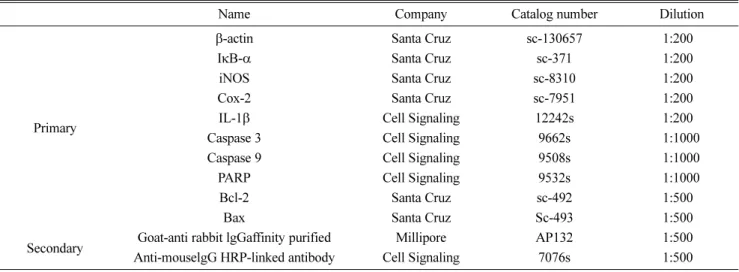
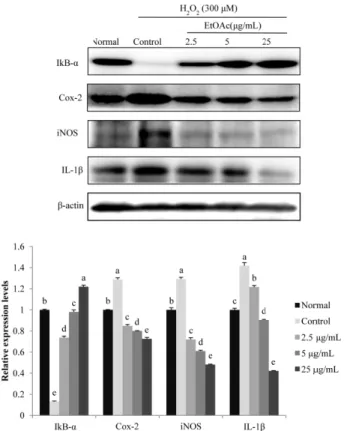
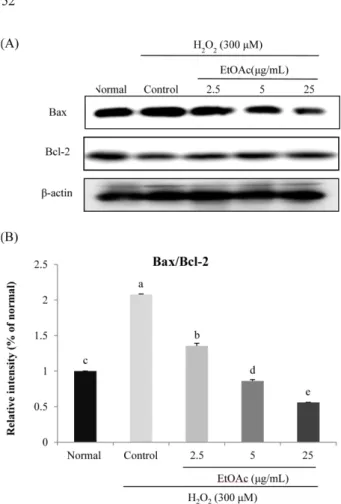
24(3) : 148-154 (2018)
https://doi.org/10.20307/nps.2018.24.3.148
148
Acer okamotoanum Inhibit the Hydrogen Peroxide-Induced Oxidative Stress in C6 Glial Cells
Soo Yeon Choi
1, Ji Hyun Kim
1, Norman G. Quilantang
2, Sanghyun Lee
2,*, and Eun Ju Cho
1,*
1
Department of Food Science and Nutrition & Kimchi Research Institute, Pusan National University, Busan 46241, Republic of Korea
2