359
책임저자:정상영, 광주시 동구 학1동 8번지
501-757, 전남대학교병원 외과 Tel: 062-220-6472, 6456, Fax: 062-227-1635 E-mail: [email protected]
접수일:2007년 7월 16일, 게재승인일:2007년 8월 29일
Tacrolimus (FK506)에 의해 유도된 세포 고사의 신호 전달 기전에 대한 연구
전남대학교 의과대학 외과학교실 정미란ㆍ최수진나ㆍ정상영
Tacrolimus (FK506) Induced Apoptotic Signal Transduction Pathway
Mi Ran Jung, M.D., Soo Jin Na Choi, M.D. and Sang Young Chung, M.D.
Purpose: This study examined the effects of Tacrolimus (FK506) on the expression of the apoptotic signal trans- duction proteins of Jurkat human T-lymphocytes.
Methods: The cell viability was examined by a MTT assay, DAPI stain, enzyme activity of caspase family proteins, and western blotting for Bcl-2, Bak, Fas, and Fas-L. The cells were cultured in the presence or absence of FK506. FK506 induced cell death was confirmed to be apoptosis by the observation of nuclear fragmentation.
Results: The viability of Jurkat cells was decreased by the addition of FK506 in a dose- and time- dependent manner.
The FK506 induced activation of caspase-3 protease was observed. FK506 didn't increase the catalytic activity of cas- pase -6, -8, and -9 proteases of Jurkat cells in a time-de- pendent manner. The viability was improved when a cas- pase-3 inhibitor was added. However, the caspase-9 inhibitor did not affect the viability. Bak protein expression was in- creased, and the Bcl-2 protein was decreased for some time.
The expression of Fas and Fas-L were unaffected by FK506.
Conclusion: FK506 induces dose- and time-dependent apop- totic cell death, and enhances the apoptosis of Jurkat cell by increasing the expression of Bak and caspase-3. (J Korean Surg Soc 2007;73:359-3)
Key Words: Tacrolimus (FK506), Jurkat cell, Apoptosis, Bak protein, Caspase-3
중심 단어: Tacrolimus (FK506), Jurkat 세포, 세포고 사, Bak 단백질, Caspase-3
Department of Surgery, Chonnam National University College of Medicine, Gwangju, Korea
서 론
FK506 (Tacrolimus)은 T 림프구의 신호 전달 과정을 억제 하는 면역억제제로 토양진균인 Streptomyces tsukubaensis에 서 얻어지는 macrolide계 항생물질이다. FK506은 1982년 일 본에서 처음 개발되었으며, 1990년 미국 피츠버그대학 이 식팀이 간이식에 최초로 임상 적용을 하였고 그 성공적인 결과를 바탕으로 현재 전세계적으로 광범위하게 사용되는 면역억제제 중의 하나가 되었다.(1)
현재 면역억제요법의 근간을 이루는 FK506과 사이클로스 포린(Cyclosporin)은 모두 Calcineurin 억제제로서 Interleukin-2 (IL-2)의 발현을 억제함으로써 T 림프구의 활성화를 억제한 다.(2) 특히 FK506은 각종 시험관 내 시험에서 그 면역 억제 기능이 사이클로스포린보다 10∼200배 강력한 것으로 보고 되어 있으며, 거부반응의 예방은 물론 난치성 거부반응의 구조요법으로도 효과적인 것으로 알려져 있다.(3)
FK506은 T 림프구의 세포질 내에서 FK-Binding Protein (FKBP)과 복합체를 형성하고 이 복합체가 세포질의 calm- odulin-calcineurin복합체에 결합하여 T 림프구 활성화와 관 련된 IL-2 유전자의 전사를 억제하며, IL-3, IL-4, GM-CSF, TNF-α 등 T 림프구의 초기 활성 단계에 관련되는 사이토 카인(cytokine)들의 유전자 전사를 저해하는 것으로 알려져 있다.(4-6) 그러나, FK506이 세포고사의 신호 전달 체계에 미치는 영향 및 기전에 대한 것은 잘 알려져 있지 않다. 이 에 저자들은 사람 T 림프구를 이용하여 FK506이 세포 고사 의 신호 전달 기전에 유의한 영향을 미치는 결과를 얻었기 에 보고하는 바이다.
방 법 1) 재료
(1) 세포주: 사람 T 림프구 세포주인 Jurkat 세포는 한국 세
포주 은행에서 구입, 계대 배양하면서 실험을 실시하였다.
(2) 시약 및 기기: 실험에 필요한 RPMI 1640 배지, 소태아 혈청(Fetal bovine serum, FBS)은 GIBCO BRL사(Grand Island, NY, USA)에서, 배양 용기(24-well plate와 10cm dish) 는 Falcon사(Becton Dickinson, San Jose, CA, USA)에서 구입 하여 사용하였다. 3-[4,5- dimethylthiazol-2-yl]-2,5-diphenyl tet- razolium bromide (MTT), DAPI, dimethyl sulfoxide (DMSO), N-acetyl-L-cysteine (NAC)은 Sigma사(St. Louis, Missouri, USA)에서 구입하였다.
2) 방법
(1) Jurkat 세포주 배양 및 시약 처리: Jurkat 세포는 CO2
세포 배양기(37oC, 5% CO2)에서 10% 소태아 혈청이 포함된 RPMI 1640 배지로 배양하였으며, 24시간 간격으로 배양액 을 교체하여 log phase에 있는 세포에 FK506을 처리한 후 세포 고사 현상과 이에 연관된 생화학 실험을 수행하였다.
FK506은 10μM 농도로 DMSO에 녹여 -70oC에 보관하였 고, RPMI 1640 배지에 희석하여 사용하였다.
(2) 세포활성도 측정: 세포의 활성도 측정은 MTT 분석법 으로 측정하였다. 세포 배양판(24-well plate)에 세포(1×105 세포수/ml)를 1 ml씩 분주하여 3시간 이상 CO2 세포 배양기 안에서 안정시킨 후 시료를 각각의 조건에 따라 처리하였 다. MTT 용액(5 mg/ml, phosphate buffered salin: PBS, pH 7.4) 각각의 배양 세포에 배양액의 1/10을 첨가하여 4시간 반응 하였다. 살아있는 세포에 의해 생성된 보라색 formazan은 10% SDS가 포함된 0.01N HCl 용액을 1 ml/well 첨가하여 용해시킨 후, 분광 광도계(THERMO max, USA)를 이용하여 540 nm 파장에서 흡광도를 측정하였다. 세포 활성도는 정 상 대조군과 비교하여 백분율(%)로 표시하였다.
(3) DAPI에 의한 세포핵 염색: FK506으로 처리한 세포는 포집하여 냉 PBS로 세척한 후 cytospin (Shandon Southern Products Ltd., England)으로 600 rpm, 3분간 회전 분사하여 슬라이드 글라스에 부착시켰다. 슬라이드 글라스에 부착된 세포는 4% formaldehyde 용액으로 고정하고 PBS로 세척하 여, 10μM DAPI로 20분간 실온에서 반응한 후 다시 PBS로 세척하여 형광 현미경(Leica MPS 60, Germany)으로 핵 형태 를 관찰하였다.
(4) Caspase계 cysteine protease 활성도 측정: FK506 처리 후 포집한 Jurkat 세포(1×106)를 4oC에서 15분간 lysis buffer (1% Triton X-100, 0.32 M sucrose, 10 mM Tris/HCl, pH 8.0, 5 mM EDTA, 2 mM DTT, 1mM PMSF, 1μg/ml aprotinin, 1μ g/ml leupeptin)으로 용해하고 13,000 rpm으로 15분간 원심 분리하였다. 원심 분리하여 얻은 세포 파쇄액은 bicincho- ninic acid (BCA, Sigma Co. MO, U.S.A.)법으로 단백질을 정 량하고, 세포 파쇄액을 분석 완충용액(100mM HEPES, pH 7.5, 10% sucrose, 1.0% Chaps, 2 mM DTT, 1 mM PMSF, 1 μg/ml aprotinin, 1μg/ml leupeptin)에 희석된 형광 기질과
37oC에서 30분간 반응시켰다. Caspase protease의 활성 측정 은 fluorogenic substrate를 사용하였으며, 이 기질의 proteo- lytic cleavage에 의하여 생성되어지는 형광값의 차이를 형 광 분광 광도계(Molecular Devices Co, USA)로 측정하여 caspase 활성을 결정하였다.
(5) Western blotting: FK506을 처리한 Jurkat 세포는 포집 하여, 냉 PBS로 2회 세척하였다. 얻어진 세포는 파쇄 용액 (50 mM HEPES, pH7.4, 150mM NaCl, 1% deoxycholate, 1mM EDTA, 1 mM PMSF, 1μg/ml aprotinin)과 4oC에서 30분간 반 응시켰다. 동량의 세포 파쇄액(200μg)과 2X sample buffer 를 혼합하여 100oC에서 3분간 가열하여 단백질의 변성을 유도한 후에 12.5% 및 15% SDS-polyacrylamide gel electro- phoresis (SDS-PAGE)를 시행하였다. 전기영동이 끝난 gel의 단백질은 electrotransfer system (Ellard Inc, Seattle, WA, USA)을 이용(0.8 mA/cm)하여 nitrocellulose membrane으로 이동시키고, blocking buffer (5% skim milk)와 상온에서 2시 간 동안 반응시켰다. Bcl-2, Bak, Fas 및 Fas-L에 대한 항체는 0.05% (v/v)의 tween-20이 함유된 Tris-buffered saline (TBS- T)에 1:1,000으로 희석하여 nitrocellulose membrane과 상온 에서 2시간 반응시켰으며, 각 항체에 대한 이차항체 an- ti-rabbit IgG conjugated horse radish peroxidase (HRP)와 an- ti-mouse IgG conjugated HRP는 TBS-T로 희석(1:3,000)하여 상온에서 1시간 반응시킨 후, enhanced chemilluminescence (ECL) kit (Amersham, England)를 이용하여 현상하였다.
(6) 통계 처리: 표시된 결과는 3번 이상의 독립적인 실험 결과이며, 실험 결과의 통계 처리는 Student's t-test에 준하 여 처리하였고, P-value가 0.05 미만인 경우를 유의한 것으 로 판정하였다.
결 과
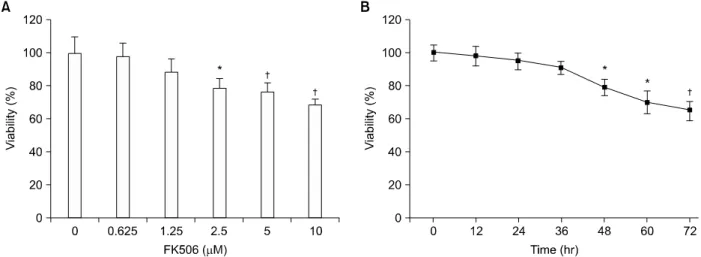
1) FK506이 Jurkat 세포의 생존율에 미치는 영향 FK506의 Jurkat 세포의 생존율에 대한 영향을 조사하기 위하여 FK506을 다양한 농도로 72시간 처리한 후, 세포 활 성도를 MTT 방법으로 조사하였다. FK506 0.625μM농도에 서 세포생존율은 96%, 1.25μM농도에서는 89%, 2.5μM 농 도에서는 77%, 5μM 농도에서는 76%의 세포 생존율을 보 였으며, 10μM의 농도에서는 대조군에 비교하여 64%의 세 포 생존율을 보였다(Fig. 1A). 또한, FK506에 의한 Jurkat 세 포 생존율의 시간 의존적인 변화를 확인하기 위하여 10μM 농도의 FK506을 처리한 후 시간별로 세포 생존율을 조사하 였다. Jurkat 세포의 생존율은 FK506 처리 12시간 후부터 감 소하기 시작하여, 72시간 이후에는 약 65%로 유의한 세포 생존율의 감소를 보였다(Fig. 1B). 이상의 결과는 FK506에 의한 Jurkat 세포의 생존율이 농도 및 시간 의존적으로 감소 함을 보여 주었다.
Fig. 1. FK506 decreased the viability of Jurkat cells in dose- and time-dependent manners. Cells were treated with various concentrations of FK506 for 72 hr (A) or various periods (B) and then, viability was measured by MTT assay after FK506 treatment. Data represent the mean±standard deviation (S.D.) of quadruplicates. *P<0.05, †P<0.01 by Student's t-test, compared to control group.
Fig. 2. FK506 induced the morpho- logical change in Jurkat cells in a time-dependent manner. Cells were treated with FK506 (10 μM) for various periods. Then, cells were stained with DAPI and observed under fluorescence microscopy. (A) control cells, (B) FK506 treated cells for 24 hr, (C) 48 hr and (D) 72 hr.
2) FK506에 의한 Jurkat 세포의 세포 죽음 성격 규명 FK506에 의한 세포 죽음이 세포 고사 기전에 의하여 매개 됨이 제시되었기 때문에 세포 고사 현상의 형태학적 특징
을 관찰하기 위하여 핵산의 DAPI 염색을 시행하였다.
Jurkat 세포에 FK506 (10μM)을 각각 24시간, 48시간 및 72 시간 처리한 후 DAPI 염색을 시행한 결과, 정상 대조군의 핵은 타원형의 온전한 형태의 균질한 형광 염색을 나타냈
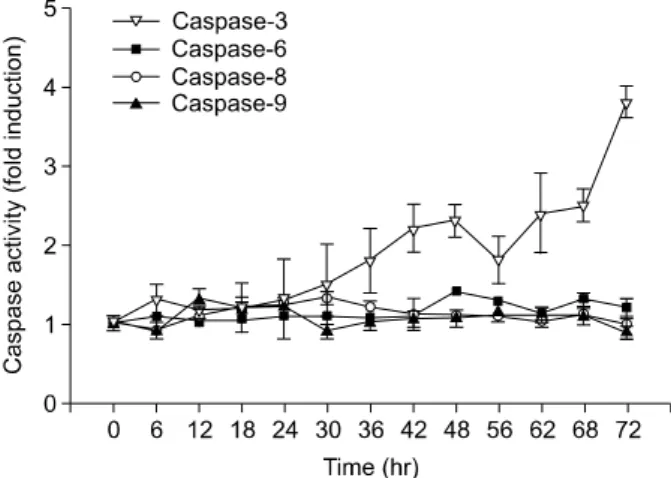
Fig. 3. FK506 did not increase the catalytic activity of caspase -6, -8, and -9 proteases of Jurkat cells in a time-dependent manner. But, it increased the catalytic activity of caspase-3 protease of Jurkat cells in a time-dependent manner. Cells were treated with 10μM FK506 for various periods and lysed to measure the activity of caspase proteases by using fluorogenic biosubstrates. Data represent the mean±S.D. of quadraplicates. Only caspase-3 shows statistical signifi- cance.
Fig. 4. When the caspase-3 inhibitor was added, the viability was improved. But the caspase-9 inhibitor did not affect viability. Cells were treated with 10μM FK506, 10μM FK506 with 30mM DEVD-AMC (caspase-3 inhibitor), 10 μM FK506 with 100 mM YVAD-AMC (caspase-9 in- hibitor), 30 mM DEVD-AMC only, and 100mM YVAD- AMC (caspase-9 inhibitor) only for 72hrs, and then, via- bility was measured by MTT assay.
Fig. 5. Expression of Bcl-2 and Bak proteins in FK506 treated Jurkat cells. Cells were treated with 10μM FK506 for var- ious periods. The equal amount of protein from cell lysate was subjected on 15% SDS-PAGE, transferred onto nitro- cellulose membrane and immunoblotted with anti-Bcl-2, anti-Bak and anti-β-actin antibodies. The immunoreactive signals were visualized by ECL detection kit. Bak protein expression was increased, and Bcl-2 protein was decreased for a while.
으나(Fig. 2A), FK506 72시간 처리군의 핵은 여러 조각으로 분절된 형광 절편이 증가되어 관찰되었다(Fig. 2D).
3) FK506의 caspase family proteins의 활성에 미치는 영향
세포 고사의 특징적인 현상을 초래하는 세포 내 신호 전
달 기전에서 caspase의 중요성이 이미 잘 알려져 있다.
FK506에 의한 Jurkat 세포의 고사 현상이 caspase 활성화와 관계가 있는지를 확인하기 위하여 caspase-3, -6, -8, 및 -9 proteases의 효소적 활성을 이들 효소의 형광 기질 (fluorogenic substrate)을 이용하여 조사하였다. Jurkat 세포는 FK506 (10μM)으로 처리하였으며 12시간 간격으로 caspase protease의 활성을 측정하였다. 그 결과 Caspase-6, -8, 및 -9 protease의 활성은 대조군과 비교해 유의한 변화가 없었으 나, caspase-3 protease는 시간 의존적으로 증가하여 FK506 처리 72시간 후에는 대조군에 비하여 최대 3.8배의 증가된 활성을 보였다(Fig. 3). Caspase-3, -9 inhibitor를 추가하였을 때 caspase-3 inhibitor는 세포생존율 향상 효과를 보였으나 caspase-9 inhibitor를 첨가하였을 때에는 생존율 변화가 없 었다(Fig. 4).
4) FK506이 Bcl-2 및 Bak 단백질의 발현에 미치는 영향 FK506 처리 시 Bcl-2 및 Bak 단백질 발현을 알아보기 위하 여 Jurkat 세포에 10μM의 FK506을 처리한 후 Western blot 을 실시하였다. 그 결과, FK506은 시간 의존적으로 pro- apoptotic 기능을 갖는 Bak 단백질의 발현을 증가시켰지만, anti-apoptotic 기능을 갖는 Bcl-2 단백질은 FK506 처리 24, 48시간 후에 약간 감소하였으나 72시간 후에는 증가되는 발현 양상을 보였다(Fig. 5). 이때 단백질양은 β-actin을 통 하여 동일한 양임을 확인하였다.
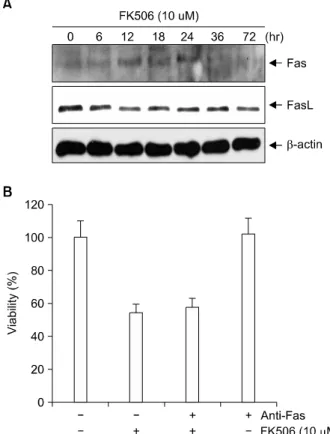
5) FK506이 Fas 및 Fas-L 단백질의 발현에 미치는 영향 FK506 처리 시 Fas 및 Fas-L 단백질 발현을 알아보기 위 하여 Jurkat 세포에 10μM의 FK506을 처리한 후 Western blot을 실시하였다. Fas와 Fas-L 단백질 발현은 FK506 처리 24, 48, 72시간이 지나도 변화되는 양상을 보이지 않았다.
이때 단백질양은 β-actin을 통하여 동일한 양임을 확인하였
Fig. 6. Expression of Fas and Fas-L proteins in FK506 treated Jurkat cells. Cells were treated with 10μM FK506 for var- ious periods. The equal amount of protein from cell lysate was subjected on 15% SDS-PAGE, transferred onto nitro- cellulose membrane and immunoblotted with anti-Fas, an- ti-Fas-L and anti-β-actin antibodies. The immunoreactive signals were visualized by ECL detection kit. There were no expressions of Fas and Fas-L (A). When Fas inhibitor was added, the viability was not improved (B).
다(Fig. 6A). 또한 anti- Fas항체를 첨가하여도 세포 생존율에 영향을 주지 못하는 것을 확인할 수 있었다(Fig. 6B).
고 찰
FK506은 토양진균인 Streptomyces tsukubaensis에서 얻어 지는 macrolide계 항생물질이다. 이는 1982년 일본에서 개 발되어 1990년 미국 피츠버그대학 이식팀이 간이식에 최초 로 임상 적용을 하였고, 그 성공적인 결과를 바탕으로 현재 는 전세계에 걸쳐 널리 사용되고 있다.(1) 특히, 간이식환자 에서 FK506이 사이클로스포린에 비해 간세포 보호 기능이 우월하다는 연구결과가 발표되어(7) 현재는 간이식 환자의 과반수 이상이 FK506을 복용하고 있으며, 신장과 간이식뿐 만 아니라 췌장, 심장, 골수이식에서도 사용되는 등 FK506 의 점유율이 점점 확대되고 있다.(8,9)
사이클로스포린 A와 FK506은 상이한 구조를 가지지만
그 작용 기전은 유사한 것으로 알려져 있다. 이 두 약제는 세포 내에서 immunophilin이라는 수용체와 결합하는데, 사 이클로스포린 A은 cycliophilin과 결합하고, FK506의 경우에 는 FK-binding protein (FKBP)와 결합한다.(4-6) 이 세포내 수 용체들은 peptidyl-prolyl isomerase 활성을 가지고 있어서 세 포 내에서 단백질 샤페론으로 작용한다. 즉 약제와 im- munophilin의 복합체는 calcineurin과 결합하여 calcineurin의 기질인 nuclear factor of activated T cells (NF-AT)의 탈인산 화를 억제함으로서, NF-AT에 의한 각종 염증 유발성 사이 토카인의 전사를 억제함이 보고되었다.(4,5) 그러나, FK506 에 의한 T림프구의 세포고사에 대한 신호 전달 과정 및 작 용 기전에 대한 연구는 미흡한 실정이다. 따라서 본 연구에 서는 사람 T 림프구인 Jurkat 세포에 대한 FK506의 세포 고 사 유도 현상과 신호 전달 기전을 조사하고자 하였다.
세포의 죽음은 크게 괴사(Necrosis)와 고사(Apoptosis)로 구분할 수 있다. 세포괴사는 세포막의 파괴로 물과 전해질 이 세포 내로 유입되어 미토콘드리아 등 세포 내 구조물이 팽창하여 파괴되는 특징을 가지고 있다. 이에 반하여 세포 고사는 빠른 탈수 현상에 의한 세포의 수축, 세포막의 기포 화 현상, 세포질 내의 칼슘농도의 증가, chromatin의 응축, endonuclease의 활성화에 의한 DNA의 분절형성 및 핵의 절 단과 세포고사 소체(Apoptotic body)의 형성을 특징으로 한 다.(10) 본 연구에서 FK506을 첨가한 Jurkat 세포의 형태학 적 변화에서 이러한 세포고사의 특징적 형태들을 확인함으 로써 FK506이 세포고사를 촉진함을 확인하였다.
최근 caspase의 작용과 관련된 단계적인 반응 기전이 밝 혀지면서 세포 고사와 관련된 연구가 활발히 이루어지고 있다. 세포고사의 기전은 크게 사망-수용체 경로(Death re- ceptor pathway)와 사립체 경로(Mitochondrial pathway)로 나 누어 설명되는데, 두 경로에서 모두 caspase-3의 활성화가 중요한 기전으로 기여하고 있다. 본 연구는 이 두 경로와 관련된 항목들의 발현정도에 FK506이 어떤 영향을 미치는 가를 알아보고 이를 통해 FK506이 세포고사에 어떻게 작용 하는지 알기 위해 본 연구를 고안하였다.
세포고사 신호전달체계에서 중요성이 잘 알려진 caspase family cysteine proteases는 현재 14종류가 알려져 있다.
Caspase는 세포고사에 중요한 역할을 할 뿐만 아니라 우리 에게 직, 간접적으로 세포고사가 일어나고 있음을 알게 해 준다. Caspase 분석을 통해 세포가 활성화된 caspase를 포함 하는지를 검사하거나 활성화된 caspase에 대한 항체를 검사 함으로써 세포고사의 진행여부를 판단할 수 있으며, DNA 분절화 등 쉽게 탐지할 수 있는 세포변화를 일으켜 세포고 사의 진행여부를 알 수 있게 해 준다.(11) Caspase는 화학구 조와 활성화 방법에 따라 크게 두 종류로 나눌 수 있다. 세 포질내에 주로 비활성형태인 이중체(Dimer)로 존재하다가, 분열(Cleavage)에 의해 활성화되어 여러 기질 단백질들을 절단하여 세포고사 현상을 일으키는 실행 caspase (Execu-
tioner caspases)와 이러한 실행 caspases를 활성화시키는 초 기화 caspases (Initiator caspases)로 구분한다. 실행 caspases 에는 caspase-3, -6, -7이 포함된다. 초기화 caspases는 실행 caspases와는 달리 비활성화 형태인 단일체(Monomer)로 존 재하다가 이중체화(Dimerization)되면서 활성화되며, 사망- 수용체 경로에서는 caspase-8이, 사립체 경로에서는 cas- pase-9이 중요한 역할을 한다.(12) 저자들은 이러한 이론을 바탕으로 실행 caspase인 caspase-3, -6, 초기화 caspase인 cas- pase-8, -9의 영향을 실험을 통해 확인하였으며, 그 결과 cas- pase-3만이 FK506과 시간의존적인 상관관계를 보였다.
사망 수용체는 종양괴사인자(Tumor necrosis factor; TNF) 수용체의 한 부류로, TNFR1, Fas, DR3/TNF receptor apopto- sis-mediating protein, TNF-related apoptosis-inducing ligand re- ceptor등이 있다. 이들은 삼중체(Trimer)의 형태로 존재하며, 사망 리간드(Death ligand)가 결합하면 사망수용체의 사망 영역(Death domain; DD)과 세포 내의 Fas-associated death domain (FADD) 간의 결합이 일어나며, FADD의 다른 영역 인 death effector domain (DED)는 비활성 형태의 초기화 cas- pases가 가진 DED와 결합함으로써 초기화 caspases를 이중 체화(dimerization)하여 활성화시킨다. 이러한 사망수용체, FADD, caspase의 복합체를 사망유도 신호 복합체(Death-in- ducing signaling complex)라고 하며, 이 활성화된 복합체가 caspases-3를 활성화시킨다.(11) 본 실험에서 FK506이 cas- pase-3의 활성도를 증가시킴이 확인되었으며, 저자들은 사 망 수용체 경로에 FK506이 어떤 영향을 미치는지 확인하고 자 FK506과 사망수용체인 Fas, Fas 리간드의 활성화에 대해 조사하였으나 상관관계는 없었다.
사립체 경로는 cytochrome c의 방출에 의해 시작한다. 정 상적으로, cytochrome c는 사립체 내에서 존재하지만, 세포 고사 중에는 사립체 외막 투과화(Mitochondrial outer mem- brane permeabilization; MOMP)가 발생하여 사립체 외로 방 출된다.(13) 방출된 cytochrome c는 매개 분자(Adoptor mole- cule)인 apoptotic protease-activating factor-1 (APAF-1)의 결합 을 유도한다. APAF-1는 정상세포의 세포질에서는 단일체 (monomer)형태로 존재하지만 cytochrome c에 의해 다중체 를 형성하여 거대분자인 apoptosome이 되면서 활성화되며 다중체를 형성하는 과정에서 APAF-1의 caspase recruitement domain (CARD)이 노출되고 이는 초기화 caspase인 cas- pase-9의 CARD와 결합하여 caspase-9의 이중체화(dimeriza- tion)을 유도함으로써 caspase-9을 활성화한다.(11) 이렇게 활성화된 caspase-9은 실행 caspase인 caspase-3를 활성화한 다. 사립체 경로의 시작점으로 볼 수 있는 MOMP는 Bcl-2 계열에 의해 조절된다.(13,14) Bcl-2 계열 중 anti-apoptotic protein으로는 Bcl-2, Bcl-xL, A1, Bcl-W, Mcl-1 등이 있으며, 이들은 MOMP를 저해함으로써 세포고사를 방지한다. 이와 반대로 MOMP를 촉진하는 pro-apoptotic proteins로는 MOMP를 직접 촉진하는 Bax, Bak, Bok 등이 있으며,(15,16)
이 pro-apoptotic proteins를 활성화함으로써 세포고사를 촉 진하는 단백으로 Bim, Bid, Bad 등이 있다.(17) 저자들은 FK506이 사립체 경로에 미치는 영향을 확인하고자 하였으 며, 그 방법으로 FK506과 Bak, Bcl-2 단백의 관계를 조사하 였다. 그 결과 Bak 단백 발현이 시간의존적으로 증가하는 것과 48시간까지 Bcl 단백 발현이 점차 감소함을 확인할 수 있었다. 그러나 caspase-9의 활성과는 상관관계를 보이지 않 았다. 이러한 결과로 미루어 FK506은 사립체 외막 투과성 의 변화를 통해 세포고사를 촉진하는 것으로 생각한다.
FKBP38은 다른 FKBP 군과는 달리 FK506과의 결합력과 calcineurin에 대한 길항 작용이 약하며, Bcl-2 단백이 사립체 막에 위치하는 것을 돕는데,(18) Kang 등(19)은 FKBP38을 억제하였을 때 Bcl-2가 불안정해지며 세포고사를 유도함을 보고하였다. 본 연구에서 72시간 후 Bcl-2가 증가하는 결과 와 Kang 등의 연구결과를 연관지어 추정해 볼 때 72시간 후 FKBP38의 발현에 의한 Bcl-2단백증가가 일어났을 가능 성이 있다. 그러나 이에 관해서는 더 많은 연구가 시행되어 야 기전을 명확히 알 수 있을 것이다.
결 론
결론적으로 FK 506은 FK506 결합 단백(FK506 binding protein; FKBP)과 결합하여 Bak 단백 발현을 증가시켜 사립 체 경로를 촉진하였을 것이며, 그 결과 caspase-3의 활성이 증가되어 세포고사를 촉진한 것으로 추정된다.
REFERENCES
1) Mayer AD, Dmitrewski J, Squifflet JP, Besse T, Grabensee B, Klein B, et al. Multicenter randomized trial comparing ta- crolimus (FK506) and cyclosporine in the prevention of renal allograft rejection: a report of the European Tacrolimus Multicenter Renal Study Group. Transplantation 1997;64:436- 43.
2) Goto T, Kino T, Hatanaka M, Okuhara M, Kohasaka M, Aoki H, et al. FK506: historical perspectives. Transplant Proc 1991;23:2713-7.
3) Jordan ML, Narachi R, Shaporo R, Smith D, Vivas CA, Scantlebury VP, et al. Tacrolimus rescue therapy for renal alograft rejection-five year experience. Transplantation 1997;
63:223-8.
4) Liu J, Farmer JD Jr, Lane WS, Friedman J, Weissman I, Schreiber SL. Calcineurin is a common target of cyclo- philin-cyclosporin A and FKBP-FK506 complexes. Cell 1991;66:807-15.
5) Clipstone NA, Crabtree GR. Identification of calcineurin as a key signalling enzyme in T-lymphocyte activation. Nature 1992;357:695-7.
6) O'Keefe SJ, Tamura J, Kincaid RL, Tocci MJ, O'Neill EA.
FK-506- and CsA-sensitive activation of the interleukin-2 pro- moter by calcineurin. Nature 1992;357:692-4.
7) Gómez-Lechon MJ, Serralta A, Donato MT, Jimenez N, O'connor E, Castell JV, et al. The immunosuppressant drug FK506 prevents Fas-induced apoptosis in human hepatocytes.
Biochem Pharmacol 2004;68:2427-33.
8) Meier-Kriesche HU, Baliga R, Kaplan B. Decreased renal function is a strong risk factor for cardiovascular death after renal transplantation. Transplantation 2003;75:1291-5.
9) Pirsch JD, Miller J, Deierhoi MH, Vincenti F, Filo RS. A comparison of tacrolimus (FK506) and cyclosporine for im- munosuppression after cadaveric renal transplantation. FK506 Kidney Transplant Study Group. Transplantation 1997;63:977- 83.
10) Wyllie AH, Kerr JF, Currie AR. Cell death: the significance of apoptosis. Int Rev Cytol 1980;68:251-306.
11) Green DR. Overview: apoptotic signaling pathways in the im- mune system. Immunological Rev 2003;193:5-9.
12) Boatright KM, Renatus M, Scott FL, Sperandio S, Shin H, Pedersen IM, et al. A unified model for apical caspase activation. Mol Cell 2003;11:529-41.
13) Kluck RM, Bossy-Wetzel E, Green DR, Newmeyer DD. The release of cytochrome c from mitochondria: a primary site for
Bcl-2 regulation of apoptosis. Science 1997;275:1132-6.
14) Cory S, Adams JM. The Bcl2 family: regulators of the cellular life-or-death switch. Nat Rev Cancer 2002;2:647-56.
15) Wei MC, Zong WX, Cheng EH, Lindsten T, Panoutsakopoulou V, Ross AJ. Proapoptotic BAX and BAK: a requisite gateway to mitochondrial dysfunction and death. Science 2001;292:
727-30.
16) Kuwana T, Mackey MR, Perkins G, Ellisman MH, Latterich M, Schneiter R, et al. Bid, Bax, and lipids cooperate to form supramolecular openings in the outer mitochondrial membrane.
Cell 2002;111:331-42.
17) Puthalakath H, Strasser A. Keeping killers on a tight leash:
transcriptional and post-translational control of the pro-apop- totic activity of BH3-only proteins. Cell Death Differ 2002;9:505-12.
18) Shirane M, Nakayama KI. Inherent calcineurin inhibitor FKBP38 targets Bcl-2 to mitochondria and inhibits apoptosis.
Nat Cell Biol 2003;5:28-37.
19) Kang CB, Feng L, Chia J, Yoon HS. Molecular character- ization of FK-506 binding protein 38 and its potential regu- latory role on the anti-apoptotic protein Bcl-2. Biochem Biophys Res Commun 2005;337:30-8.