이온빔을 이용한 미생물의 균주 개량
정해영*·김계령1
한국생명공학연구원 바이오화학/에너지연구센터, 1한국원자력연구원 양성자기반공학기술개발사업단
Strain Improvement Based on Ion Beam-Induced Mutagenesis. Jeong, Haeyoung* and Kye-Ryung Kim1. Industrial Biotechnology & Bioenergy Research Center, Korea Research Institute of Bioscience and Bio- technology (KRIBB), Daejeon 305-806, Korea, 1Proton Engineering Frontier Project, Korea Atomic Energy Research Institute (KAERI), Daejeon 305-353, Korea − For decades, traditional mutation breeding technologies using spontaneous mutation, chemicals, or conventional radiation sources have contributed greatly to the improvement of crops and microorganisms of agricultural and industrial importance. However, new mutagens that can generate more diverse mutation spectra with minimal damage to the original organism are always in need. In this regard, ion beam irradiation, including proton-, helium-, and heavier-charged particle irradiation, is considered to be superior to traditional radiation mutagenesis. In particular, it has been suggested that ion beams predominantly produce strand breaks that often lead to mutations, which is not a situation frequently observed in mutagenesis induced by gamma-ray exposure. In this review, we briefly describe the general prin- ciples and history of particle accelerators, and then introduce their successful application in ion beam technol- ogy for the improvement of crops and microbes. In particular, a 100-MeV proton beam accelerator currently under construction by the Proton Engineering Frontier Project (PEFP) is discussed. The PEFP accelerator will hopefully prompt the utilization of ion beam technology for strain improvement, as well as for use in nuclear physics, medical science, biology, space technology, radiation technology and basic sciences.
Key words: ion beam, proton beam, mutation breeding, irradiation, mutagenesis
서 론
방사선을 생명공학 분야에서 사용하기 시작한 것은 유전 자의 구조와 복제 및 발현에 대한 분자적 이해가 이루어지 기 훨씬 이전부터였다. 1927년 Muller가 초파리를 이용한 실 험을 통해 방사선이 생명체에 돌연변이를 일으킨다는 사실 을 처음으로 보고하면서 큰 반향을 불러일으키게 되었고[23], 1928년 Stadler가 엑스선과 감마선에 의한 보리의 돌연변이 유발을[36] 재차 확인한 이래 식물 육종학자들은 방사선을 이용한 돌연변이 육종을 통해 유용한 작물의 개량에 노력해 오고 있다. 2007년 현재 2,500종 이상의 돌연변이 품종이 국 제연합과 국제 원자력 기구에 공식 등록되어 있으며, 이 중 에서 약 3/4은 감마선 조사에 의해 유발된 것들이다[1]. 반 면 동식물에 비해 상대적으로 유전체 해독과 제어 및 인공 배양이 용이한 미생물 분야에서는 유용한 효소나 일, 이차 대사산물, 또는 생체량(biomass) 자체를 얻기 위하여 더 다 양하고 고도화된 방법을 적용할 수 있다. 벡터에 클로닝된 단백질이나 효소를 개량하고자 하는 경우 error-prone PCR
이나 DNA shuffling[37]을 통해 다양성을 유발하여 라이브 러리를 만든 뒤, 이를 대용량 스크리닝 시스템과 연결하여 반복적으로 우수한 돌연변이체를 선별하는 방향적 진화 (directed evolution) 방법을 예로 들 수 있으며[28], 단백질 의 입체 구조를 고려한 돌연변이체의 설계도 가능하다[24].
단일 DNA construct를 세포에 도입하는 전통적인 클로닝 기 반 유전자 조작 기술 이외에도 게놈을 직접 조작하여 유전 자를 필요에 따라 삽입, 치환 또는 제거하거나[4, 6, 26, 33, 43], 합성 올리고뉴클레오타이드를 이용하여 게놈에 동시 다 발적인 다양성을 유발할 수 있는 engineering 기법도 등장하 였다[41]. 뿐만 아니라 데이터와 in silico 분석 결과를 통합 하여, 세포내의 대사흐름 최적화를 통해 목적 산물을 최대 수율로 생산하거나[5, 27], 더 나아가서는 시스템 생물학적 접근을 통해 균주 개량과 생산공정 개선을 꾀하려는 시도에 도 관심이 모아지고 있다[25].
방사선 조사와 같이 무작위적으로 유발되는 돌연변이를 이용한 균주 개량 방법은 사전에 목적 균주를 합리적으로 설 계한다거나 개발 과정을 단축하는 데에 근본적으로 어려움 이 있지만, 적용 방법이 비교적 단순하고 조사 설비를 갖추 고 있으면 표준화된 프로세스에 의해 대량 시료의 적용이 가 능하며, 유전체 정보가 아직 공개되지 않았거나 개량하고자 하는 형질과 관련된 유전자 정보가 알려지지 않은 균주의 개
*Corresponding author
Tel: 82-42-860-4237, Fax: 82-42-879-8595 E-mail: [email protected]
량에 유용하다. 특히 유전체 상에 항생제 내성 유전자와 같 은 marker를 남기지 않으며, 자연계에서 일어나는 진화 과 정과 유사하게 균주의 개량이 일어난다는 점이 장점이다. 최 근에는 대사공학적 방법으로 개량된 균주를 최적화하기 위 하여 오랜 기간 동안 연속 배양을 하면서 자발적으로 발생 한 고성장 변이체를 선별해내는 이른바 adaptive evolution 에 대한 관심이 증대되고 있다[2, 7]. 이 방법을 통하여 일 반적인 대사공학적 접근 방법으로는 해결하기 어려운 고성 장 균주, 또는 특정 물질에 대한 내성이 증대된 균주를 선별 할 수 있다. 만약 방사선에 의한 무작위적 돌연변이 유발 과 정을 여기에 첨가한다면 이러한 개량 과정을 더욱 단축시킬 수 있을 것이다.
최근에는 다양한 원리의 입자 가속기가 개발되면서 감마선 이나 엑스선과 같은 전자기파 방사선뿐 아니라 양성자나 중 성자와 같은 핵자 또는 중이온(헬륨보다 무거운 원자를 방전 시켜 전자를 제거한 뒤 얻은 양이온)을 가속시켜 얻는 입자 방사선 등 다양한 방사선원을 본연의 업무인 핵 및 입자 물 리학이나 암 치료 이외에도 유용 생명 자원의 개발 용도로 사 용하게 되었다. 본 총설에서는 입자 가속기의 원리와 역사를 간략히 알아보고, 가속기를 통해 인출되는 고속의 이온들, 즉 이온빔을 이용한 생물자원의 개량 현황을 소개하고자 한다.
입자 가속기의 원리
이온빔의 발생장치로서 가장 중요한 것은 전기를 띤 입자 에 전자기장을 가하여 속도를 높인 뒤 빔 형태로 인출하는 장비인 입자 가속기(particle accelerator)이다. 1920년대에 물 리학자들은 입자 방사선으로 원자핵을 타격하여 그 구성 입 자를 밝히고자 많은 노력을 기울였으나, 자연 방사선원에서 나오는 알파(헬륨 원자핵)나 베타 입자(전자)는 그 수가 너 무 적고 에너지가 낮아서 핵의 전기장 장벽을 넘을 수 없었 으므로 다른 방법으로 고속의 입자를 만들어내는 방법을 강 구하기 시작하였다. 1931년 미국의 Lawrence가 개발한 최 초의 입자가속기는 직경이 4.5인치에 불과하였지만 1,800 볼 트의 전압을 걸어서 수소 이온을 빛의 속도의 1%에 해당하 는80 KeV로 가속하는데 성공하였다(Fig. 1). 후에 싸이클로 트론(cyclotron)으로 명명된 이 장비는 핵물리학이라는 새로 운 연구 분야를 개척하는데 큰 공헌을 하였으며, Lawrence 는 이 업적으로 인해 1939년 노벨 물리학상을 수상하였다.
이 장비는 현재에도 양성자 및 중이온(중입자)을 가속하는데 널리 쓰이는 원형(圓形) 가속기의 원형(原型)이 되었다.
입자 가속기는 하전 입자의 종류(양성자, 중입자 또는 전 자), 하전 입자를 가속하는 방법(직류 또는 교류)과 형태(선
Fig. 1. Particle accelerator examples: (A) The first cyclotron built by Ernest Lawrence, which had a diameter of only 4.5 inches; (B) an MC-50 cyclotron, a 50 MeV cyclotron of the Korea Institute of Radiological & Medical Sciences (KIRAMS), which is the larg- est high-energy proton accelerator in domestic operation except for a 230 MeV cyclotron used only for cancer therapy at the National Cancer Center; (C) a 150 keV gas ion beam implanter developed by PEFP (Proton Engineering Frontier Project); (D) a proton accelerator research center scheduled for construction in Gyeonju, Gyeongsangbuk-do by 2012, where a 100 MeV, 20 mA large-current proton pulse beam generator built by PEFP will be installed (the 20 MeV proton beam is presently in operation and a 90 MeV accelerator was also successfully developed at the end of 2009); and (E) a schematic drawing of the proton beam line under development by PEFP.
형 또는 원형)에 따라 여러 가지로 나뉘며, 인출할 수 있는 빔의 에너지와 빔 전류 및 통제 용이성 등의 특성이 모두 다 르다. 가속 입자의 활용성은 에너지에 따라 매우 다양한데, 예를 들면 이온 주입기(ion implanter)에서 발생하는 수십 KeV의 입자는 물질 속에 주입될 수 있으며, 재료의 표면 개 질이나 반도체 도핑에 쓰인다. 수십 MeV의 가속 입자는 핵 반응을 유발할 수 있고 돌연변이 육종이나 의료용 동위원소 생산에 쓰이게 되며, 가속 에너지가 GeV 단위에 이르게 되 면 원자핵을 구성하는 소립자와 반응이 가능하여 원자의 기 본 단위를 탐구하는 중간자/중성미자 과학에 응용될 수 있다.
이온 주입기(ion implanter)는 원래 실리콘 웨이퍼와 같은 고체 표면에 침투시킬 목적으로 불순물 이온의 고속도 빔을 생산하는 저에너지의 입자 가속기인데, 침투 길이가 짧기 때 문에 돌연변이 유발용으로는 거의 쓰이지 않고 있었다. 그 러나 1980년대 중반부터 비롯된 중국 과학자들의 노력으로 식물과 미생물의 육종 및 유전자의 전달 목적으로 쓰일 수 있음이 알려지게 되었다[44]. 이는 중이온을 이용한 식물체 의 개량[39]과 더불어 이온빔을 이용한 육종의 대표적인 성 공 사례가 되고 있다.
노벨 물리학상 수상자 중 약 20%가 가속기에 관련될 정 도로 입자 가속기는 물리학 연구의 매우 중요한 도구인 동 시에 생명과학, 의학, 핵물리, 나노산업 등 그 응용분야가 점 차 확대되고 있다. 의학 및 생명공학 분야만을 살펴보아도 의학용 동위원소 생산, 암 치료, 방사선 생물학 및 육종학 등에 널리 응용되고 있다. 특히 양성자는 우주선(cosmic ray) 의 90% 이상을 차지하고 있는데[30], 지구의 자기장에 의해 차폐가 되므로 지상까지 도달하는 양은 우려할 만한 수준이 되지 않는다. 그러나 우주에서 임무를 수행해야 하는 우주 비행사의 경우 적절한 보호 대책을 세우지 않는다면 최대
~1020 eV에 이르는 초고에너지의 우주 방사선에 의해 생명 이 위험할 수 있다. 이는 인류가 건설한 가장 큰 입자가속기 에서 만들어내는 양성자에 비해 약 1천만 배에 달하는 막대 한 에너지에 해당한다. 따라서 현재 실현 가능성이 논의되 고 있는 행성 간 유인 우주선 탐사 계획을 성공적으로 실현 하기 위해서는 입자 가속기를 이용하여 고에너지의 양성자 빔이 생물체와 전자기기에 미치는 영향을 미리 파악하고 대 응책을 마련하는 것이 필수적이다. 어쩌면 우주선은 화학 진 화의 시기를 이끌어서 지구 상에 생명체가 탄생하게 만든 가 장 중요한 외부적인 요인이었을 수도 있으며, 따라서 생명 자원의 개량에 양성자 이온빔을 이용한다면 지구 상에서 실 제 일어났던 것과 유사한 방식의 진화를 가속하여 적용한다 는 의미를 부여할 수 있을 것이다.
대용량 가속기의 경우 건설과 운용에 막대한 자금이 소요 되는 거대 과학의 대표적인 장치로서 그 파급효과가 막대함 에 따라 과학기술분야 국력의 척도로 인식되고 있다. 우리 나라의 경우 1994년 포항방사광가속기(http://pal.postech.
ac.kr/)가 건설되면서 세계 10대 가속기 보유국의 반열에 오
르게 되었으며, 한국원자력연구원의 양성자기반공학기술개 발사업단(PEFP: Proton Engineering Frontier Project, http:/
/www.komac.re.kr/)에서는 2012년 완공을 목표로 경상북도 경주시 건천읍에 100 MeV급의 대용량 선형 양성자 가속기 를 건설하고 있다. 또한, 정부에서는 과학비즈니스벨트의 핵 심 연구 시설로서 중이온 가속기의 건립을 계획하고 있다.
이온빔의 특성
일반적으로 에너지선은 표적 분자에 직접 손상을 주거나 (직접 작용), 세포 내의 물을 전리시켜 이로부터 발생한 라 디칼에 의한 간접 작용을 통해 생체 내 분자에 영향을 미친 다. 감마선이나 엑스선과 같은 일반적인 방사선은 주로 간 접작용에 의해 생체분자에 작용하지만, 이온빔은 직접작용 의 역할이 대부분을 차지하는 것으로 알려지고 있다. 방사 선의 세포 내 표적 물질로서 가장 중요한 것은 DNA로서, 단일 가닥 절단(single strand break, SSB), 이중 가닥 절단 (double strand break, DBS), 염기의 손상 혹은 손실, 그리고 분자내/분자간 가교 형성 등의 손상을 유발할 수 있으며, 특 히 밀집된 손상(clustered damage)이 중요하게 여겨지고 있 다[9, 12, 34]. 이 중에서 사실상 복구가 불가능하여 세포 사 멸을 유도하는 가장 중요한 손상은 DSB로서, 방사선 치료 과정에서 암세포를 죽이는 주된 메커니즘이 된다. 하지만, 물질이 같은 정도의 선량(dose)을 흡수했다 하더라도 방사선 의 종류가 다르면 작용이나 효율이 달라지는데, 이와 같이 비적을 따라 일어나는 전리 및 여기의 분포상태를 선형 에 너지전달(LET, linear energy transfer)로 나타낸다. 저 LET 방사선으로 대표적인 것은 엑스선과 감마선이며, 알파선, 중 성자, 중입자 등은 고 LET 방사선으로 분류된다. 일반적으 로 양성자빔은 저 LET 방사선으로 간주되지만 생체분자에 대한 영향력은 매우 큰 것으로 알려져 있다.
입자 가속기에서 발생되는 이온빔은 다른 방사선과는 달 리 물질에 가하는 에너지가 대단히 크고 조사의 위치와 심 도의 조절이 가능하다는 장점이 있다. 엑스선은 조직에 침 투하는 즉시 대부분의 에너지를 전달하지만, 이온빔은 시료 에 침투하였을 때 속도가 느려지고 궤도 전자와 상호작용이 증가하면서 최종적으로 양성자가 멈추는 지점에서 최대의 에너지를 주변에 전달하는 Bragg peak 특성을 보인다(Fig.
2). 따라서 선량의 집중화가 가능하므로 주변 조직의 파괴를 최소화하면서 생체 조직 내의 특정 위치를 선택적으로 타격 할 수 있다. 이러한 특성으로 인하여 이온빔의 일종인 양성 자빔에 의한 암 치료가 우리나라를 비롯하여 전 세계적으로 확대되고 있으며, 탄소를 포함한 중이온 가속기 또한 치료 용으로 쓰이고 있다.
최근에는 전산학적 모사에 의해 이온빔 또는 이온화 방사 선이 세포를 타격할 때 DNA에 나타나는 손상의 정도를 strand break의 수치로서 예측하는 것이 가능할 정도가 되었
다[8, 14]. 그러나 이 방법으로는 아직 염기의 손실 및 손상 정도를 예측하기는 어려운 것이 현실이다. Hada와 Sutherland 는 순수 분리한 T7 DNA(40-kb)를 여러 종류의 방사선으로 조사한 뒤 DNA 손상에 특이적으로 반응하는 효소를 이용 하여 DNA의 절단 및 염기의 손실 정도를 조사한 결과 LET 가 증가할수록 손상 정도 역시 증가하였으며, 엑스선과 같 은 전자기파 방사선에 비해 입자 방사선의 경우 염기의 탈 락이나 손상보다 DBS의 비율이 더 높음을 보고하였다[11].
DBS의 경우는 복구되기가 대단히 어려우므로, 세포의 사멸 이나 돌연변이 발생으로 이어지는 비율이 더 높은 것으로 알 려지고 있다.
방사선이 유발하는 돌연변이의 다양성 방사선을 통해 DNA가 손상을 입으면 정도가 심한 경우 에는 사멸을 초래하지만 세포내의 복구 기작에 따라 원상태 로 회복되거나 영구적인 돌연변이로 남게 된다. 균주 개량 이나 육종의 측면에서는 방사선의 종류나 선량에 따라 어떤 종류의 염기서열 변이 혹은 염색체 수준의 이상이 빈발하는 지 관심을 가질 수밖에 없다. 가장 간단한 assay 방법으로는 표현형의 관찰이 용이한 유전자를 포함하고 있는 플라스미 드에 방사선을 직접 조사한 뒤 대장균에 형질전환한 다음, 돌연변이체를 선별하여 해당 유전자 부위의 염기서열을 확 인하는 것이다. 그러나 DNA는 단백질과 결합한 상태로 세 포 내에 존재하고 있으며 염색체와 플라스미드의 돌연변이 발생 기작이 다른 것으로 알려져 있으므로[3, 29], 실제로 살 아있는 세포 내의 염색체에 위치한 유전자를 대상으로 연구 를 하는 것이 더 정확한 결과를 보장할 것이다.
돌연변이원 처리에 의해 세균에 발생하는 염기서열 변이 의 다양성을 측정하기 위한 유전자로서 대표적인 것으로는 tonB[18], lacI[32], lacZ[13], crp[38], rpoB[45] 등이 있다.
그러나 이와 같이 단일 유전자만을 이용한 검사 방법으로는 염색체 수준의 결실/삽입이나 rearrangement를 확인하기는 어려운 단점이 있다. Yu는 대장균에 저에너지N+ 이온을 주 입하여 rifampicin에 내성이 발생한 돌연변이체를 얻은 다음 rpoB 유전자를 PCR로 증폭하여 염기서열을 해독한 결과 감 마선을 이용한 돌연변이와 비교했을 시 밀집도가 높게 나타 나며 여러 위치에 염기열 변이가 발생하는 것으로 보고하였 다[45]. Tanaka 등은 고등 식물인 애기장대를 대상으로 한 이온빔 조사 과정에서 만들어진 tt 및 tl 돌연변이체의 염기 서열 분석 내용을 종합한 결과 대조군으로 사용한 전자빔과 비교했을 때 large DNA alteration의 비율이 월등히 높음을 보고하였으며[39], 대부분의 경우 DSB 혹은 clustered damage를 복구하는 과정에서 나타나는 non-homologous end joining(NHEJ)이 관련되어 있는 것으로 보았다[12, 34].
이온빔을 이용한 돌연변이 유발 방법의 실제 이온빔을 이용한 미생물의 돌연변이 유발 방법은 자외선 이나 감마선 조사를 통한 일반적인 방법과 크게 다르지 않 다. 그러나 입자 가속기는 일반 연구실 수준에서 갖출 수 있 는 장비가 아니므로, 외부 연구자에 개방되어 있는 주변의 가속기 시설에 사용 신청을 하여 빔 조사 실험을 진행해야 한다(Table 1). 국내에서 이용 가능한 연구용 가속기는 대부 분 양성자 사업단을 통해서 사용 신청을 할 수 있으며, MC- 50 싸이클로트론의 경우 한국원자력의학원을 통해 직접 이 용 신청을 할 수도 있다.
생물체에 방사선을 조사하면 일반적으로 생존율은 지수적 으로 감소하게 된다[47]. 생물종과 방사선(또는 방사되는 이 온)의 종류에 따라 생존율은 매우 다르므로, 본격적인 돌연 변이 유발 전에 선량에 따른 생존율 곡선(dose-effect curve) 을 실험적으로 구해야 한다. 종자나 포자와 같이 건조한 상 태의 생물학적 시료는 방사선에 대한 생존율이 일반적인 조 직이나 세포보다는 훨씬 높다. 또한 동일 종이라 하더라도 유전적 배경에 따라 생존율에 큰 차이가 있음에 유의해야 한 다. 생존율 곡선을 작성한 다음 생존세포 대비 돌연변이의 발생 빈도가 가장 높은 것으로 알려진 생존율 30~50% 정도 의 선량을 흡수하도록 조사한 뒤 적절한 방법에 의해 원하 는 돌연변이체를 선별해 나간다. 실제 적용에서 대부분의 이 온빔은 수평 방향으로 진행하도록 빔 라인이 구성되어 있으 므로, 액체 배지 상태로 담겨있는 미생물 시료의 경우 배양 용기를 적절히 위치시킬 방법을 마련해야 한다. 그리고 빔 의 에너지에 따라 투과할 수 있는 시료의 깊이가 다름을 감 안해야 한다. 앞서 설명했다시피 이온빔은 시료에 침투하면 서 일정 거리만큼 진행한 뒤 최대의 에너지를 내면서 멈추 Fig. 2. Bragg peak of a proton beam in water (solid line). While
X-rays used in cancer therapy (dotted line) deliver maximal energy upon entrance into a material, protons and other ion rays release most of their energy (about 80%) immediately before the particles come to rest. For uniform irradiation of an entire tumor, a Bragg peak can be modified into a SOPB (Spread Out Bragg Peak) using a passive beam modulator (dashed line). The peak can be further spread to deliver maximal energy from the point of entry.
게 되므로, 이러한 Bragg Peak가 시료를 완전히 투과한 이 후에 나타난다면 효율적인 조사가 되지 못한다. 따라서 빔 출구와 시료 사이에 특수한 장치를 설치하여 spread-out Bragg peak(SOBP)를 만드는 것이 중요하다(Fig. 1). 한국원 자력의학원의 MC-50 사이클로트론의 경우 양성자 사업단에 서 개발한 전용 빔라인에서 SOBP 발생 장치를 이용하면45 MeV의 양성자 빔을 물에 조사했을 때 입사지점부터 12 mm 의 깊이까지 고르게 선량이 부여되도록 조정되어 있다. 조 사 면적도 이온빔 실험에서 고려해야 할 요소이다. 원리적 으로 가속기로부터 인출되는 빔은 Gaussian 분포를 가지게 되어 정 중앙에서 가장 강도가 강하고, 주변부로 수직 거리 가 멀어질수록 점차 약해지는 분포를 보인다. 따라서 중앙 부 대비 90% 이내의 강도를 나타내는 위치를 조사 면적으 로 한정하여 시료를 위치시켜야 한다.
미생물 시료의 전처리 과정도 매우 중요하다. 보통 대수 기의 건강한 세포를 수확하여 PBS나 증류수로 배지 성분을 씻어 낸 다음 현탁액 상태로 시험관에 담아서 빔을 조사하 게 되는데, 빔 조사 전후의 시간을 최소화하여 생존력을 최 대화하는 것이 필요하다. 빔 에너지가 낮은 경우에는 투과 할 수 있는 생물학적 시료의 깊이에 한계가 있으므로, 시험 관에 세포 현탁액을 담는 대신 대수기의 세포를 세척하여 106~108 cells/mL 정도로 조정한 뒤 100 µL 정도를 petri dish에 점적하여 말린 뒤 빔을 조사하기도 한다. 뒤에 소개 할 이온 주입장치에서는 진공 분위기가 필요하므로, 이런 방 법으로 건조시킨 세포를 써야만 한다.
미생물 시료를 현탁액 상태로 동결하여 가속기 설비까지 이송한 다음 녹여서 빔을 조사하는 것도 가능한데, 이때 첨 가하는 글리세롤이나 DMSO 등의 동결보존제가 이온빔의 작용에 영향을 미치는지의 여부를 고려해야 하며, 동결 및 융해 과정에 의한 생존율 저해 작용도 감안해야 한다.
DMSO는 방사선 조사에서 유발되는 자유 라디칼을
quenching하는 효과가 있어서 세포[35] 또는 식물 종자[19]
에 처리하거나 동물에 투여했을 때[22] 생존율을 높인다는 것이 오래 전부터 알려져 있지만, 이 물질이 세포 내에서 일 어나는 DNA 절단 및 돌연변이 유발 미치는 영향에 대해서 는 산소의 존재 여부에 따라 영향이 다르게 나타난다[31].
스트레스에 대한 보호제로 쓰이는 glycine betaine 역시 방 사선에 대한 보호 작용이 있는 것으로 보고되고 있다[21].
저에너지 이온 주입장치를 사용할 때에는 진공을 유지해야 하므로, 생체 시료가 진공 조건에서 얼마나 생존을 유지할 수 있는지의 여부와 시료가 수분을 많이 함유한 경우 진공 을 걸어줄 때 냉각이 일어나면서 손상을 입지 않는지를 미 리 확인해야 할 것이다.
이온빔을 이용한 생명자원 개량 현황 방사선을 이용한 육종은 대부분 경제적으로 중요한 작물 에 치중하고 있으며, 아직까지는 감마선을 이용한 전통적인 방사선 돌연변이 육종이 강세를 보이고 있는 실정이다. 그 러나 최근 20년 동안 대규모의 이온 가속 장치가 건립되면 서 다양한 형태의 이온빔을 이용한 식물 품종 개량 성과가 보고되고 있다. 일본에서는 1991년 Takasaki Ion Accelera- tors for Advanced Radiation Application(TIARA)가 설립 되면서 이온빔에 의한 작물의 품종개량 및 생물학적 영향에 관한 연구를 시작한 것을 필두로 하여 현재는 이화학연구소 (RIKEN) 등 여러 곳에 이온빔 조사장치를 마련하여 융복합 연구를 진행하고 있다[39]. 대표적인 연구 성과로는 질병 저 항성 쌀과 UV-B 내성 애기장대[40], 화훼 작물의 개량[42]
등을 예로 들 수 있다. 중국의 경우에는 중국 과학원 플라스 마 연구소의 Yu를 중심으로 20여년 전부터 이온 주입기에 서 발생되는 저에너지 이온빔을 사용한 유전자원 개량 연구 를 독자적으로 진행해 오다 최근 들어 그 성과가 널리 알려 Table 1. Domestic ion beam accelerators for research purposes
Type Location Energy Ion type
45 MeV proton beam irradiation test facility KIRAMS1 10~38 MeV Proton
30 MeV high current cyclotron ARTI2 15~30 MeV Proton
20 MeV proton linear accelerator PEFP3 ~20 MeV Proton (pulses)
1 MV tandem accelerator PEFP3 0.3~1 MeV Proton
1.7 MV tandem accelerator KIGAM4 0.3~8 MeV H, D, B, C, O, F, Si, P, Cl, Fe
Gaseous ion implanter PEFP3 20~150 keV H, D, He, N, Ar, Xe, Kr
Metallic ion implanter PEFP3 20~120 keV Fe, Cu, Co, Gd
Ion implanter for semiconductor PEFP3 20~100 keV B, BF2, As, P, H, N, Ar
300 KeV ion beam irradiator ARTI2 ~300 keV H, He, N, Ar, Kr, Xe, Ne
Dual ion beam irradiator PEFP3 0.5~30 keV He, Ar, Xe
1Korea Institute of Radiological and Medical Sciences, Seoul.
2Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute, Jungeup-si, Jeollabuk-do.
3Proton Engineering Frontier Project, Korea Atomic Energy Research Institute, Daejeon.
4Korea Institute of Geology, Mining and Materials, Daejeon.
지기 시작하였다[45]. 1994년 이래 이온빔을 이용하여 21종 이상의 신품종을 확보하였으며, 아미노산과 유기산, 항생제, 효소 및 비타민 생산 능력이 증대된 수십 종의 개량 미생물 균주를 개발하였으며[10], 이외에도 이온빔을 이용한 식물 세포로의 성공적인 유전자 전달[46]을 보고하였다.
국내의 이온빔 장치 및 생명공학 분야의 활용 현황
국내에서는 1970년대 후반부터 치료 및 연구 목적의 입 자 가속기가 도입되기 시작하였으며, 반도체 제조 공정의 불 순물 도핑(doping) 공정에 쓰이는 100 KeV 급 이온 주입기 는 생산 라인에 이미 100대 이상 보급되어 있다. 그러나 생 명공학 분야, 특히 균주 개량 및 육종 분야에 이온빔이 활용 되기 시작한 것은 스웨덴 Scanditronix사에서 도입되어 1986 년부터 한국원자력의학원에서 가동되기 시작한 50 MeV 급 사이클로트론인 MC-50이 효시라 할 수 있다. MC-50은 양 성자 및 중양자, 헬륨을 가속하여 Be 표적에 조사할 때 발 생하는 고속 중성자를 이용한 치료를 목적으로 도입되었으 며, 동위원소 생산 목적으로 운용되어 오다가 현재는 국가 가속기 인프라로서 널리 활용되고 있다. 최근 5년간 평균 가 동률은 95.2%에 달하며, 2009년 한 해에만 31개 기관 51개 연구과제를 통해 327건의 이용 건수를 기록하였다. MC-50 이외에도 한국원자력연구원의 양성자기반공학기술개발사업 단과 정읍방사선과학연구소, 한국지질자원연구소 등에 다수 의 입자 가속기 및 이온 주입기가 설치되어 있다.
국내의 방사선 돌연변이 육종 연구는 주로 감마선을 이용 한 식량 작물과 화훼, 원예 및 산업소재용 식물 개량에 집중 되어 있다. 최근의 대표적인 연구 성과로는 한국원자력연구 원 정읍방사선과학연구소에서 감마선을 조사하여 아미노산 의 총 함량을 76% 높인 신품종 벼 골드아미 1호(GoldAmi- 1)를 개발한 것을 예로 들 수 있다. 한국원자력연구원은 1990 년대 이후 고품질·고기능성 특수미 개발에 주력하여 10개 품종의 품종보호권을 획득하고 국가품종목록에 등재한 바 있다.
이온빔을 이용한 국내의 생명자원 개량 연구는 MC-50 사 이클로트론의 활용이 본격화되고 2002년 출범한 양성자사업 단의 가속기 개발 및 빔 이용 연구 과제가 시작되면서 본 궤 도에 오르기 시작하였다. 양성자 사업단에서는 세부과제의 형태로서 ‘양성자 이온빔을 이용한 환경친화적 유용 유전자 원 개발’, ‘양성자빔을 이용한 고부가가치 화훼류 및 채소류 신품종 개발과 변이체 해석’, ‘저에너지 양성자 세라피 시스 템 개발’, ‘고에너지 대전류 양성자 빔을 이용한 방사성 동 위원소 개발’ 및 ‘양성자빔을 이용한 돌연변이 연구 허브 구 축’ 과제를 지원해 왔으며, 2002년부터 2010년까지는 대용 량 선형 가속기의 활용, 미래 이용자 육성 및 우수 연구분야 발굴을 위한 ‘양성자가속기 이용자 프로그램 개발 및 운영’
과제를 통해 이용자 프로그램을 운영하면서 약 50여개의 소 과제를 선정하여 지원하였다. 여기에는 나노공학, 생명공학, 정보통신, 우주항공, 핵물리, 의(공)학, 기초과학 등 다양한 분야가 망라되어 있는데, 생명공학 분야로는 조경수 및 유 채의 돌연변이 육종, 암 세포 사멸의 분자생물학적 기작 연
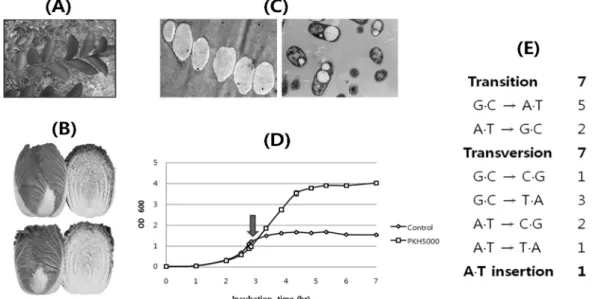
Fig. 3. Examples of strain improvement using proton beam-induced mutagenesis: (A) A Crape myrtle (Lagerstroemia indica) vari- ety with a characteristic flame-like reddish circumference around the leaf; (B) two varieties of Chinese cabbage, (upper, with improved flavor; lower, with disease resistance); (C) Escherichia coli strain producing a large amount of polyhydroxybutyrate (left), which eventually leads to autolysis (right, wild type control); (D) growth profile of 1-butanol-resistant Escherichia coli strain PKH5000 upon the addition of 0.9% (v/v) butanol (marked by the arrow); and (E) mutational pattern identified using high-cover- age sequencing of PKH5000.
구, 혈관성질환 치료에 대한 효용성 연구, 금속 나노수용체 의 표재성 종양모델에서 PIXE효과 연구, 질환모델 동물 개 발, 바이오매스 전처리와 당화효소 생산 균주 개발 과제 등 이 포함되어 있다. 양성자사업단의 지원을 통한 생명공학 분 야의 가장 대표적인 성과로는 대장균을 이용한 생분해성 플 라스틱 생산 균주의 개발이다. 한국원자력연구원에서는 대 장균에 양성자빔을 조사하여 유기산 생성이 억제된 돌연변 이 균주를 얻은 다음, 외래 유전자를 도입하여 생분해성 플 라스틱인 polyhyrdoxybutyrate(PHB)를 체내에 99.5% 이상 축적함과 동시에 autolysis가 일어나게 함으로써 중간 분리 공정을 제거하고 생산 단가를 현저하게 낮출 수 있음을 보 고하였다[17]. 또한 양성자 빔을 배추 종자에 조사함으로써 품종 개량 시간을 전통 육종방식보다 40% 가량 줄일 수 있 었으며, 이렇게 만들어진 돌연변이 유전자원 70종을 이용하 여 신품종 배추를 개발하여 기업에 이전하였는데 이는 양성 자빔을 이용한 국내 최초의 농작물 품종 개량 성과이기도 하 다[15]. 한편 필자는 2008년부터 양성자 빔 조사와 장기간 의 계대 배양을 통한 adaptive evolution을 병용하여 미생물 의 특성을 개량해 나가는 연구를 진행해 오고 있다. 원유의 부족과 온실가스 배출 문제로 인하여 재생 가능한 바이오매 스를 원재료로 하여 생물학적 방법으로 생산되는 바이오연 료에 대한 연구가 현재 매우 활발하게 이루어지고 있는데, 이 중에서 부탄올은 이미 실제 수송용 연료로 쓰이고 있는 에탄올에 비하여 여러 가지의 장점이 있다. 원래 부탄올의 생합성 경로는 Clostridium만이 갖고 있지만, 배양과 유전자 조작이 손쉬운 대장균에 이를 도입하여 부탄올을 생산하려 는 시도가 꾸준히 이어지고 있다. 그러나 부탄올은 유기용 매 중에서 가장 독성이 높은 부류에 속하므로 Clostridium이 나 일부 그람 양성 세균을 제외한 대부분의 미생물은 1%(V/
V) 이상의 농도에서 생육을 하지 못하는 문제가 있다[20].
본 연구진은 대장균에 양성자 빔을 조사한 뒤 최소 배지에 서 매일 계대 배양을 하면서 점차 부탄올의 농도를 높여 나 가고, 다시 빔 조사와 계대 배양을 반복해 나가면서 부탄올 에 대한 내성이 높아진 돌연변이체를 자연적으로 농화시켜 나가는 방법을 시도하였다[16]. 약 115일에 거쳐 9회의 빔 조사와 계대 배양을 거친 결과 약 1.4%의 부탄올에서 생존 가능한 돌연변이 집단을 확보하였으며, 유전체 염기서열 해 독을 통해 15 곳에서 염기서열 변이가 일어났음을 확인하였 다. 확인된 돌연변이(7 transitions, 7 transversions, 1 insertion)는 전부 최종 집단 내에서 83% 이상의 빈도를 차 지하고 있었다. Coding region에 발생한 11개의 돌연변이 중 에서 막을 구성하는 지방산의 특성을 변화시키는 유전자의 기능에 관한 연구를 현재 진행하고 있다. 이외에도 양성자 이온빔 조사를 통해 항생물질 생산능이 증대된 토양 세균과 방선균의 선발 작업이 진행 중이다.
결 론
입자가속기는 첨단 기술의 결정체로서 핵물리학이나 재료 공학, 의학의 분야를 넘어서 돌연변이 유발을 통한 생명 자 원의 개량에도 적용 가능한 국가 기반 연구 시설이 되고 있 다. 효율적인 돌연변이원이 갖추어야 할 특성으로는 돌연변 이 발생률이 높아야 하고, 다양한 형태의 돌연변이가 나타 나야 하며, 돌연변이 이외의 손상은 적게 유발해야 한다. 이 온빔은 이러한 측면에서 볼 때 전통적으로 널리 쓰여온 화 학적 돌연변이원 또는 기타 방사선에 비하여 우수한 것으로 알려지고 있으나 이온빔과 생명 자원의 종류에 따른 구체적 인 적용 방법에 대한 좀 더 체계적인 연구 결과가 필요하 며, 최근 각광을 받고 있는 차세대 염기서열 해독 기법(next generation sequencing)이 유전체 수준에 발생한 염기서열 변 이의 양상을 해석하는 데에 크게 기여할 것으로 생각된다.
특히 경주에 건설 중인 양성자가속기연구센터는 양성자빔을 이용한 첨단 융합 연구의 산실이 될 것이며, 이를 통한 생명 자원의 개량 연구도 더욱 가속화될 것이다.
감사의 글
본 논문을 쓰기 위한 자료와 사진을 제공해 주신 양성자 기반공학기술개발 사업단 사무국, 한국원자력연구원 윤상필 님, 그리고 한국원자력의학원 양태건 박사님과 황원택님에 게 감사를 드립니다. 본 연구는 과학기술부 21세기 프론티 어 미생물유전체활용기술개발사업 및 양성자기반공학기술개 발사업의 지원에 의해 이루어졌습니다.
REFERENCES
1. FAO/IAEA Mutant Variety and Genetic Stock Database.
http://mvgs.iaea.org/
2. Buckling, A., R. Craig Maclean, M. A. Brockhurst, and N.
Colegrave. 2009. The Beagle in a bottle. Nature 457: 824- 829.
3. Cairns, J., J. Overbaugh, and S. Miller. 1988. The origin of mutants. Nature 335: 142-145.
4. Datsenko, K. A. and B. L. Wanner. 2000. One-step inactiva- tion of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. U S A 97: 6640-6645.
5. Edwards, J. S., R. U. Ibarra, and B. O. Palsson. 2001. In silico predictions of Escherichia coli metabolic capabilities are consistent with experimental data. Nat. Biotechnol. 19:
125-130.
6. Ellis, H. M., D. Yu, T. DiTizio, and D. L. Court. 2001. High efficiency mutagenesis, repair, and engineering of chromo- somal DNA using single-stranded oligonucleotides. Proc.
Natl. Acad. Sci. U S A 98: 6742-6746.
7. Fong, S. S., A. P. Burgard, C. D. Herring, E. M. Knight, F.
R. Blattner, C. D. Maranas, and B. O. Palsson. 2005. In
silico design and adaptive evolution of Escherichia coli for production of lactic acid. Biotechnol. Bioeng. 91: 643-648.
8. Friedland, W., P. Jacob, P. Bernhardt, H. G. Paretzke, and M.
Dingfelder. 2003. Simulation of DNA damage after proton irradiation. Radiat. Res. 159: 401-410.
9. Goodhead, D. T. 1994. Initial events in the cellular effects of ionizing radiations: clustered damage in DNA. Int. J. Radiat.
Biol. 65: 7-17.
10. Gu, S. B., S. C. Li, H. Y. Feng, Y. Wu, and Z. L. Yu. 2008.
A novel approach to microbial breeding-low-energy ion im- plantation. Appl. Microbiol. Biotechnol. 78: 201-209.
11. Hada, M. and B. M. Sutherland. 2006. Spectrum of complex DNA damages depends on the incident radiation. Radiat.
Res. 165: 223-230.
12. Hada, M. and A. G. Georgakilas. 2008. Formation of cluster- ed DNA damage after high-LET irradiation: a review. J.
Radiat. Res. 49: 203-210.
13. Hoebee, B., J. Brouwer, P. van de Putte, H. Loman, and J.
Retel. 1988. 60Co gamma-rays induce predominantly C/G to G/C transversions in double-stranded M13 DNA. Nucleic Acids Res. 16: 8147-8156.
14. Hsiao, Y. and R. D. Stewart. 2008. Monte Carlo simulation of DNA damage induction by x-rays and selected radioiso- topes. Phys. Med. Biol. 53: 233-244.
15. h t t p : / / w w w. d t . c o . k r / c o n t e n t s . h t m l ? a r t i c l e _ n o = 2008110402019922732016.
16. Jeong, H. and J. Han. 2010. Enhancing the 1-butanol tolerance in Escherichia coli through repetitive proton beam irradiation. J. Kor. Phy. Soc. 56: 2041-2045.
17. Jung, I. L., K. H. Phyo, K. C. Kim, H. K. Park, and I. G. Kim.
2005. Spontaneous liberation of intracellular polyhydroxy- butyrate granules in Escherichia coli. Res. Microbiol. 156:
865-873.
18. Kanbashi, K., X. Wang, J. Komura, T. Ono, and K. Yamamoto.
1997. Frameshifts, base substitutions and minute deletions constitute X-ray-induced mutations in the endogenous tonB gene of Escherichia coli K12. Mutat. Res. 385: 259-267.
19. Kaul, B. L. 1970. Studies on radioprotective role of dimethyl sulfoxide in plants. Radiation Botany 10: 69-78.
20. Knoshaug, E. P. and M. Zhang. 2009. Butanol tolerance in a selection of microorganisms. Appl. Biochem. Biotechnol.
153: 13-20.
21. Monobe, M., A. Uzawa, M. Hino, K. Ando, and S. Kojima.
2005. Glycine betaine, a beer component, protects radiation- induced injury. J. Radiat. Res. 46: 117-121.
22. Moos, W. S. and S. E. Kim. 1966. Radioprotective effect of topically applied dimethyl sulfoxide on mice. Experientia 22: 814.
23. Muller, H. J. 1927. Artificial Transmutation of the Gene.
Science 66: 84-87.
24. Park, H. S., S. H. Nam, J. K. Lee, C. N. Yoon, B.
Mannervik, S. J. Benkovic, and H. S. Kim. 2006. Design and evolution of new catalytic activity with an existing protein scaffold. Science 311: 535-538.
25. Park, J. H., S. Y. Lee, T. Y. Kim, and H. U. Kim. 2008.
Application of systems biology for bioprocess development.
Trends Biotechnol. 26: 404-412.
26. Posfai, G., G. Plunkett, 3rd, T. Feher, D. Frisch, G. M. Keil, K. Umenhoffer, V. Kolisnychenko, B. Stahl, S. S. Sharma, M. de Arruda, V. Burland, S. W. Harcum, and F. R. Blattner.
2006. Emergent properties of reduced-genome Escherichia coli. Science 312: 1044-1046.
27. Reed, J. L. and B. O. Palsson. 2003. Thirteen years of build- ing constraint-based in silico models of Escherichia coli. J.
Bacteriol. 185: 2692-2699.
28. Romero, P. A. and F. H. Arnold. 2009. Exploring protein fitness landscapes by directed evolution. Nat. Rev. Mol. Cell Biol. 10: 866-876.
29. Rosenberg, S. M. 2001. Evolving responsively: adaptive mutation. Nat. Rev. Genet. 2: 504-515.
30. Rust, D. M. 1992. Solar flares: An overview. Adv. Space Res. 12: 289-301.
31. Sapora, O., F. Barone, M. Belli, A. Maggi, M. Quintiliani, and M. A. Tabocchini. 1991. Relationships between cell killing, mutation induction and DNA damage in X-irradiated V79 cells: the influence of oxygen and DMSO. Int. J.
Radiat. Biol. 60: 467-482.
32. Sargentini, N. J. and K. C. Smith. 1994. DNA sequence analysis of gamma-radiation (anoxic)-induced and spontane- ous lacId mutations in Escherichia coli K-12. Mutat. Res.
309: 147-163.
33. Sharan, S. K., L. C. Thomason, S. G. Kuznetsov, and D. L.
Court. 2009. Recombineering: a homologous recombination- based method of genetic engineering. Nat. Protoc. 4: 206- 223.
34. Shikazono, N., M. Noguchi, K. Fujii, A. Urushibara, and A.
Yokoya. 2009. The yield, processing, and biological conse- quences of clustered DNA damage induced by ionizing radiation. J. Radiat. Res. 50: 27-36.
35. Singh, D. R., J. M. Mahajan, and D. Krishnan. 1976. Effect of dimethyl sulfoxide (DMSO) on radiation-induced hetero- allelic reversion in diploid yeast. Mutat. Res. 37: 193-200.
36. Stadler, L. J. 1928. Mutations in Barley Induced by X-Rays and Radium. Science 68: 186-187.
37. Stemmer, W. P. 1994. Rapid evolution of a protein in vitro by DNA shuffling. Nature 370: 389-391.
38. Takimoto, K., K. Uchino, K. Ishizaki, and M. Ikenaga. 1991.
Specificity of mutational DNA sequence changes induced by X-rays in the cloned Escherichia coli crp gene. Mutat. Res.
254: 199-206.
39. Tanaka, A., N. Shikazono and Y. Hase. Studies on biological effects of ion beams on lethality, molecular nature of muta- tion, mutation rate, and spectrum of mutation phenotype for mutation breeding in higher plants. J. Radiat. Res. 51: 223- 233.
40. Tanaka, A., A. Sakamoto, Y. Ishigaki, O. Nikaido, G. Sun, Y.
Hase, N. Shikazono, S. Tano, and H. Watanabe. 2002. An ultraviolet-B-resistant mutant with enhanced DNA repair in Arabidopsis. Plant Physiol. 129: 64-71.
41. Wang, H. H., F. J. Isaacs, P. A. Carr, Z. Z. Sun, G. Xu, C. R.
Forest, and G. M. Church. 2009. Programming cells by multiplex genome engineering and accelerated evolution.
Nature 460: 894-898.
42. Yamaguchi, H., S. Nagatomi, T. Morishita, K. Degi, A.
Tanaka, N. Shikazono, and Y. Hase. 2003. Mutation induced with ion beam irradiation in rose. Nucl. Instrum. Meth. B 206: 561-564.
43. Yu, B. J., B. H. Sung, M. D. Koob, C. H. Lee, J. H. Lee, W.
S. Lee, M. S. Kim, and S. C. Kim. 2002. Minimization of the Escherichia coli genome using a Tn5-targeted Cre/loxP excision system. Nat. Biotechnol. 20: 1018-1023.
44. Yu, Z., J. Deng, J. He, Y. Huo, Y. Wu, X. Wang, and G. Lui.
1991. Mutation breeding by ion implantation. Nucl. Instrum.
Meth. B 59-60: 705-708.
45. Yu, Z. 2007. Study on the interaction of low-energy ions with organisms. Surf. Coat. Tech. 201: 8006-8013.
46. Zengliang, Y., Y. Jianbo, W. Yuejin, C. Beijiu, H. Jianjun, and H. Yuping. 1993. Transferring Gus gene into intact rice cells by low energy ion beam. Nucl. Instrum. Meth. B 80-81:
1328-1331.
47. Zengliang, Y. Introduction to Ion Beam Biotechnology, Springer Science+Buisiness Media Inc., New York, 2006.
(Received July 21, 2010/Accepted August 10, 2010)