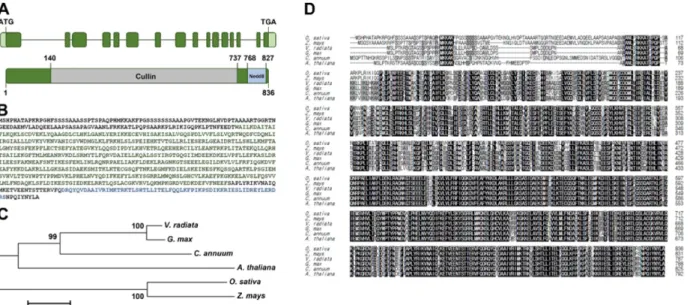
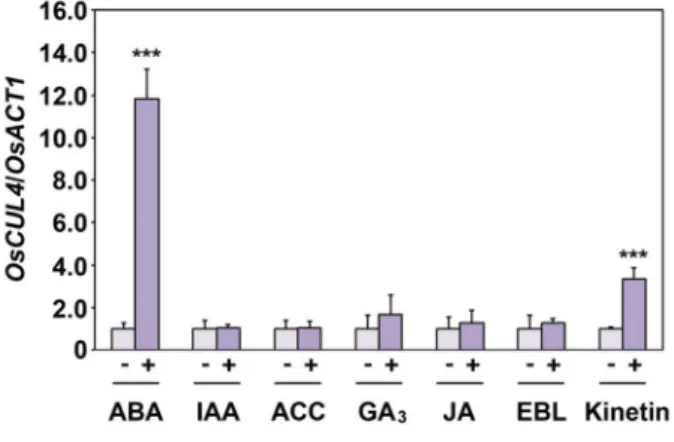
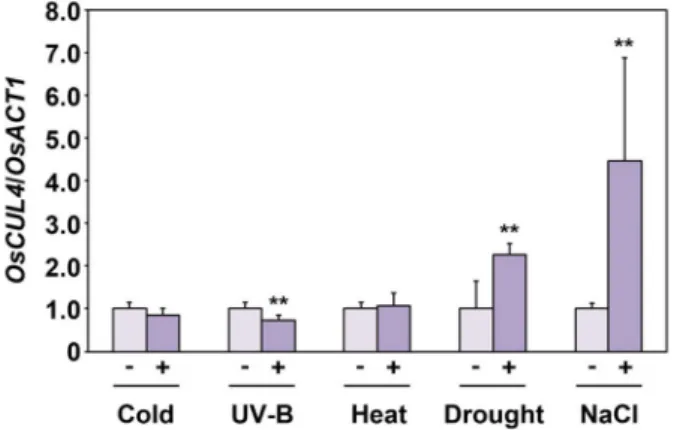
Expression Study on the Scaffold Gene of CRL4 Complex in Rice (Oryza sativa L.)
Yoowon Bae
1, Hani Kim
1,2, Sang-Hoon Kim
1and Jae-Hoon Lee
1*
1
Department of Biology Education, Pusan National University, Busan 46241, Korea
2
전체 글
1
2
수치

관련 문서
Ross: As my lawfully wedded wife, in sickness and in health, until
다양한 번역 작품과 번역에 관한 책을 읽는 것은 단순히 다른 시대와 언어, 문화의 교류를 넘어 지구촌이 서로 이해하고 하나가
The index is calculated with the latest 5-year auction data of 400 selected Classic, Modern, and Contemporary Chinese painting artists from major auction houses..
“Challenges and Opportunities for Enhancing Rice Production in Nepal.” Rice Science and Technology in Nepal.. Crop Development Directorate and Agronomy Society
Distribution of Cd in stream sediments, paddy soils, rice stalks and grains from the Kubong
To learn about the necessity of textile art education and tapestry technique, to study the value of tapestry education and to utilize it in art education
This study aims to increase the understanding of textbooks, analyzing the contents that the singing/instrumental music activities in the Expression Area in
Results : The expression of p21 was increased in boderline serous tumor and serous cystadenocarcinoma in contrast to benign serous tumors. The expression of
