Kor. J App l. Micurbio l. Bioeng Vo!. 16 , M ι 1 , 17-24 μ988)
Saccharomyces
cerevisiae의 형 질 전 환박세경·정동효* 중앙대학교 산업대학 식품가공학과
Transformation of Saccharomyces cerevisiae
Park , Sei-Kyoung and Dong-Hyo Chung*
Department of Food Sc ience and Technology , College of Industry , Chung-ang University , Ansung 456-80,α Korea
Yeast ce lI s could be transformed only when their protoplasts were treated with plasmid vector.
However , preparation of protoplasts and their subsequent regeneration of ce lI wa lI are tedious and time consuming. Therefore , development of a more convenient method for yeast transformation has been needed. The transformation of intact yeast cells was attempted by treatment of various alkaline cations and electric field pulse. A Ii thium compound , such as LiSCN , was the most effective one of a lI alkaline cations tested , and the transformation frequency with LiSCN exceeded that abtained by the protoplast transformation. Conditions for the transformation of Saccharomyces cerevisiae DKD-5D with YRp 7 vector were investigated with LiSCN. The optimum concentration of Li+ and polyethylene glycoI4000(PEG) were O.lM and 35070 , respectively. The induction time was 60min and transformation frequenc y increased with increasing concentration of plasmid vector. PEG was ab- solutely required and the heat pulse treatment stimulated varied depending on the host-vector com- bination. An electric field pulse of intact yeast cells resu It ed in a marked improvementof the trans- formation frequency. The optimum field strength in the case of transformation of Saccharomyces cerevisiae DKD-5D and D13-1A with YEp 13 vector was about 50-70 vo It age for 3min , while higher field strengths were less effective. The optimum concentration of PEG was 20-30% , which was lower than the previous methods.
유전자 재조합기술(1)에 있어서 중요시되는 것은 재조합
plasmid
즉 vector이으로 고말현vector
흑 은 분비vector
등 역 러 유형 의 vector가 개 말되고 있다. 그러 나 아우리 우수한 vector를 개 말한다 하 여도 숙주세포의 형철전환을 안전하게 하지 않으면 숙주세포내에서의 발현은 기대할 수 없고 그러므호 목척산울을 얻을 수 없다. 또한 많은 비용과 노력으 로 마량 얻어지는 재조합 DNA를 손쉽고 효율적으로 숙주세포내호msertíon
시켜야 할 것이다.1978년에
Hinnen
등 (2) 은Le u 2
유전자 (3) 를 재 조합시 킨Col E 1
plasmid를 사용하여 효모의leu.
균주를
leu+
균주로 형질을 전환시컸다. 이 때 사용 한 형질전환법은 효모의 견고한 세포벽을 세포벽 용 해효소 (4, 5) 를 사용하여 일단 protoplast로 만든다 음plasmid
DNA를 msertlOn시키고 다시 세포벽을 재생시켜 주는 것으로서 일영 protoplast화에 의한 형질전환엽이라고 부른다. Hinnen의 형질전환법은 Beggs(6) 나Struhl
듯(7)에 이어져 효모중에서 자율Key words : Yeast transformation , Sacchaomyces cerevisiae , YEp13 , YRp7 , YCp19
*Corresponding author
17
18
증식 이 가능한 vector플 보고하였고, 효모의 숙주 -vector계 (SC 계) (8)가 확립되었다.
1982년에
Kimura
등 (9) 은 계면활성제로 처리된 효모의 막투과성의 변화를 검토한 바 DNA의 mcorporatton이 저빈도로 일어난다는 것을 발견하였 다. 이것을 토대로 protoplast화에 의한 형칠전환법 이 가지는 몇가지 단점을 보완하기 위하여 KU볍 즉, 알칼리 긍속이 온이 나Li
화합물로 처 리한 효모에 고빈도로 재조합 DNA를 incorporation시키는 새로 운 형 질전환법을 고안하였다.한편, 1978년
Weber
등 (10) 은 효모에 있어서polyethylene glycol(PEG)
존재하에protoplast
세포 의 융합벤도가 전기자극에 의해 상송된다고 보고하 였다. 1978년Senda
등 (ll) 은 식울세포의proto-
plast
배양세포에 전기자극을 주므로써 세포융합을성공시켰으며, 1982년
Neumann
등(1 2) 은 전기자극 에 의 해 서 DNA 가 mouse 의myeloma
세 포에 mcorporatlOn되는 것을 보고하였다.따라서 본 연구에서는 효므의 숙주-vetor 계를 통한 유전자 재조합 실험의 중요한 단계인 형질전환에 있 어서 안전하고 효율적인 방법을 규명하기 위하여 지 금까지 사용되고 있는 protoplast화에 의한 형질전환 법과 KU법에 관하여 알아보았다. 또한 더욱 빈도가 높으며 신속한 방법을 개발하기 위하여 천기적 자극 에 의한 세포융합과 식물
protoplast
배양세포에 DNA를 mcorporatlOn시키는 것에 착안하여 효모의intact
cell에 형질전환운 시도한 바 새로운 사실을발견하였으므로 그 결과플 보고하는 바이다.
Table 1. Li st of yeast strains and plasmids used.
Kor. f. App l. Microbio{. Bioeng.
재료 및 방법
사용균주 및
plasmid DNA
실험에 사용된 균주와 plasmid DNA의 특성은
Table
1 과 같다.형질전환 방법
Protoplast화에 의한 형질전환법은 Boggs(6) 의 방 법을 개정하여 사용하였다. 이 때
Protoplast
수율,재생효율은
Stewart
등 (17) 의 계산방볍에 준하였다.알칼리 금속이온,
lithium
화합물 처 리 에 의 한intact yeast
cell의 형질전환법 (KU볍)은Kimura
등 (14) 의 방법을 사용하였다. 즉 대수증식기 후기의 배양액(7.0X10
7cells/
ml) 을 원심분리하여 집균 후TE buffer (pH
7.5) 에 세척, 현탁하고 동량의0.2 M
알칼리 금속화합물 혹은 lithium염을 가하여30'C
에서 1시간 진탕하였다. 이 처리액 0.1ml를 새로운eppendorf
tube에 취 하고 여 기 에 2mg의 DNA가 함 유된DNA
용액을 첨가하여 30'C 에서 30 분간 incubation하였으며 동량의70% PEG
용액을 가하 여 30'C 에서 1시간incubation
한 후 42'C 에서 5분간 의 열처리 단계를 거쳐 형질천환하였다. 열균수로 세척하고 이 현탁액을 영양요구성에 따라 아미노산 이 첨가된 선택배지에 plating항요로써 30'C 에서2
-3일간 incubation후 나타나는 colony를 형질전환체 로 하였다. 이때 형질전환빈도는 금속이온 흑은lithium
염 처리한 후의 생균수에 대한 형질전환체의바로써 계산하였다. 한편 KU법의 개량법인 KUR볍
Strains & Plasmids Genotype & Characteristics Source & Reference Strain:
Sa cch. cerel ’ lszae
DKD-5D Dl3-1A DK-13D Plasmid vectors (kb):
YRp7(5.7)
YEp13( 1O.7) YCp19( 1O.6)
MATa , trp1, leu2-3, leu2-112 , his3
MATa , his3-532, trp1, ga12
MAT , trp1, his3, leu2
trp1 , ampr, tetr
leu2 , ampr ’ tetr
ura3, trp1 , leu4, ampr, tetr
Y. Oshima (1 3) Y. Oshima , (1 3) 1to (14) y. Oshima (13 , 15)
Y. Oshima (1 3)
L. Clarke (16)
Y. Kikuchi
L. Clarke (1 6)
lι 1. 16, Mι l
팎떻펀추 a
Fig. 1. Scheme of discharge circuit.
a: Ac 100V b: Power supply c: Fuse d: Platinum electrodes
은 KU법의 단계중에서 열처리 단계와 멸균수로 PEG를 세척, 제거하는 단계를 삭제한 일종의 신속,
간편한 방법이다.
전 기 자극
(electric field pulse)
에 의 한mtact yeast
cell의 형질전환은intact cell
자체에 전기자극을 주 는 방법으로 매우 간단하게 이루어 진다. 즉 대수증 식기의 효모배양액을 원심분리로 집균 후 열군수에 두 벤 세척하고 여기에DNA
용액을 첨가하고 최종 농도가 35%되 게PEG 4000
용액을 가하였다. 이 혼 합액에 백금전극을 맘그고 일정시간 방전하여 형질 전환시켰다. 수 분간 방치 후 이를 멸균수로 세척하 여PEG
용액을 제거하고 선택배지에 plating하여2
-3일간 30"C 로 incubation하여 나타나는 colony를 형질전환체로 하였다. 이때 사용한 방전장치는Weber
등(10) 의 전기융합장치를 토대로 하였으며회로는 Fig.1과 같다.
각기의 형질전환 실험에서 형질전환체의 확인을 위하여 형질전환체로부터
plasmid
DNA를 검출하였 는데 이 것 은Rodriguez
등(18)
의 방법 에 따라 얻은Table 2. Transformation frequncy of protoplast cells Viable cells per Protoplast yield
Strain ml (%),)
Sacch , cere
1!isiae
7.3 X 10 7 99.8 DKD-5D (YRp7)
Sacâι cere. !tsxae
6.9 X 10 7 99.5 Dl 3.1A (YRp7)
Sacch. cerevisiae
DK-13D (YRp7) 7.0 X 10 7 99.9 a) Protoplast yield
19
cleared
iysates를agarose gel electrophoresis
함으 로써 자외 선 조사시 나타나는 band로써 확인하였다.결과 및 고찰
Protoplast화에 의한 형질전환
Protoplast화에 의한 효모의 형질전환은 크게 3단 계 즉, 세포의 protoplast화,
Ca
2+ , PEG
존재하여plasmid
DNA의mcorporatlon ,
형 질 환된protoplast
의 세포벽재생으로 。l 루어 진다. 그러으로plasmid
DNA의mcorporatlon
빈도가protoplast
수율이 나 재생효율에 따라서 결정되는 것으로 생각된다.Table
2는Sacc h.
cerevisiae를 숙주로protoplast
수율, 재생효율 그리고YRp 7
vector의mcorpora- tlon
정도를 살펴본 것£로 일반적 으로protoplast
수 율은 세 균주 모두 세포벽이 용해되었음을 보여준 다.Protoplast화 되 는 기 작은 전 처 리 단계 중
thiol
화 합울이 세포벽 구성성분인 protem 에 존재하는disulfide
bond를 깨뜨리며 세포벽 용해효소 처리로세포벽이 용해되어 protoplast화 된다고 보고된 바 있다. 또한 protoplast화에 있어서 용해효소의 처리 정도,
protoplast
수율이 높을수록 재 생 빈도는 낮아 진다고Stewart
등 (18) 은 보고하였다.Table
2에서 도protoplast
수율이 높을수록 재생효율은 감소되 는 것으로 나타났다.그러 나 형 질전환빈도는 동일한
plasmid
DNA인Regeneration Transformants per Transformation efficiency (%)b) 2μg of DNA frequenc y<=)
4.8 413 5.7 x 10냉
1 1. 6 583 8 .4 X 10- 6
3 .4 626 8.9 X 10- 6
lni tial cell number-Non protoplast number P.Y.(%) = -"'-'-' -_ ..
"-~"-~'.- _ ..
r '-' - r ' - - ' . _ -X 100
lnitial cell number b) Regeneration efficiency
Regenerat ion cell number - Non protoplast number R.E . (%) =
--;o.~:':'-';,-"---- --_ ••• __ • -'-"
"._._"._~. "~"'V~'x 100
Initial cell number - Non protoplast number c) Transformation frequency Transformants per 2ug 0fDNA
Viable cell number
Kor. J App l. Microbiol
‘Bioeηg
뻐
1 , ‘
i
「 ,
•
、~、e{L
•‘’ l
l ' 3 LiSCN concentration αu
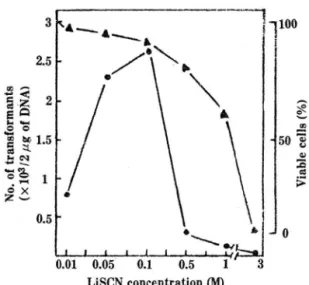
Fig. 2. Effect of LiSCN concentration on the transfor- mation of Sacch. cerevisiae DKD-5D with YEp13 vector and cell viability •
• - . Number of DKD (YEp13) transformants
‘ - ‘ Viable cells
(
-
s、
)
-
∞50
용-
.,Q..
g.- >
0
./F·\
3
2.5 2
1
'" '"""
-
동 들 a- z
... ‘
i-
。훌 폴1.5
뇨 N
~'O
,..‘
~O
X
ι)
0.5
YRp7 인 vector 로 mtegratlOn 시 켰을 때Sacc h.
cerevisiae DK-13 D , D 13-1 A , DKD-5 D
순a
로,protoplast
수율이나 재생빈도에 비례하 、 않는 것으로 나타났다. 그러표로 protoplast화에 의한 형질전 환에 있어서 그 빈도가
protoplast
수율, 재생효율에 따라 결정되지 않으며 숙주균과 plasmid간의 숙주 -vector계의 상호관련에 의하여 영향을 받는 것으로 사료된다.한편, protoplast에
plasmid
DNA가incorporation
되는 기작은 정확하게 알려지지는 않았£나proto-
plast에 PEG나 DNA가 형성하는 막전위의 변화에 기인한 것으로 볼 수 있다.20
모의 형질전환반도가 크게 상승하였다.
한편, 알캉라금속 특히
lithium
화합물은 소수결함 을 절단하는 힘 이 강하고 세포표면의 소수결합에 변 화를 주는 것으로 생각된다. 따라서 KU법에서는 먼 저 알캉리금속 등의 처리에 의하여 세포표면과DNA
에 변화를 일 a 켜 DNA의 incorporation이 쉽게 일어 날 수 있는 상태로 된다고 볼 수 있다.Li SCN
농도의 영향:
KU 법중 가장 형질전환이 잘 이루어지는 LiSCN의 최적농도는 O.lM로 나타났무 며 이 처리가 생존균의10%
정도를 사멸시키는 것 을 알 수 있다. Ito
등의 연구에서도 같은 일가 양이 온인Cs +, Rb+
이온O.lM
처리가 형질천환이 가장 잘 일어났으며 이때 이들 이온에 의한 사열율은 KU법에 의한 형질전환Table
3은 KU볍을 사용하여 몇가지 화합물을 선택하여 형질전환 빈도를 딸아본 것으로서
KC1 , CsCl
등의 화합물도intact
cell에 DNA를mcorpo
ration시 키 며, thiol
화합물인dithiothreitol , 2-mer.
captoethanol
등도 적은 빈도나마 형질전환이 얼어냥 을 볼 수 있다. Lithium염인 Li2
SO.와 LiSCN을 사 용하였을 때 다른 화합울보다 많은 형질전환체를 얻 을 수 있었으며 특히LiSCN
처리 경우에는DNA
μg 당10-
5 빈도플 나타내어 가장 효과적이었다.K+ ,
Cs+와 같은 1가 양이온은PEG , DNA
등과 복장한 전기적인 charge를 형성하여 세포의 업체적 구조변화와 응집 을 야기 함으로써DNA incorpora.
tlOn이 가능하다고
Kimura
등(9)
이 보고하였다.Kimura
등(9)
이 2-mercaptoethanol을 사용하여 효모를 형질전환시킨 결과를 보면 척은 빈도로DNA
의incorporation
이 가 능 하 였 다. 반 면2-mer.
captoethanol처 리 후 다시 lithium염 으로 처 리 한 효
Sacch. cere
l'isiae D13-1A (YRp7)
a b
Table 3. Effect of various compounds on transformation
\ \ Strains
DKD-5D(YRp7)
Treatment a b
DK-13D (YRp7)
a b
100 550 1121 2024 310 92 9.2 X 10 7
7.3 X 10 7 9.5 X 10 7 9.1 X 10 7 7.2 X 10 7 8.2 X 10 7 130
355 465 985 295 74 8.2 X 10 7
7.5 X 10 7 8.1 X 10 7 6
‘2 X 10 7 5.7 X 10 7 8.0 X 10 7 151
168 1210 2650 109 185 5.5 X 10 7
4.0 X 10 7 8.5 X 10 7 8 .4 X 10 7 4.7 X 10 7 3
‘5 X 10 7 KCI
CsCl Li2S04 LiSCN Dithiothreitol 2-Mercaptoethanol
a: Viable cells per m/ after the treatment of various compounds
b: Transformants per 2μg of DNA
V{ i l. 16 , Mι 1 21
.,‘-‘
j /\ • r •\ \
.. . \ ‘ l
I 、、
•
I • ‘
/ \
.l--L.- .
o 20 40 60 80 100 120140160 Induction time (min)
Fig. 4. Effect of induction time on the transformation of Sacch. cerevisiae DKD.5D with YEp13 vector .
• - . Number of DKD-5D (YEp13) transfonnants
‘ -‘ Transfonnation frequency
10- 4
‘〔 gζgil
〕!}』}ζ@;
∞E』S ∞를여』← -녕
F애
m
m
3
-꾀
5
--aι (북
Za }。
Ma
N
감-{× )
@를혀
E‘。‘∞
Z∞뇨 등
.。z
2 2.5
1 10- 4
>‘
g
a
@
a cr 10- 5
효걷 。
....
<=
틀
‘.
。
“
∞ Ia 10- 6
얻3
2.5
픈육 ~~
들 z
듣 a
L
‘
i
。흩 양
1. 5
될얀
응음。 × -.,,-. -
2
0.5
vector를 사용한 결과 Fig.4에서와 같이
induction
tU1'le에 따라plasmid DNA incorporation
정도에 큰 차이를 나타냈a며 최적 시 간은 60분무로 나타났다.그러므로 일단
LiSCN
처리플 하여competent cell
로 만들어 졌다해도 DNA와 PEG와 상호작용으로 숙주균으로plasmid
DNA가mcorporat lC n
되는데 일정시간이 소요됨을 알 수 있다.Plasmid DNA
농 도 의 영 향Sacc h. cerevisiae
DKD-5D에
YEp 13
vector플 농도별로incorporation
>‘ 9
-
..,
i
cr
..,
‘ t
t
10- 5 1
응g
E
i‘
。g。i ζ<=
~
10- 6
50%로써Li+
이온보다 생존균에 미치는 영향이 크다는 것을 알 수 있다.
LiSCN
농도가0. 01%
미만에서는 그 농도가 낮아 서pl asmid
DNA가 도입 될 수 있는competent cell
로 만드는데 우리가 있는 반면1M
이상의 고농도에 서는 균주의 90% 가 사멸되기 때문에 당연히 형질전 환빈도도 낮은 것으로 생각된다.PEG 4000
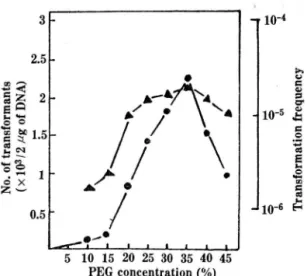
농도의 영 향:
Fig.3은plasmid
DNA로 형질전환시키는데 PEG를 첨가하지 않은 경우에 전 혀 형질전환체를 얻을 수 없는 것 A로 나타나PEG
가 필수적임을 보여주며,35%
PEG처리가 반도면에 서 효과적임을 나타내는데 이는Ito (l 9)
등의 결과와 일치한다.PEG는 탈수작용에 의하여 세포내외의 산화환원
전위의 변화를 유말하여 숙주내로
plasmid
DNA를 도입하거나 또는, 음으로 하전되어Li+
이온과DNA
용액 그리고 세포표면사이의 복잡한 전기적charge
를 형성하여 세포의 구조변화와 응집현상이 일어나 서 이 원리로plasmid
DNA의 mcorporatIOn이 가능 하다는 두 가지 작용을 추정할 수 있다.PEG
농도40%
이상이 되면 자체가 가지는 세포독성과 심한 응집현상£호 형질전환빈도가 낮아진다고 할 수 있다.
/:_4- / ‘
7. ’
。그/
3
2.5 2
‘-‘ • -、‘
‘ / ·’ \ \
jf 진
“ .
d二3
/
5 10 15 20 25 30 35 40 45 PEG concentration (%)
Fig. 3. Effect of PEG concentration on the transforma- tion of Sacch. cerevisiae DKD-5D with YEp13 vector .
• - . Number of DKD (YEp13).transfonnants
‘ -‘ Transfonnation frequency
'" -‘
: ‘컨
Oi Z 5Q
』
“
s
。뚱 약 1.
5
여 “
.... ""
- -
‘~.,
。 o
,...
므 ×
• )
o 2 4 6 8 10 Plasmid vector concentration (μg)
Fig. 5. Effect of plasmid vector concentration on the transformation of Sacch. cerevisiae DKD-5D with YEp13 vector .
• - . Number of DKD-5D (YEp13) transfonnants
“ Transfonnation frequency Induction
time의 엄 합:
KU업 에 서competent cell
에 DNA와 PEG를 첨가한 다음 얼정시간 30.C에서 incubation하게 되 는데 이 기 간을
inductir;n
time이 라고 한다 Sacch.cerevisiae DKD-5
D에YEp 13
22
Table 4. Effect of PEG and heat pulsc treatment on the transformation
PEG treatment Heat pulse Transformants per 2μgof DNA
+ + 2120
+ 1250
+ 0
0 Transformation was carried out with YEp 13 vector of Sacch.
cerel때iaeDKD-5D. Other transformation condi- tions were as discribed in the tex t.
시켜서 형질전환빈도를 살펴보았다 (Fig.
5).
그 결과DNA
농도가 증가함에 따라서 형질전환빈도도 거의 비례적으로 증가되었다.PEG와
heat pulse
treatment의 영향:
형 질전 환과 정중에 포함되는PEG
첨가와 열처리단계의 필요성 과 상호관련성을 조사하였는데(Table 4) Table
4에 서 같이 PEG를 첨가하지 않s!-.연 거의 형칠전환이 얼어 나지 않았으며 또 PEG를 첨 가한 후 420C 에서5
분간 열처리조작을 통하여 그 반도가 배로 상승됨을 알았다.Kimura
등의 실험에서도 실제 DNA가 균체 내로 도입되는 것은 PEG플 첨가한 후에야 비로서 가능하다고 보고하였다. 따라서 KU 볍에 있어서LiSCN
처 리 는competent
cell로 만을고 DNA가 숙 주균내로incorporat lO n
되는데는 PEG가 필수적이며 열처 리 플 통한heat
pulse는PEG
작용을 활성 화시 켜 그 빈도를 크게 높여주는 것으로 사료된다.KU법과 KUR법의 비교
:
KU법에 이어 개량법우로 KUR법 이 보고되 였는데Table
5는 이 두 방법을 통 한 형질전환 빈도플 비교하여 본 것이다.Ha-
Table 5. Comparison of transformation frequency bet- ween KU and KUR method
Strain KU KUR
a b a b
1785 2.6 x 10- 5 630 9.2 x 10냉
DKD-5D (YRp7) Sa cch. cerevisiae
1250 1. 8 x 10- 5 869 1. 3 X 10- 5 D1 3-1A (YRp7)
Sa cch. cerevisiae
1650 2 .4 x 10- 5 265 3.9 X 10 -6 DK-13D (YRp7)
a: Transformants per
2.μgof DNA b: Transformation frequency
]'ransformats per 2μg of DNA - Viable cell number after LiSCN treatment
KIIχ
J App l. Microbio l.
Bi"eηιshimoto
등 (13) 은 PEG의 세포독성이 그리 크지 않 아서 세포와PEG
흔합액 그대로을 직접 선택배지에 plating하여도 KU법과 비교하여 그 빈도는 같은 정 도이거나 그 보다 높다고 하였으나, 본 실험결과에 서는 간펀하고 손쉽기는 하나 그 빈도면에서 KU법 보다 떨어졌 q~Table5).
Plasmid pNA
종류에 따른 형칠전환빈도: Table 6
에서와 까ε! 형질전환빈도늦 모두10-
5 반도를 보였t
는데 YCp19가YEp 13, YRp 7
vector보다는 약간많은 형 질전환체를 얻을 수 있었다. 이 것은
It o
연구 와는 약간의 차이를 보여서 YRp7이 분자량이 작아 서 형질전환이 잘 일어난다고 한 결과와는 일치하지 않았다. 효모를 숙주로 한 여 러 연구(1 9, 20, 21)
에 서 형질전환빈도를 살펴보면 동열한 조건에서 동열 한 vector를 사용하였다하더라도 차이가 있음을 볼 수 있다.Table
6에서도 반도로 따지면 큰 차이 라고는 할 수 없지 만 같은 유형 의 vector라 하더 라도 숙주균에 따라 그 반도가 을려지는 것으로 보아 숙 주균과 vector의 상호관련에 의한 영향이 더욱 지배 적이며, 따라서 효모의 형질전환시 host-vector계의 선딸이 매우 중요하다고 할 수 있다.전기자극 (electric
field
pulse) 에 의한 형질전환Plasmid DNA
종류에 따른 형질전환빈도Weber
등 (10) 은 전기자극을 통하여 효모 protoplast의 세포 융합도를 상승시켰으며 그는 이것을 이용하여 transformation도 가능하리 라고 보았다. 또한 KU법 에서도 아직 빈도가 낮아서 더욱 효과적인 형칠전환Table 6. Transformation frequency of LiSCN treated Sacch. cerevisiae cells with different plasmids
Plasmid Transfor- Transforma- Strain vector mants per tion fre-
2μgof DNA quency' DKD-5D YRp7 2010 2.6 X 10- 5 DK-13D YRp7 1650 2.0 X 10- 5 D13-1A YRp7 1240 1. 8 X 10- 5 DKD-5D YCp19 1640 2.1 X 10- 5 DK-13D YCp19 3110 3.8 X 10- 5 D1 3-1A YCp19 2605 3.7 X 10- 5 DKD-5D YEp13 2085 2.7 X 10- 5 DK-13D YEp13 2680 3.3 X 10- 5
Transformants per 2.μg of DNA
Viable cell number after LiSCN treatment
vector의 도입 정도는 꽤 높아서
10-
4 빈도를 나타냈 다.이 방법은 protoplast화에 의한 형질전환이나
KU
볍도 그 빈도연에서 동일하거나 그 이상으로 실험조 작이 간편하고 매우 신속한 장정을 가지고 있다. 전 기자극과plasmid
DNA의 세포내로의incorporation
과의 관계는 아직 병확하지는 않으나 전기자극시 세 포막의 변화에 대 한Morigawa
등(22)
의 총설을 창 조로 하면 전기자극으로, 첫째 막의 흥분현상이 야 기되어 막의 conformation이 변화하고, 둘째 막의 물질투과성이 증대하며, 셰째 막부근의ATP
합성이 활성화된다고 하였다. 따라서 이들 요인들에 의하여 DNA 가 세포내로 도입된다고 볼 수 있 a 나 정확한 mechanism에 관한 규명 이 있어 야 할 것 이 마.Electric discharge(
voltage)에 의 한 영 향:
방 전 voltage에 따른 형칠전환빈도 차이와 생존균에 미치 는 영향을 살펴본 결과를 Fig.6에 나타내었다.Fig.6에 서 와 같이
Sacc h. cerevisiae DKD-5
0는50V
부근이, DK-13D의 경우에는75-100V
영역 이 가장 형질전환빈도가 높았으며 이때 방전으로 인 한 사옐율은 10-20% 이었다.PEG 4000
농도의 영향 Protoplast화에 의 한 형 질전환과 KU엽에서 PEG의 존재는 필수적이있다.그러나 Fig.7에서와 같이 전기자극에 의한 형질전환 시
PEG 4000
의 최 종 농 도 는Sacch. cerevisiae
DKD-5D가
25% ,
DK-13D는25-30%
그리고D 13-1
A가 20%로서 앞서의 방법보다10%
정도가 낮V ,,!. 16 , N". 1 23
Table 7. Transformation frequency 01 various plasmid vector by electric field pulse treatment
Transfonna- tion fre- quency*
Transfor- mants per
2μg of DNA Plasmid
vector Strains
Sa cch cerevzstae DKD-5D DK-13D D13-1A DKD-5D DK-13D Dl3-1A DKD-5D DK-13D
2.0 X 10- 5 3_2 X 10- 5
8_3 X 10- 5 7_3 X 10- 5 1. 7 X 10
-41. 5 X 10
-45_2 X 10- 5 3 .4 X 10- 5 1840
1980 4820 6720 7200 8920 4810 2110 YRp7
YRp7 YRp7 YCp19 YCp19 YCp19 YEp13 YEp13
법이 필요로 되는 것은 물론이다.
Table
7은 방전장치를 사용하여50V ,
6mA에서 3분간 백금전극을 담가서 방전하여Sacc h. cerevisiae DKD-5D , 013-1 A , DK-13
D에 각 vector를incor- porat lO n
시킨 결과이다. 대부분10-
5 빈도플 나타내 고 특별 히DK-13
0와0 13-1 A
군주에YCp19
Za
(에6
‘-%
~ a S{
X -ιiZ@E 』()‘∞ ‘‘ )
Zgi-·@Z 4
Transfonnants per 2.μg of DNA
Viable ce l1 number after electric field pulse treatment (50V , 3min)
100
•
。/' ‘
~
n
/4g/·
gy/
L _
J 5 10 15 20 25 30 35 40 45 PEG concentration (%)
Fig.7. Effect of PEG concentration on the transforma- tion of Sacch. cerevisiae by electric field pulse treat- ment.
。-。
e-e
“-..,‘
4
F
。h ‘
- - - - l . . --e ‘ - 25 50 75 100 125 150 175
Eledric discharge (V oltage)
Fig. 6. Effect of electric discharge (voltage) on the transformation of Sacch. cerevisiae and cell viability.
e - e Number of DK-13D (YEp13) trans(onnants
。-
0 Number of DKD-5D (YEp13) transfonnants
‘ -‘ Viable cells of DK-13D
6. - 6.
Viable cells of DKD-5D
4
3
(육Z 디 강
야그;-’
NX )
을5*E』응∞
za뇨응
.。z ($)
을응 잉효영〉
50 5
3
2
0 0
Number of DKD-5D (YRp7) transfonnants
Number of DK-13D (YRp7) transfonnants
Number of D13-1A (YRp7) transfonnants
24
은 것이다. 또한 PEG를 전혀 첨가해 주지 않은 것 도 형질전환이 적은 빈도나마 이루어 진 것으로 보 아서 종전의 방법을에서 필수척이던 PEG의 역할이 직접적인 전기방전으로 달성된 것으로 고려된다.
본 실험 결과 전기방전에 의한 형질전환은 반도면 에서 기존의 방법보다 우수하고 조작면이나 시일변 에서도 유리한 것으로 나타났다. 앞으로 완전한 장 치를 제작하고 또 천처리로서
lithium
염 처리 응을 병행한다면 그 효율은 매우 상승될 것무로 믿는다.요 。 t--,
효모의 숙주-vector계플 통한 형질전환에 있어서 지금까지 사용되고 있는 protoplast화에 의한 형질전 환엽과 KU 법에 관해 알아보았다. 또한 전기자극£
로
intact yeast
cell에 형질전환을 시도하였다.Protoplast화에 의한 형질전환은
DNA
μg당10-
6 빈도가 나타났무며protoplast
수율, 재 생효율 등에 형 질전환이 영 향을 받는다기보다는 숙주-vector계의 상호관련성이 지배적임을 나타났다.KU 법에 의한 형질전환은
KCl ,
CsCI과 같은 일가 양이온과 lithium염 그리고thiol
화합물을 사용하였 는데LiSCN
처 리 가 가장 효과적 이 었다.LiSCN
농 도O. 1 M , PEG 4000
농도35% , induction
tine은 60분, 그라 고42
0C
5분간의heat pulse
treatment를 통하여 가장 높은 빈도를 나타났다. KU볍에 있어 PEG의 존재는 필수적이며 KUR법보다 KU법에 효 과적이고 또한 숙주의 특성에 따라 빈도가 차이플 보였다.전기자극에 의한 형질전환은 기존의 방법보다 조 작이 간편하며 그 빈도가 동일하거나 높은 것으로 나타났다. 방천 voltage에 따라 숙주균간에 약간의 차이 를 보였는데
Sacc h. cerevisiae DKD-5
D는50 V , 3min ,
DK-13D의 경우75-100V ,
3min이 가 장 형질전환반도가 높았으며 이때 방전으로 인한 사 멸율은 10-20% 이었다. 또한 PEG의 최적농도는 DKD-5D가25% ,
DK-13D는25-30%
그리 고013
-IA의 경우 20%로 종전의 방법보다10-15%
낮 거l 나타났다.Kor. j. App l. ,' vlicrobio l. Bioeη£·
참고문헌