355 KOR. J. HORT. SCI. & TECH. 16(3), OCTOBER 1998
원예과학기술지 16(3):355-357. 1998溫度와 生長調節物質이 무궁화( Hibiscus syriacus
‘’ Honghwarang’) 삽수의 萌芽에 미치는 영향
백이화1*․조근호1․박천호1․허무룡2․곽병화1
1고려대학교 원예과학과, 2경상대학교 원예학과
Effects of Temperature and Plant Growth Regulators on Bud Sprouting of Stem Cutting of Hibiscus syriacus ‘Honghwarang’
Baek, Yi Hwa1*․Cho, Keun Ho1․Pak, Chun Ho1․Huh, Moo Ryong2․Kwack, Beyoung Hwa1
1
Dept. of Hort. Sci., Korea University, Seoul 136-701, Korea
2
Dept. of Horticulture, Gyeongsang National University, Chinju 660-701, Korea
*corresponding author
ABSTRACT Various temperatures and plant growth regulators were assessed
to investigate the nature of the dormancy and bud sprouting of stem cutting ofHibiscus syriacus ‘Honghwarang'. The bud sprouting was promoted as tem-
perature increased. Under 30℃ condition, stem cuttings began sprouting at 4 days after cutting and showed 100% sprouting at 6 days after cutting. With GA or BA treatment, the cuttings started sprouting at 6 days after cuttings under 25℃ condition and reached 65% sprouting, which was not different from the results without hormone treatments. However, ABA dipping under below 25℃ condition resulted in low sprouting, whereas under over 25℃ condition they started sprouting at 6 days after cutting and reached 93% at 12 days after cutting, which indicated that ABA is not effective on inhibition of sprouting under high temperature condition.Additional key words: abscisic acid, benzyladenin, dormancy, gibberellic acid
0 2 4 6 8 10 12
10 15 20 25 30
Temperature(℃)
∑12
Con ABA GA BA z)
3
Fig. 1. Effect of various temperatures and
plant growth regulators on bud sprout- ing of Hibiscus syriacus cv. Honghwa- rang during the 12-day experimental period after stem cutting. Each point represents the mean ∑12 of sixteen buds.서 언
나라꽃 무궁화에 대한 연구는 다방면으로 이 루어 졌지만 류 등(1974, 1975)의 보고 이후로 休眠生理에 대한 연구는 부족한 실정이다. 류 등(1975)의 무궁화 개화특성에 관한 보고에서 무궁화의 생육은 온도보다 일장에 지배되며 고 온장일에서는 생육이 계속되나 고온단일에서는 생육이 완전히 정지하였다고 했다. 따라서 무궁 화의 휴면이 단일 조건에서 온대원산 식물의 휴 면타파에 필요한 저온처리를 받아야 타파되는 지와 휴면타파를 촉진 또는 지연시키는데 관여 하는 외부 생장조절물질의 영향을 받는지 알아 보고자 하였다. 또한 Lang 등(1987)이 구분한 휴면단계 중 우리나라 기후조건에서 무궁화 휴 면의 깊이는 어느 정도인지 알아보고 그에 따른 생리적 변화에 대한 연구의 기초자료로 삼고자 본 실험을 시행하였다.
재료 및 방법
공시재료는 홍단심계 무궁화인 ‘홍화랑’으로 1997년 10월 20일에 고려대학교 화훼온실에서 생육하고 있는 당년생 가지를 눈(芽)과 잎 하나 만을 남기고 7cm정도로 조제하였다. 배양토는 버미큘라이트와 펄라이트를 1:1 (V/V)로 섞어 사용하였으며, 직경 10 cm 의 플라스틱화분에 한 화분 당 10주를 1반복으로 하여 처리 당 3 반복으로 완전임의배치하여 각 온도별 생장상 에 두었다. 萌芽는 눈이 두터워지면서 녹색을
나타낼 때를 기준으로 하였다.
온도 조건은 10, 15, 20, 25, 30℃로 하였고, 일장은 단일(8시간)조건으로 하였다. 생장조절 물질로는 ABA 200mg/L (천연 racemic형, Research Organics社), gibberellic acid 100mg/L (GA3, BDH Limited Poole Eng- land), benzyladenin 50mg/L(Research Or- ganics社)을 삽수에 24시간 침지처리 하였다.
조사항목은 腋芽가 처음 맹아하기까지 소요 되는 날과 그에 따른 出芽長과 出芽率을 조사하 였는데, 측정방법은 Timson(1965)에 의한 방 법으로, 삽목 후 1일만에 맹아가 되었다면 ∑12
value = 11, 2일 ∑12 value = 10, 12일만에 맹아가 되었다면 ∑12 value = 0으로 계산하였 다. 맹아에 따른 생리적 변화를 알아보기 위하 여 25℃의 생장상에서 삽목한 삽수 액아의 peroxidase (POD), catalase activity (CAT) 의 변화량은 Kar와 Mishra(1976)의 방법으로 측정하였다.
결과 및 고찰
무궁화의 휴면특성을 알아보기 위해 온도와 생장조절물질의 처리에 따른 맹아차이를 조사 한 결과는 Fig. 1과 같다. 맹아에 소요되는 시 일은 여러 온도 처리중 30℃에서 가장 빠른 맹 아율을 보였다. 생장조절제 처리의 경우 GA, BA처리는 무처리와 차이가 없었으나 ABA처리 에 의해서 맹아가 지연되었다(Fig. 1).
무처리의 맹아율은 보면 10℃에서는 거의 반
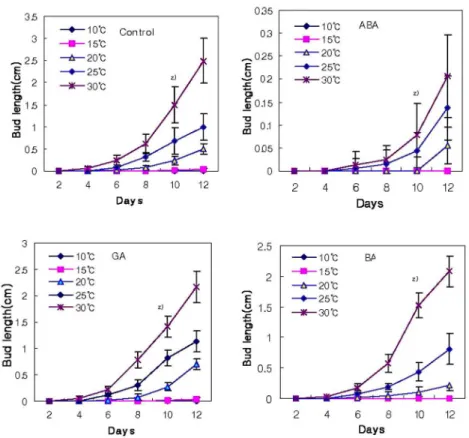
응을 보이지 않았으나, 15℃에서는 10일경에 맹아를 시작하였고, 20℃에서는 6일경 맹아를 시작하여 10일경에는 100% 맹아했다. 25℃에 서도 6일경 맹아가 시작되어 2일후인 삽목 후 8일경에는 100%의 맹아하였고, 30℃에서는 4 일경에 맹아하기 시작하여 6일후에 100% 맹아 하였다(Table 1). 또한 온도가 높아짐에 따라 액아의 맹아뿐만 아니라 생장도 촉진되었다 (Fig. 2).
100 mg/L GA 처리 경우 20℃와 25℃의 온 도에서는 6일째 맹아가 시작되어 삽목 10일 후 에는 100%, 30℃에서는 4일째에 시작되어 6일 에는 100%의 맹아했다(Table 1, Fig. 2). BA 50mg/L 처리도 10℃에서는 반응을 보이지 않 다가 15℃에서는 삽목 12일부터 맹아가 시작되 었고, 20℃에서는 6일 후에 시작되어 12일에는 75%정도의 맹아율을 보였으나 대조구인 무처 리구나 GA처리에 비하면 약간 저조하였다.
25℃는 6일 후에 시작되어 12일에 100 %를 나 타내었으나 무처리나 GA처리에 비하여 저조하 였다. 30℃에서는 6일에 100%맹아를 보여 무 처리나 GA처리와 유사한 결과를 나타내었다 (Table 1, Fig. 2).
ABA 200mg/L 처리의 경우 25℃이하의 온 도에서는 거의 맹아하지 않았으나, 25℃ 온도에 서는 6일 후에 맹아가 시작되어 12일 후에는 90%이상 맹아하였고, 30℃에서는 12일에 100% 맹아했다(Table 1). 액아의 생장도 무처 리에 비해 억제되었다(Fig. 2). Barros와 Neil (1986)은 버드나무(Salix viminalis) 휴면 연구 에서 내생 ABA의 양과 휴면과는 상관관계가 없고, 휴면중이거나 휴면에 들어갔을 때에만 ABA에 의해서 휴면이 유지될 수 있다고 했는 데 이 결과에 따르면 본 실험의 시기가 10월 중 하순경으로 아직 잎의 탈리현상도 일어나지 않 았을 때 시행하였으므로 이미 저온처리를 받았 다고는 생각할 수 없어 아직 휴면에 돌입하지 않은 상태이며, 휴면 호르몬인 고농도의 ABA 에 침지하였어도 맹아가 되는 것으로 보아 휴면 에 들어가지 않은 상태라는 것을 알 수 있었다.
Control
KOR. J. HORT. SCI. & TECH. 16(3), OCTOBER 1998 356
Treatment Bud sprouting ratio(%) Growth
regulators Temperature
(℃) 2z 4 6 8 10 12
Control
10 0 0 0 0 0 0
15 0 0 0 0 25 37.5
20 0 0 43.8 62.5 100 100
25 0 0 87.5 100 100 100
30 0 62.5 100 100 100 100
ABA 200mg/L
10 0 0 0 0 0 0
15 0 0 0 0 0 0
20 0 0 0 0 0 56.3
25 0 0 6.3 18.5 43.8 93.8
30 0 0 12.5 25 56.3 100
GA 100mg/L
10 0 0 0 0 0 0
15 0 0 0 0 25 43.8
20 0 0 50 68.8 100 100
25 0 0 62.5 100 100 100
30 0 56.3 100 100 100 100
BA 50mg/L
10 0 0 0 0 0 0
15 0 0 0 0 6.25 12.5
20 0 0 18.8 50 68.8 75
25 0 0 62.5 81.3 93.8 100
30 0 31.3 100 100 100 100
zDays after stem cutting.
Table 1. Effect of various temperatures and plant growth regulators on bud-sprouting
ratio of Hibiscus syriacus ‘Honghwarang’ during the 12-day period after stem cutting.Control
0 0.5 1 1.5 2 2.5 3 3.5
2 4 6 8 10 12
Da y s
Bud length(cm)
10℃
15℃
20℃
25℃
30℃
z)
ABA
0 0.05 0.1 0.15 0.2 0.25 0.3 0.35
2 4 6 8 10 12
Days
Bud length(cm)
10℃
15℃
20℃
25℃
30℃
z)
GA
0 0.5 1 1.5 2 2.5 3
2 4 6 8 10 12
Da y s
Bud length(cm)
10℃
15℃
20℃
25℃
30℃ z)
BA
0 0.5 1 1.5 2 2.5
2 4 6 8 10 12
Days
Bud length(cm)
10℃
15℃
20℃
25℃
30℃
z)
Fig. 2. Effect of various temperatures and plant growth regulators treatment on
the bud growth of Hibiscus syriacus ‘Honghwarang’ after stem cutting. ZMean±S.E.a z)
b b
ab a
0 0.2 0.4 0.6 0.8 1
2 4 6 8 10
Days after stem cutting
Cayalase activity (unit/mgprotein/min.)
↑
Fig. 3. Changes of peroxidase and cata-
lase activity of axillary bud of Hibiscussyriacus cv. Honghwarang under 25℃
growth chamber after stem cutting.
ZMean separation within row by Dun- can's multiple range test at the 5% level.
↑start bud sprouting.
즉, 무궁화는 다른 온대산 목본식물과는 달리 저온처리를 받지 않아도 맹아할 수 있다고 생각 되었다. Borkowska와 Powell (1892) 역시 사 과나무의 휴면연구에서 위와 같은 결과를 보고 한 바 있다.
25℃ 무처리구에서 맹아하는 동안 액아의 내 부물질 변화를 알아보기 위하여 POD와 CAT 활성을 측정한 결과 (Fig. 3), POD는 맹아가
진행될수록 감소하였으며, CAT는 맹아가 시작 되는 4~6일 사이에 감소하다가 완전히 맹아한 후 다시 증가하는 경향을 나타내었다. 현재 휴 면의 시작과 종결시 생장조절물질의 변화가 분 명하게 밝혀지지 않고 있으며 몇가지 질소화합 물이나 CAT활성의 변화에 대한 보고가 있다 (Omran, 1980). Nir 등 (1986)의 포도나무휴 면의 연구에서도 눈이 휴면상태에서는 CAT활
성이 높다가 저온 처리나 휴면타파물질을 처리 하면 활성이 급격히 감소하는데 이것은 눈 조직 내의 H2O2의 양이 증가함으로 일어나며, 증가 된 H2O2는 pentose-phosphate pathway를 활 성화시켜 맹아하는 것이라고 보고하였는데, 본 실험 무궁화의 경우에도 이 보고와 유사한 결과 를 나타내었다.
대부분 낙엽성 관목들의 눈은 가을에 형성된 후 온도가 bloom에 알맞은 상태라 할지라도 성 장하지 않는 것을 볼 수 있는데, Lang 등 (1987)은 이 단계를 endo-dormancy라고 구분 하여 확실한 저온처리에 의해서 타파된다고 하 였다. 온대산 식물이 일정기간의 저온처리로 휴 면이 타파(Ammon 등, 1979; Ammon, 1987) 되는 것과는 달리 무궁화는 저온처리 없이도 20℃ 이상의 온도에서 맹아가 빨리 일어나는 것으로 미루어 endo-dormancy상태에서도 이 상적인 환경만 주어지면 저온처리를 받지 않아 도 언제든지 타파가 가능한 휴면상태로 있음을 알 수 있다. Dennis(1987) 역시 endo-dor- mancy는 저온에 의해 유도되고 그 후 고온에 의해 상쇄될 수 있다고 하였다. 따라서 무궁화 의 휴면은 일정온도를 유지시켜주면 휴면기간 을 짧게 할 수 있거나 휴면이 없다는 것을 알 수 있으며, 앞으로 휴면단계에 따른 세포생리학 적인 연구가 뒷받침되어 이 결과의 확인이 필요 하며 이는 무궁화의 번식이나 이용, 연구에 큰 기초가 될 것으로 기대한다.
초 록
무궁화 (Hibiscus syriacus ‘Honghwa- rang’)의 휴면특성을 구명하고자 온도와 생장조 절물질을 처리하여 萌芽 차이를 비교하였다. 맹 아는 온도가 높을수록 빨랐으며(10~30℃), 3 0℃ 무처리구는 삽목 후 4일만에 맹아하기 시
Z
357 KOR. J. HORT. SCI. & TECH. 16(3), OCTOBER 1998
작하여 6일경에는 100% 맹아했다. GA와 BA처리도 대조구인 무처리와 큰 차이 없이 20℃
이상의 온도에서는 6일만에 맹아하여 62.5%의 높은 맹아율을 나타내었다.
ABA 침지처리는 25℃이하의 온도에서는 맹 아율이 저조하였으나, 25℃이상의 온도에서는 삽목 6일 후에 맹아하기 시작하여 12일 후에 93% 맹아되어, 고온 조건에서는 생장억제제인 ABA처리가 맹아억제에 전혀 효과가 없었다.
추가주요어: abscisic acid, benzyladenine, 휴 면, gibberellic acid,
인용문헌
Ammon, E. 1987. Chemical control of bud- break. HortScience 22:1240-1243.
Ammon, E., G.A. Couvillon, and C. H.
Hendershott. 1979. The effect of cycle length on chilling negation by high tem-
peratures in dormant peach leaf buds. J.
Amer. Soc. Hort. Sci. 104:573-576.
Barros, R.S. and S.J. Neil. 1986. Perio- dicity of response to abscisic acid in lateral buds of willow (Salix viminalis L.). Planta 168:530-535.
Borkowska, B. and L. E. Powell. 1982.
Abscisic acid relationships in dormancy of apple buds. Sci. Hort. 18:111-117.
Dennis, F.G. 1987. Two methods of study- ing rest : Temperature alternation and genetic analysis. HortScience 22:820- 824.
Kar, M. and D. Mishra. 1976 Catalase, peroxidase, and polyphenoloxidase activ- ities during rice leaf senescence. Plant Physiol. 57:315-319.
Lang, J.D., J.D. Early, and G.C. Martin, and R.L. Darnell. 1987. Endo-, para-,
and ecodormancy : Physiological termin- ology and classification for dormancy research. HortScience 22:371-377.
Nir, G., Y. Shulman, L. Fanberstein, and S. Lavee. 1986. Changes in the activ- ities of catalase in relation to the dor- mancy of grapevine (Vitis vinifera L.) buds. Plant Physiol. 81:1140-1142.
Omran, R.G. 1980. Peroxidase levels, and the activities of catalase, peroxidase and IAA oxidase during and after chilling cucumber seedlings. Plant Physiol. 65:
407-408.
Timson, J. 1965. New method for recoding germination data. Nature 207: 216- 217.
류달영, 염도의. 1974. 무궁화의 개화특성에 관 한 연구. 한원지 15:173-177.
류달영, 염도의, 김일중. 1975. 무궁화 개화특 성에 관한 연구. 한원지 16:106-113.