발생 중인 포유류 망막으로 골수기질세포의 이식
이은실1, 권오주2, 예은아3, 전창진1,
*
1경북대학교 자연과학대학 생물학과, 대구 702-701
2부산정보대학 보건웰빙학부 안경광학과, 부산 616-737
3신경과학프로그램, 아이오와 주립대학교, 에임즈, 아이오와 50011, 미국 투고일(2013년 10월 31일), 수정일(2013년 11월 21일), 게재확정일(2013년 12월 14일)
···
목적: 골수기질세포는 생체 내·외에서 신경세포와 신경교세포로 교차분화 할 수 있는 능력을 가지고 있는 것으로 밝혀져 있다. 발생 중인 숙주 환경에 따라 이식된 골수기질세포의 생존여부, 형태학적 그리고 분자적 분화영향을 조 사하기 위해 브라질산 주머니쥐 안구에 마우스 골수기질세포를 이식하였다. 방법: GFP를 발현하는 골수기질세포를 발생 중인 브라질산 주머니쥐의 각 시기별로 이식하여, 이식 후 최대 4주까지 생존시킨 후 각 시기별로 면역조직화 학법을 시행하였다. 결과: 이식한 골수기질세포의 일부는 숙주동물의 유리체 내에서 생존하며 일부 돌기를 내는 신경 세포로 형태학적 분화가 됨을 관찰할 수 있었다. 또한 유리체에 존재하는 일부 세포는 신경세포 표지인자인 TuJ1(class III β-tubulin), 신경교세포 표지인자인 GFAP(glial fibrillary acidic protein), 또는 신경줄기세포 표지인자인 Nestin 단백 질을 발현하였다. 게다가, 일부 골수기질세포는 신경절세포층으로 이동함을 관찰했으나, 이동한 세포들은 형태학적 또 는 분자적 분화를 나타내지는 않았다. 결론: 이번 연구에서 가장 효율적인 이식시기는 생후 16일째의 포유류 망막으 로, 이는 망막세포의 분화양상과 층분화 패턴으로 미뤄볼 때 생후 4~5일 정도의 마우스 망막과 발생학적으로 상동함 을 알 수 있었다. 또한 이식 받은 숙주 망막의 미세환경이 이식된 세포운명에 영향을 미치는 것을 확인할 수 있었다.
주제어:골수기질세포, 발생, 망막, 이식, 분화, 이동
···
서 론
신경세포는 극히 제한적인 자기 회복능력만 가지고 있 기에 이의 사멸/손상으로 인한 중추신경계의 신경퇴행성 질환은 특별히 난치성 질환에 해당한다. 그리하여 신경세 포의 사멸을 방지하거나 손상에 의해 소멸된 신경세포들 을 회복시키기 위하여 다양한 노력들이 이뤄지고 있다. 그 중 한 방법인 줄기세포 이식치료는 줄기세포의 다양한 세 포로 분화 가능한 능력을 이용하여 손상된 신경 조직을 재생 할 수 있는 잠재력이 큰 치료법으로 대두되고 있으 며, 많은 연구에서 이를 효과적으로 이용하려는 노력이 진 행되고 있다.[1,2]
망막은 중추신경계의 한 부분으로, 다른 조직에 비해 줄 기세포 치료를 위한 표적으로서의 특별한 장점을 가지고 있다. 망막은 복잡한 외과적 수술 없이 유리체를 통한 접 근이 용이하며, 눈의 면역회피기능(immune privilege)은 이식된 세포의 거부 반응을 감소시킨다.[3] 이 때문에 망막 은 중추신경계의 발달(development), 가소성(plasticity), 그 리고 재생(regeneration) 연구와 줄기세포의 연구에 이상적
인 실험 모델로서의 역할을 수행할 수 있다. 이에 따라 현 재 다양한 줄기세포 이식을 통해 망막퇴행성 질환의 진행 을 지연시키거나, 손상된 신경 세포를 재생하고자 하는 연 구가 활발하게 진행되고 있다. 망막퇴행성 질환 모델의 유 리체방(vitreous chamber)이나 망막밑공간(subretinal space)에 망막전구세포(retinal progenitor cell)를 이식한 결과, 이식 된 전구세포들이 로돕신(rhodopsin), 옵신(opsin), 리커버린 (recoverin) 등 광수용체 단백질을 발현할 뿐만 아니라 빛 자극에 대한 반응도가 향상되는 것을 보여주고 있다.[4,5]
그 중 유도만능줄기세포는 일부 망막신경세포와 망막색소 상피세포로 분화함으로써 망막 질환의 치료의 가능성을 보여 주었고, 배아줄기세포를 이용하여 3D 망막 조직 형 성에 성공하여 줄기세포를 이용한 인공망막의 가능성을 보여주고 있다.[6-8] 하지만 이러한 전구세포나 줄기세포들 은 면역 거부와 윤리적인 문제가 존재하기에, 많은 연구자 들은 성체줄기세포들을 이용하는 연구들도 활발하게 진행 하고 있다. 성체줄기세포는 다른 줄기세포보다 상대적으 로 분리 및 세포배양이 쉽고 자가이식을 통해 면역억제가 필요 없으며 윤리적인 문제가 없기에 이식치료로서 매우
*Corresponding author: Chang-Jin Jeon, TEL: +82-53-950-5343, E-mail: [email protected]
<초청논문>
큰 기대를 모으고 있다. 그 중 골수기질세포(중간엽 줄기 세포)는 뼈, 연골, 지방, 근육, 간, 신장, 심장뿐만 아니라 신경세포로도 교차분화(trans-differentiation) 할 수 있다.
망막밑공간으로 이식된 골수기질세포는 광수용단백질을 발현하고 망막상피색소세포 등 망막의 특정 세포로도 분 화한다고 보고됨에 따라 골수기질세포의 다양한 망막세포 분화로의 잠재력을 보여주고 있다.[9-12]
이와 같이 다양한 종류의 공여세포를 숙주 망막에 이식 한 이후 세포의 분화와 통합을 확인하는 연구와 더불어 이식 받는 숙주 망막의 미세환경이 이식된 세포의 분화에 영향을 준다는 것 또한 밝혀져 있다.[13-15] 망막은 발생 과 정 동안 다양한 신호전달인자와 망막분화를 결정짓는 인 자들이 활성화되는 미세환경을 가지며, 이러한 다양한 인 자들이 망막에 이식된 줄기세포의 효율적인 분화와 통합 을 유도한다고 보여주고 있다.[16-18] 하지만 이러한 미세환 경 변화에 따른 이식된 줄기세포의 생존, 분화에 대한 체 계적인 연구는 아직 미흡하다.
브라질산 주머니쥐(Brazilian gray short-tailed opossum, Monodelphis domestica)는 남아메리카 산의 유대목 (Marsupialia) 포유동물로서, 다른 포유동물과는 달리 새끼 가 태어날 때 극도로 미성숙한 상태이며 출생 후에도 오 랜 발생 기간을 갖게 된다. 따라서 태아와 같은 발생단계 를 가진 모델로 큰 외과적인 수술 없이 생체 외에서의 실 험을 통해 이식된 줄기세포의 운명을 알아내는 데 최적일 수 있다. 본 연구자는 이러한 최적의 동물모델을 이용하여 발생 중 망막에 골수기질세포를 이식 한 후 이식된 세포 가 망막내로의 생존, 구조적, 분자적 분화에 미치는 영향 을 연구하였다. 이러한 연구는 줄기세포치료기술을 개발 하기 위한 기반지식을 제공하는데 있어 반드시 필요한 가 치가 있다고 사료된다. 또한 이에 따른 연구결과는 기존의 이식을 통한 치료회복 연구를 더욱 발전시키고, 손상된 시 각계의 재생과 회복을 촉진 및 치료하는 많은 후속적인 방법을 개발하는데 크게 기여 할 것이라 사료된다.
대상 및 방법
1. 대상
이번 실험에 사용된 브라질산 주머니쥐는 남 아메리카 산의 작은 유대목 동물로 현재 인간의 발생, 가소성, 재생, 생리학과 질병 민감성 등과 관련된 광범위한 주제에서 비 교 연구를 위한 귀중한 동물 모델로 사용되고 있다.[19] 브 라질산 주머니쥐의 임신기간은 13.5일이며, 한번에 3~13 마리의 새끼를 낳는데, 새끼가 태어난 날을 1PN(1 postnatal day)으로 정하고, 개안(eye opening)은 35PN 정도에 일어 난다. 이번 실험에서는 4PN에서부터 35PN 동물을 사용하
였으며, 이식 후 생존기간을 7, 14, 그리고 28DPT(28 day post transplantation)로 두었다. 각 PN당 6-12마리를 이식 하였으며 각 DPT당 3마리를 배정하였다. 모든 동물실험과 정들은 미국 시각 및 안과학회(ARVO)의 규정을 준수하면 서 수행되었다.
2. 방법
이식 후 추적을 위해 GFP로 표지된 마우스 골수기질세 포는 Texas A&M Health Science Center에서 구입하였으 며, Iscove’s Modified Dulbecco's Medium(IMDM; Gibco- BRL, Rockville, MD, USA), 2 mM L-glutamine, 100 U/ml penicillin, 100 µg/ml streptomycin, 0.25 µg/ml amphotericin B, 10% FBS, 10% horse serum(모두 Invitrogen Corp., Carsbad, CA, USA)에 배양하였다. 배양된 골수기질세포들은 spheres 상태로 collect 된 후에 800 g에서 3분간 원심분리한 후에 골수기질세포를 Dulbecco’s PBS (Gibco-BRL)에 부유시켰 다. 동물들은 흡기 챔버에서 3% halothane이 30% NO와 70% O2로 혼합되어 있는 마취제를 이용해서 마취하였다.
주머니쥐의 새끼들은 20PN까지는 어미의 젖에 입이 완전 히 물려있는 상태로 있으며 어미로부터 분리되지 않기 때 문에 어미와 새끼를 함께 마취하고, 20PN 이후의 동물들 은 개별적으로 마취하였다. 이식은 비스듬하게 잘린 glass micropipette를 이용하여 진행 하였는데 micropipette은 0.9% saline-filled polyethylene tubes에 연결시킨 후, 다시 20-L syringe(Hamilton, Reno, NV)에 연결을 시켜 정확한 양을 조절할 수 있도록 했다. 각각의 동물들은 눈의 등측 면(dorsolateral aspect of the eye)쪽으로부터 유리체 안으 로 (intraocular injections) 이식을 받았다. 20PN 이전의 동 물은 1.0~1.5 µl의 세포 부유액(50,000 cells/µl)을 이식 받 았으며 20PN 이후의 동물들은 2.0 µl의 세포 부유액을 이 식 받았다.
특정 생존 기간 후, 동물들을 halothane으로 호흡 마취하 고 안구 및 망막을 적출해내는데, 그 후 고정방법은 동물 의 이식시점의 나이와 이식 후 생존 나이의 합에 따라 다 른 방법으로 진행하였다. 그 합이 20일 이하인 동물들은 마취 후 두부 전체를 4% paraformaldehyde에서 이틀 동안 고정하고, 합이 20~40일 기간의 동물들은 마취 후 두부 전 체를 4% paraformaldehyde에서 고정한 상태에서 안구 부 분만을 따로 분리 해, 같은 고정액에서 하루 동안 더 고정 하였다. 마지막으로 합이 40일 이상의 동물들은 위의 고 정액으로 30분간 심장 관류법으로 조직을 고정한 후 안구 를 분리하여 같은 고정액에서 하루 동안 더 고정하였다.
고정된 조직들은 고정 후 0.1M phosphate buffer(PB)로 3번의 세척단계를 거친 후 조직들은 설탕용액(10, 20, and 30% sucrose/0.1M PB)에 담궈 cryoprotection하였다. 이 후
조직들은 OCT(optimal cutting temperature) 표본제작 용액 (Tissue-Tek; VWR International, West Chester, PA)을 이용 하여 표본을 만든 후, −80oC에 하루 이상 보관 후 냉동미 세절단기(cryostat)로 20 µm 두께로 수직방향으로 잘라
−20oC에 보관하였다.
준비된 조직은 다음의 표준면역조직화학적 방법의 단계 를 거쳤다. 1차 항체는 Rabbit anti-Nestin antibody(1:200- 400, Chemicon, Temecula, CA), Mouse anti-TuJ1 antibody (class III β-tubulin, 1:500, R&D Systems, Minneapolis, MN, USA) 및 Mouse anti-GFAP antibody(glial fibrillary acidic protein, 1:500, ICN, Aurora, OH, USA)으로 4oC 항 온기에서 24~48시간 동안 반응을 시킨 후 3번의 세척 단 계를 거쳤다. 2차 항체는 Cy3-conjugated secondary antibody(Jackson Immuno Research Laboratories, PA, USA;
1:400)을 이용하였다. 여기서 GFP로 이식된 세포는 GFP 의 형광감도를 증가시키기 위해서 Chicken anti-GFP(Abcam, Cambridge, UK)를 처리하여 Biotinylated donkey anti-
chicken IgY(Millipore, Bedford, MA, USA)와 anti-Biotin Streptoavidin-Cy3(Jackson Immuno Research Laboratories;
1:400)의 연속 결합을 이용하였다. 면역조직화학 반응이 끝난 조직은 핵의 염색을 위해 4'-6-Diamidino-2-Phenylindole (DAPI, 1:1000, Sigma-Aldrich Corp., St. Louis, MO, USA)으 로 30분간 염색 후 슬라이드 글라스에 올리고 Vectashield- mounting medium(Vector Laboratories)을 사용하여 커버글 라스를 덮어 마무리를 하였다. 모든 단계에서 0.1M PB용 액을 사용하였다. 이식된 망막은 이식되지 않은 같은 나이 의 정상망막과 비교하였으며, 이식 후 생존기간 안에 정상 적인 발달이 일어났는지를 비교하였다. 조직의 관찰은 형 광 장치가 부착된 Zeiss Axioplan(Ver. 4, Carl Zeiss) 현미 경을 이용하였다.
결 과
Fig. 1은 20 µm두께로 수직절편한 망막조직에서 이식한
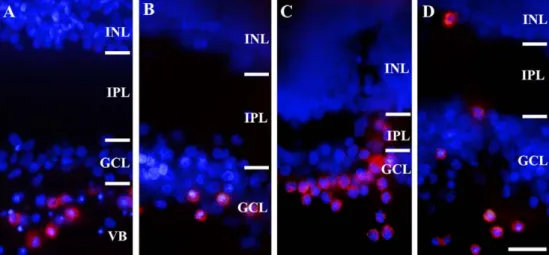
Fig. 1. Migration of transplanted bone marrow stromal cells. (A) Transplanted bone marrow stromal cells (red) migrated throughout the 16PN 28DPT host vitreous body. (B) Transplanted bone marrow stromal cells (red) migrated into the ganglion cell layer at 16PN 14DPT retina. (C) Transplanted bone marrow stromal cells (red) passed through the ganglion cell layer and migrated into the inner plexiform layer at 16PN 14DPT retina. (D) Transplanted bone marrow stromal cells (red) migrated closely to inner nuclear layer at 16PN 14DPT retina. VB, vitreous body; GCL, ganglion cell layer; IPL, inner plexiform layer;
INL, inner nuclear layer. Scale bar = 50 µm.
Fig. 2. Morphological differentiation of transplanted bone marrow stromal cells. (A-B) The transplanted cells morphologically differentiated into the cell with long processes in the vitreous body. Arrowheads indicate processes. Scale bar = 50 µm.
세포들을 추적한 것이다. GFP로 표지한 이식된 세포는 Biotin/Streptoavidin-Cy3를 통해 확인했으며, 망막세포의 핵을 염색하기 위해 DNA 결합 형광 물질인 DAPI를 이용 하였다. Fig. 1A는 16PN 28DPT 망막으로 이식된 골수기 질세포들이 망막의 유리체 내에 분포하며 신경절세포층 부근까지 이동한 것을 보여준다. Figs. 1B-D는 16PN 14DPT의 망막으로 이식된 골수기질세포들이 망막의 신경 절세포층에 존재하는 것을 보여주고 있으며(Fig. 1B), 이 중 이식한 세포 중 일부는 내망상층을 통과하여(Fig. 1C), 내핵층 가까이까지 이동한 것을 보여주고 있다(Fig. 1D).
이를 통해 이식된 골수기질세포 일부가 망막 내로 이동하 는 것을 확인할 수 있다.
Fig. 2은 이식된 세포들이 신경돌기가 나오는 것을 관찰 함에 따라 형태적으로 분화됨을 확인할 수 있다. Fig. 2A 는 4PN 21DPT이며 Fig. 2B는 4PN 28DPT로 골수기질세 포들이 안구의 유리체 내에서 이식 후 3~4주가 지나도 생 존함과 형태학적으로 신경돌기를 내는 것을 확인할 수 있 다. 하지만 이러한 이식된 골수기질세포의 형태학적 분화 는 유리체에서만 일어나며 망막세포층으로 이동한 세포들 에서는 나타나지 않았다.
이식된 골수기질세포가 망막세포로 분자적으로 분화하
는 지 확인하기 위해 면역조직화학법을 수행하였다. Fig.
3은 이식한 세포가 망막세포층으로 이동한 16PN 14DPT 의 조직으로 초기신경세포의 표지인자인 TuJ1과 신경교세 포의 표지인자인 GFAP로 염색한 것이다. Fig. 3A는 이식 한 골수기질세포가 신경절세포층에 존재하는 것을 나타내 며 Fig. 3B는 이를 TuJ1으로 염색한 것이다. Fig. 3C는 3A 와 3B를 merge한 것으로 이식된 골수기질세포가 초기신 경세포 발현물질인 TuJ1을 발현하지 않음을 보여준다. 또 한 Fig. 3D와 3G는 이식된 골수기질세포가 신경절세포층 에 존재하는 것을 나타내며 Fig. 3E와 3H는 이를 GFAP으 로 염색한 것이다. Fig. 3F는 3D와 3E를 merge한 것이며 Fig. 3I는 3G와 3H를 merge 한 것으로 이식된 골수기질세 포가 신경교세포 발현물질인 GFAP를 발현하지 않음을 확 인할 수 있다. 이를 통해 이식한지 14일이 지나도 이식된 세포들은 신경세포나 신경교세포로 분화되지 않음을 보아 망막 내 신경세포층으로 이동은 되었으나 망막세포로의 분화는 일어나지 않음을 확인할 수 있다.
Fig. 4는 4PN 28DPT 의 안구조직으로 유리체에 존재하 는 이식된 세포가 분자적으로 분화하는지를 확인한 결과 이다. Fig. 4A는 이식된 골수기질세포가 유리체에 존재하
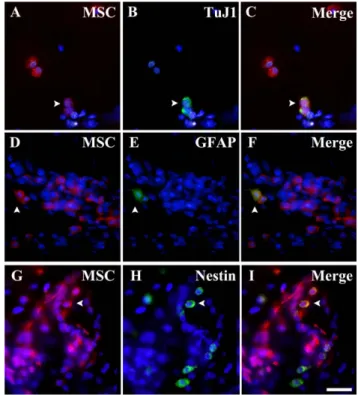
Fig. 3. Molecular differentiation of transplanted bone marrow stromal cells at 16PN 14DPT host retina. Transplanted cells (GFP-Biotin Streptoavidin-Cy3; A, D, and G), primary antibodies/ FITC immunoreactivity (TuJ1 (B), and GFAP (E, H)) and merged images (C, F, and I).
Migrated transplanted cells in the ganglion cell layer are not immunoreactive for antibodies against TuJ1 or GFAP. GCL, ganglion cell layer. Scale bar = 50 µm.
Fig. 4. Molecular differentiation of transplanted bone marrow stromal cells at 4PN 28DPT host vitreous body.
Transplanted cells (GFP-biotin streptoavidin-Cy3; A, D, G), primary antibodies/FITC immunoreactivity (TuJ1 (B), GFAP (E), and Nestin (H)) and merged images (C, F, I). Arrowheads indicate transplanted cells in the host vitreous body that co-express TuJ1, GFAP or Nestin.
Scale bar = 50 µm.
는 것을 나타내며, Fig. 4B는 이를 TuJ1으로 염색한 것이다.
Fig. 4C는 4A와 4B를 merge한 것으로 이식된 일부 골수 기질세포가 초기신경세포 발현물질인 TuJ1을 발현하는 것 을 확인할 수 있다. 또한 Fig. 4D는 이식된 골수기질세포 가 유리체에 존재하는 것을 나타내며 Fig. 4E는 이를 GFAP로 염색한 것이다. Fig. 4F는 4D와 4E를 merge한 것 으로 이식된 일부 골수기질세포가 신경교세포 발현물질인 GFAP를 발현하는 것을 확인할 수 있다. 마지막으로 Fig.
4G는 이식된 골수기질세포가 유리체에 존재하는 것을 나 타내며 4H는 신경전구세포의 표지인자인 Nestin으로 염색 한 것이다. Fig. 4I는 4G와 4H를 merge한 것으로 이식된 골수기질세포가 신경전구세포로 분화함을 확인할 수 있다.
본 연구를 통해 이식된 세포가 망막 내에 존재하는 신 경세포층으로 이동은 되었으나, 이식된 세포가 망막에 존 재하는 특정세포군으로 형태적, 분자적으로 분화됨은 확 인 할 수 없었다. 하지만 망막 내로 이동하지 못하고 유리 체에 존재하는 일부 골수기질세포는 형태적, 분자적으로 분화하는 것을 보여주고 있다. 이를 통해 이식된 골수기질 세포가 유리체에서 매우 제한적으로 분화됨을 확인할 수 있다.
고 찰
본 연구에서는 망막발생 중에 골수기질세포를 이식한 결과 이식된 세포는 유리체에 산재되었으며 그 중 일부 이식된 세포들은 망막 내경계막(inner limiting membrane) 을 통과하여 망막신경세포층 내로 이동하는 것을 확인할 수 있었다. 망막 내로 이동한 일부의 세포들은 형태학적 분화와, 신경세포 또는 신경교세포로의 분자적인 분화가 진행되지 않았지만 유리체에 남아있는 세포들은 이러한 분화가 일어나는 것으로 확인되었다. 망막 내로 이동한 골 수기질세포의 분화가 이뤄지지 않는 직접적인 원인은 밝 힐 수 없으나, 망막세포 주위기질세포(extracellular matrix) 로부터 결합이 잘 이뤄지지 않았거나, 기질세포에서 분비 하는 신호인자를 잘 받아들이지 못함으로 사료된다. 하지 만 이와 같은 결과와는 달리 발생 중인 망막 안으로 이식 된 마우스 뇌 전구세포(murine brain progenitor cells), 마 우스 망막 전구세포(murine retinal progenitor cells), 그리 고 성체 쥐 해마 전구세포(adult rat hippocampal progenitor cells)는 망막에 존재하는 여러 세포층으로 이동 한 후 특 정 신경 세포군으로 형태학적으로 분화가 일어난다는 결 과가 보고되어 있다.[14,15,20,21,22] 이를 통해 브라질산 주머니 쥐의 발생 중인 망막으로 이식한 골수기질세포가 망막으 로 통합되는 능력은 다른 신경 유래 전구세포보다 극히 제한적인 것을 확인할 수 있다.
본 연구에서 이식된 골수기질세포가 망막 신경세포층으 로 이동되는 결과들은 주로 태어난 지 16일째에 이식한 브라질산 주머니쥐에서 나타난 것이며, 이보다 초기 단계 에 이식한 결과들에서는 대부분 이식한 세포들이 유리체 에 존재하는 것을 확인할 수 있었다. 기존 연구에서 브라 질산 주머니쥐의 새끼는 다른 포유류와는 달리 임신 14일 후 태어나며, 이때에는 극도로 미성숙한 상태이다.[14,15,23,24]
즉, 출생 시의(생후 1일) 브라질산 주머니쥐의 망막은 진 성태반동물의 포유류와 상대적으로 비교해보면 배아 상태 로서, 출생 후에도 신경 발생이 진행됨을 보인다.[24]
Thymidine 유사체인 5'-bromodeoxyuridine(BrdU)를 이용 한 이 동물의 망막세포 발생학적 연구결과에 따르면 망막 신경절세포는 임신 후 13일이 되면 형성되기 시작해서 생 후 12일 쯤이 되면 세포형성이 최고조에 이르고 생후 20 일쯤에 완성이 된다. 내핵층의 세포 생성은 생후 1일 이후 에 시작되고 생후 12일 정도면 세포형성이 최고조에 이르 며 생후 30일쯤에 완성이 된다. 광수용세포는 두 시기에 걸쳐서 형성되는데, 대략 생후 2-8일에 원추세포의 생성이 일어나고, 그 후 생후 10-20일에 간상세포의 형성이 일어 난다. 세포분화나 망막 층 패턴으로 비교해보면, 이번 연 구에 이용된 생후 16일 되는 브라질산 주머니쥐는, 출생 후 4-5일쯤 되는 마우스 망막과 비슷한 상태임을 확인할 수 있다.[14,25]
흥미롭게도, 발생 중인 마우스 망막에 인간 유래 골수에 서 추출한 중간엽 줄기세포와 지방유래기질세포를 이식한 이전 연구 결과에서도 다른 발생학적 단계보다 출생 후 5일 에 이식한 줄기세포가 가장 효과적임이 보고되고 있다.[26,27]
이번 연구에서도 발생학적으로 상동한 단계에서 가장 효 과적인 결과를 나타내었다. 이와 같은 결과들은 마우스를 기준으로 출생 후 5일정도의 망막 발생 단계가 이식된 중 간엽 줄기세포와 기질세포가 망막 내로의 분화와 이동을 보여주는 가장 효율적인 숙주 단계임을 제시하고 있다.
망막 발생동안 다양하게 발현되는 뉴로트로핀분자, 신 경분화인자와 다양한 신호 전달 과정들은 특정 망막세포 의 생존과 분화에 관여하는 것으로 밝혀져 있다. 발생 중 인 닭의 망막에서 BDNF과 NT-3는 망막세포의 분화를 촉 진하며, CNTF는 출생 후의 마우스 망막에서 광수용세포 의 분화와 뮬러세포의 생성을 조절하는 것으로 밝혀져 있
다.[28-31] 이러한 망막 발생 중의 여러 인자들은 이식한 줄
기세포가 숙주 망막으로 통합 및 분화하는 데 있어 그 가 능성을 더욱 촉진시킨다고 보고되고 있다.[13,14,15,32] 이러한 여러 인자들은 나이 의존적으로 발현되며, 특히 5PN 마우 스 망막은 다른 발생단계와 비교하여 무축삭세포, 이극세 포, 뮬러세포의 분화가 제일 활발하게 일어난다.[33,34]또한 Mash1, Hes1, Hesr2, Notch1, Math3와 같은 내핵층의 신
경세포 분화에 관련된 여러 유전자들의 발현정도는 다른 연령의 눈과 비교하여 5PN에서 가장 높다.[34-36] 이번 연구 결과에서 마우스 5PN과 상동한 발생단계에서 망막 내로 의 이동이 다른 단계보다 많은 이유는 명확하지 않지만, 이 발생단계에서 관련된 여러 분화 인자들이 이식한 골수 기질세포의 생존, 망막 내로의 이동, 망막세포로의 분화와 통합에 관련을 주는 것으로 사료된다. 그리하여 앞으로 줄 기세포 이식에 있어 중요한 분화 인자들을 찾는 분자적인 연구를 진행함으로써 주요 인자와 동시에 줄기세포를 이 식하는 새로운 이식 치료 방법을 개발할 예정이다.
결 론
본 연구에서는 망막 발생 단계에 유리체 내로의 골수기 질세포 이식을 통해 이식된 세포가 생존함과 신경절세포 층으로 이동함을 확인할 수 있었다. 또한 이식한 세포는 유리체에서는 형태학적 분화 그리고 신경세포나 신경교세 포로의 분자적인 분화를 확인했으나, 망막 내로 이동된 세 포는 이러한 분화가 확인되지 않았다. 이와 같은 연구는 앞으로의 이식연구에 있어 공여세포의 종류 뿐만 아니라 숙주 망막의 미세환경 또한 이식 후 결과에 크게 영향을 주는 것을 보여주는 데 의의가 있다.
감사의 글
이 논문은 2011년도 정부(교육부)의 재원으로 한국연구 재단의 지원을 받아 수행된 기초연구사업임(No. 2011- 0024882).
REFERENCES
[1] Ramsden CM, Powner MB, Carr AJ, Smart MJ, da Cruz L, Coffey PJ. Stem cells in retinal regeneration: past, present and future. Development. 2013;140(12):2576-2585.
[2] Wang NK, Tosi J, Kasanuki JM, Chou CL, Kong J, Par- malee N et al. Transplantation of reprogrammed embry- onic stem cells improves visual function in a mouse model for retinitis pigmentosa. Transplantation. 2010;89(8):911-919 [3] Streilein JW. Ocular immune privilege: therapeutic oppor-
tunities from an experiment of nature. Nat Rev Immunol.
2003;3(11):879-889.
[4] Klassen HJ, Ng TF, Kurimoto Y, Kirov I, Shatos M, Cof- fey P et al. Multipotent retinal progenitors express devel- opmental markers, differentiate into retinal neurons, and preserve light-mediated behavior. Invest Ophthalmol Vis Sci. 2004;45(11):4167-4173.
[5] Stern JH, Temple S. Stem cells for retinal replacement
therapy. Neurotherapeutics. 2011;8(4):736-743.
[6] Meyer JS, Shearer RL, Capowski EE, Wright LS, Wallace KA, McMillan EL et al. Modeling early retinal develop- ment with human embryonic and induced pluripotent stem cells. Proc Natl Acad Sci U S A. 2009;106(39):16698- 16703.
[7] Buchholz DE, Hikita ST, Rowland TJ, Friedrich AM, Hinman CR, Johnson LV et al. Derivation of functional retinal pigmented epithelium from induced pluripotent stem cells. Stem Cells. 2009;27(10):2427-2434.
[8] Nistor G, Seiler MJ, Yan F, Ferguson D, Keirstead HS.
Three-dimensional early retinal progenitor 3D tissue con- structs derived from human embryonic stem cells. J Neu- rosci Methods. 2010;190(1):63-70.
[9] Castanheira P, Torquetti L, Nehemy MB, Goes AM. Reti- nal incorporation and differentiation of mesenchymal stem cells intravitreally injected in the injured retina of rats. Arq Bras Oftalmol. 2008;71(5):644-650.
[10] Gong L, Wu Q, Song B, Lu B, Zhang Y. Differentiation of rat mesenchymal stem cells transplanted into the sub- retinal space of sodium iodate-injected rats. Clin Experi- ment Ophthalmol. 2008;36(7):666-671.
[11] Arnhold S, Heiduschka P, Klein H, Absenger Y, Basnao- glu S, Kreppel F et al. Adenovirally transduced bone mar- row stromal cells differentiate into pigment epithelial cells and induce rescue effects in RCS rats. Invest Ophthalmol Vis Sci. 2006;47(9):4121-4129.
[12] Tomita M, Adachi Y, Yamada H, Takahashi K, Kiuchi K, Oyaizu H et al. Bone marrow-derived stem cells can dif- ferentiate into retinal cells in injured rat retina. Stem Cells. 2002;20(4):279-283.
[13] MacLaren RE, Pearson RA, MacNeil A, Douglas RH, Salt TE, Akimoto M et al. Retinal repair by transplanta- tion of photoreceptor precursors. Nature. 2006;444(7116):
203-207.
[14] Sakaguchi DS, Van Hoffelen SJ, Theusch E, Parker E, Orasky J, Harper MM et al. Transplantation of neural pro- genitor cells into the developing retina of the Brazilian opossum: an in vivo system for studying stem/progenitor cell plasticity. Dev Neurosci. 2004;26(5-6):336-345.
[15] Van Hoffelen SJ, Young MJ, Shatos MA, Sakaguchi DS.
Incorporation of murine brain progenitor cells into the developing mammalian retina. Invest Ophthalmol Vis Sci.
2003;44(1):426-434.
[16] Amato MA, Boy S, Perron M. Hedgehog signaling in ver- tebrate eye development: a growing puzzle. Cell Mol Life Sci. 2004;61(7-8):899-910.
[17] Esteve P, Bovolenta P. Secreted inducers in vertebrate eye development: more functions for old morphogens. Curr Opin Neurobiol. 2006;16(1):13-19.
[18] Van Raay TJ, Vetter ML. Wnt/frizzled signaling during verte- brate retinal development. Dev Neurosci. 2004;26(5-6):352- 358.
[19] VandeBerg JL, Robinson ES. The Laboratory Opossum
(Monodelphis Domestica) in Laboratory Research. ILAR J. 1997;38(1):4-12.
[20] Palmer TD, Takahashi J, Gage FH. The adult rat hippoc- ampus contains primordial neural stem cells. Mol Cell Neurosci. 1997;8(6):389-404.
[21] Sakaguchi DS, Van Hoffelen SJ, Young MJ. Differentia- tion and morphological integration of neural progenitor cells transplanted into the developing mammalian eye.
Ann N Y Acad Sci. 2003;995:127-139.
[22] Shatos MA, Mizumoto K, Mizumoto H, Kurimoto Y, Klassen H, Young MJ. Multipotent stem cells from the brain and retina of green mice. J Regen. Med. 2001;2(3):
13-15.
[23] Sakaguchi DS, Iqbal J, Sonea I, Jacobson CD. The gray short-tailed opossum: a novel model for mammalian development. Lab animal. 1995;24(6):24-29.
[24] Greenlee MH, Swanson JJ, Simon JJ, Elmquist JK, Jacobson CD, Sakaguchi DS. Postnatal development and the differential expression of presynaptic terminal-associ- ated proteins in the developing retina of the Brazilian opossum, Monodelphis domestica. Brain Res Dev Brain Res. 1996;96(1-2):159-172.
[25] Greenlee MH, Roosevelt CB, Sakaguchi DS. Differential localization of SNARE complex proteins SNAP-25, syn- taxin, and VAMP during development of the mammalian retina. J Comp Neurol. 2001;430(3):306-320.
[26] Lee ES, Yu SH, Jang YJ, Hwang DY, Jeon CJ. Transplan- tation of bone marrow-derived mesenchymal stem cells into the developing mouse eye. Acta Histochem Cytochem.
2011;44(5):213-221.
[27] Yu SH, Jang YJ, Lee ES, Hwang DY, Jeon CJ. Transplan- tation of adipose derived stromal cells into the developing mouse eye. Acta Histochem Cytochem. 2010;43(6):123-
130.
[28] Fuhrmann S, Kirsch M, Hofmann HD. Ciliary neurotrophic factor promotes chick photoreceptor development in vitro.
Development. 1995;121(8):2695-2706.
[29] Goureau O, Rhee KD, Yang XJ. Ciliary neurotrophic fac- tor promotes muller glia differentiation from the postnatal retinal progenitor pool. Dev Neurosci. 2004;26(5-6):359- 370.
[30] Kirsch M, Fuhrmann S, Wiese A, Hofmann HD. CNTF exerts opposite effects on in vitro development of rat and chick photoreceptors. Neuroreport. 1996;7(3):697-700.
[31] Xie HQ, Adler R. Green cone opsin and rhodopsin regu- lation by CNTF and staurosporine in cultured chick pho- toreceptors. Invest Ophthalmol Vis Sci. 2000;41(13):4317- 4323.
[32] Takahashi M, Palmer TD, Takahashi J, Gage FH. Wide- spread integration and survival of adult-derived neural progenitor cells in the developing optic retina. Mol Cell Neurosci. 1998;12(6):340-348.
[33] Cepko CL, Austin CP, Yang X, Alexiades M, Ezzeddine D. Cell fate determination in the vertebrate retina. Proc Natl Acad Sci U S A. 1996;93(2):589-595.
[34] Dorrell MI, Aguilar E, Weber C, Friedlander M. Global gene expression analysis of the developing postnatal mouse retina. Invest Ophthalmol Vis Sci. 2004;45(3): 1009-1019.
[35] Furukawa T, Mukherjee S, Bao ZZ, Morrow EM, Cepko CL. rax, Hes1, and notch1 promote the formation of Mller glia by postnatal retinal progenitor cells. Neuron.
2000;26(2):383-394.
[36] Hatakeyama J, Kageyama R. Retinal cell fate determina- tion and bHLH factors. Semin Cell Dev Biol. 2004;15(1):83- 89.
Transplantation of Marrow Stromal Cells into the Developing Mammal Retina
Eun-Shil Lee1, Oh-Ju Kwon2, Eun-Ah Ye3, and Chang-Jin Jeon1,
*
1Dept. of Biology, College of Natural Sciences, Kyungpook National University, Daegu 702-723, Korea
2Dept. of Ophthalmic Optic, Busan College of Information Technology, Busan 616-737, Korea
3Neuroscience Program, Iowa State University, Ames, Iowa 50011, USA
(Received October 31, 2013: Revised November 21, 2013: Accepted December 14, 2013)
Purpose: Marrow stromal cells (MSCs) have been known for their potential to trans-differentiate into neural and glial cells in vitro and in vivo. To investigate the influence of the developing host environment on the survival and morphological and molecular differentiation, murine MSCs transplanted into the eye of Brazilian opossum (Monodelphis domestica). Methods: Enhanced green fluorescent protein (GFP) - expressing MSCs were transplanted into developing Brazilian opossums. Animals were allowed to survive for up to 4 weeks after transplantation, at which time the eyes were prepared for immunohistochemical analysis. Results: Some transplanted MSCs survived and showed morphological differentiation into neural cells with some processes within the host vitreous chamber. Some transplanted cells expressed class III β-tubulin (TuJ1, a marker for neuronal cells) or glial fibrillary acid protein (GFAP, a marker for glial cells) or Nestin (a marker for neural stem cells). In addition, some transplanted cells were located in ganglion cell layer but did not show morphological and molecular differentiation. Conclusions: Our result show that the most effective stage of development for transplantation into the retina was postnatal day 16, which retinas developmentally corresponded to postnatal day 4-5 days mouse retina based on cell differentiation and lamination patterns. The present findings suggest that the age of the host appears to play a key role in determining cell fate in vivo.
Key words: Marrow stromal cells, Development, Retina, Transplantation, Differentiation, Migration