Shiitake Mushroom. App Environ Microbiol 57:1735-1739.
Kunihisa M, Fukino N, Matsumoto S. 2003. Development of cleavage amplified polymorphic sequence (CAPS) markers for identification of strawberry cultivars. Euphytica 134:209- 215.
Lee HY, Moon S, Shim D, Hong CP, Lee Y, Koo CD, Chung JW, Ryu H. 2017. Development of 44 Novel Polymorphic SSR Markers for Determination of Shiitake Mushroom (Lentinula edodes) Cultivars. Genes 8:109.
Li HB, Wu XQ, Peng HZ, Fu LZ, Wei HL, Wu QQ, Jin QY, Li N.
2008. New available SCAR markers: potentially useful in distinguishing a commercial strain of the superior type from other strains of Lentinula edodes in China. Appl Microbiol Biotechnol 81:303-309.
Liu J, Wang ZR, Li C, Bian YB, Xiao Y. 2015. Evaluating genetic diversity and constructing core collections of Chinese Lentinula edodes cultivars using ISSR and SRAP markers. J Basic Microbiol 55:749-760.
Liu JY, Ying ZH, Liu F, Liu XR, Xie BG. 2012. Evaluation of the use of SCAR markers for screening genetic diversity of Lentinula edodes strains. Curr Microbiol 64:317-325.
Moon S, Lee HY, Kim M, Ka, KH, Ko HG, Chung JW, Koo CD, Ryu H. 2017b. Development of CAPS Marker for Identification of Lentinula edodes Cultivars Sanmaru 1 and Chunjang 3. Kor J Mycol 14:114-120. (in Korean)
Moon S, Lee HY, Shim D, Kim M, Ka, KH, Ryoo R, Ko HG, Koo CD, Chung JW, Ryu H. 2017a. Development and Molecular Characterization of Novel Polymorphic Genomic DNA SSR Markers in Lentinula edodes. Mycobiology 45:105-109.
Moriya Y, Yamamoto K, Okada K, Iwanami H, Bessho H, Nakanishi T, Takasaki T. 2007. Development of a CAPS marker system for genotyping European pear cultivars harboring 17 S alleles. Plant cell reports 26:345-354.
National Institute of Forest Science. 2016. Golden Seed Mushroom Cultivation Technology. p. 68. National Institute of Forest Science. (in Korean)
Ng ML, Yap AT. 2002. Inhibition of human colon carcinoma development by lentinan from shiitake mushrooms (Lentinus edodes). J Altern Complement Med 8:581-589.
Qin LH, Tan Q, Chen MJ, Pan YJ. 2006. Use of intersimple sequence repeats markers to develop strain-specific SCAR markers for Lentinula edodes. FEMS Microbiol Lett 257:112- 116.
Reale S, Doveri S, Díaz A, Angiolillo A, Lucentini L, Pilla F, Martin A, Donini P, Lee D. 2006. SNP-based markers for discriminating olive (Olea europaea L.) cultivars. Genome 49:1193-1205.
Rop O, Mlcek J, Jurikova T. 2009. Beta-glucans in higher fungi and their health effects. Nutr Rev 67:624-631.
Royse DJ. 2014. A global perspective on the high five: Agaricus, Pleurotus, Lentinula, Auricularia & Flammulina. p. 1-6.
Proceedings of the 8th International Conference on Mushroom Biology and Mushroom Products.
Shim D, Park SG, Kim K, Bae W, Lee GW, Ha BS, Ro HS, Kim M, Ryoo R, Rhee SK, Nou IS, Koo CD, Hong CP, Ryu HJ.
2016. Whole genome de novo sequencing and genome annotation of the world popular cultivated edible mushroom, Lentinula edodes. J Biotechnol 223:24-25.
Terashima K, Matsumoto T. 2004. Strain typing of shiitake (Lentinula edodes) cultivars by AFLP analysis, focusing on a heat-dried fruiting body. Mycoscience 45:79-82.
Wu X, Li H, Zhao W, Fu L, Peng H, He L, Cheng J. 2010. SCAR makers and multiplex PCR-based rapid molecular typing of
Lentinula edodes strains. Curr Microbiol 61:381-389.
Xiao Y, Liu W, Dai Y, Fu C, Bian Y. 2010. Using SSR markers to evaluate the genetic diversity of Lentinula edodes’ natural germplasm in China. World J Microbl Biotech 26:527-536.
Zhang R, Huang C, Zheng S, Zhang J, Ng TB, Jiang R, Zuo X, Wang H. 2007. Strain-typing of Lentinula edodes in China with inter simple sequence repeat markers. Appl Microbiol Biotechnol 74:140-145.
Zhang Y, Molina FI. 1995. Strain typing of Lentinula edodes by random amplified polymorphic DNA assay. FEMS Microbiol Lett 131:17-20.