© Copyright
Keimyung University School of Medicine 2019
Differentiation of preadipocyte, also named adipogenesis, leads to the phenotype of mature adipocyte that is filled with many lipid droplets.
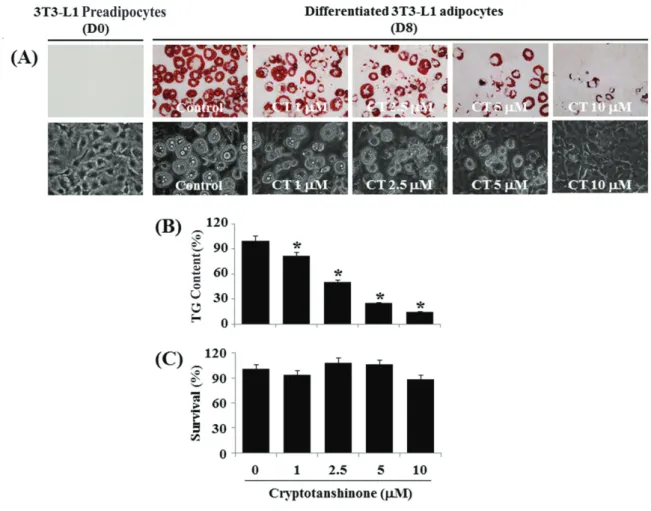
Excessive lipid accumulation in adipocytes leads to the development of obesity. In this study, we investigated the effect of 11 different natural compounds on lipid accumulation during the differentiation of 3T3-L1 preadipocytes into 3T3-L1 adipocytes. Strikingly, among the natural compounds, cryptotanshinone at 10 μM most strongly reduced triglyceride (TG) contents in 3T3-L1 cells after 8 days of the differentiation.
Furthermore, cryptotanshinone at 10 μM significantly suppressed lipid accumulation in 3T3-L1 cells after 8 days of the differentiation.
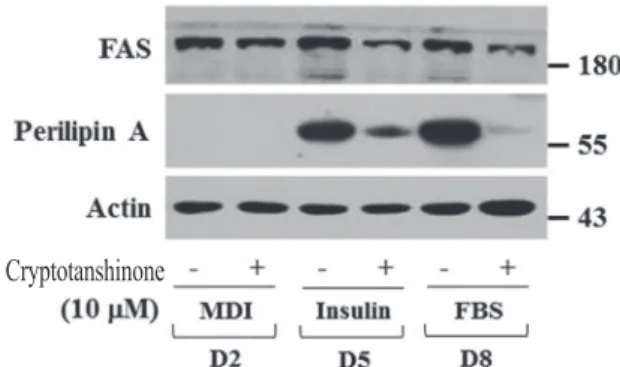
Cryptotanshinone at 1 to 10 μM tested did not affect the survival of 3T3-L1 cells after 8 days of the differentiation. On mechanistic levels, cryptotanshinone time-differentially decreased the expression levels of CCAAT/enhancer-binding protein-α (C/EBP-α), peroxisome proliferator- activated receptor-γ (PPAR-γ), fatty acid synthase (FAS), and perilipin A but also the phosphorylation levels of signal transducer and activator of transcription-3 (STAT-3) during the 3T3-L1 cell differentiation. Taken together, these findings demonstrate that cryptotanshinone inhibits lipid accumulation in differentiating 3T3-L1 cells, which appears to be mediated through the reduced expression and/or phosphorylation levels of C/EBP-α, PPAR-γ, FAS, Perilipin A, and STAT-3.
Keywords: Cryptotanshinone, CCAAT/enhancer-binding protein-α, Perilipin A, Perilipin A transcription-3
Received: October 15, 2019 Revised: October 22, 2019 Accepted: October 30, 2019
Corresponding Author: Byeong Churl Jang, Department of Molecular Medicine, Keimyung University School of Medicine, 1095 Dalgubeol-daero, Dalseo-gu, Daegu 42601, Korea
Tel: +82-53-258-7404
E-mail: [email protected] (B-C Jang)
• The authors report no conflict of interest in this work.