면역세포의 Metabolic reprogramming 강수진 Page 1 / 8 BRIC View 2019-T04
면역세포의 Metabolic reprogramming
강 수 진
Osaka University E-mail: [email protected] 요약문 Metabolism은 세포기능의 근본이 되는 중요한 요소이다. 최근 면역세포의 Metabolism과 면역반응조절의 관계에 관한 연구가 보고되면서, 면역대사 -Immunometabolism- 라는 개념이 주목을 받고 있다. 염증질환 및 암의 발병에는 비정상적인 면역대사 활성화조절이 크게 관여하고 있음이 밝혀지면서, 신규의 면역학적 치료표적으로서 대사관련분자가 주목을 받고 있다. 본 리포트에서는 3대 영양소인 당, 아미노산, 지질의 대사와 면역반응의 관계를 소개하고자 한다.Key Words:Metabolism, 면역세포, 당, 아미노산, 지질
목 차
1. 서론 2. 본론 2.1 면역대사에 관여하는 주요한 대사경로 2.2 각 대사경로에 의한 면역반응조절 2.3 암과 아미노산 대사 3. 결론 4. 참고문헌면역세포의 Metabolic reprogramming 강수진 Page 2 / 8
1. 서론
감염에 대한 방어의 역할을 담당하는 면역세포가 과잉 에너지상태로 인한 활성화가 유도되어 염증반응을 일으키는 것이 알려져 있다. 이러한 자연면역계반응에는 마크로파아지가 중심적인 player로 알려져 있지만, 최근 여러 보고에 의하여 상상이상의 다양한 종류의 면역세포가 관여하며, 자연면역세포뿐만 아니라 획득면역계의 활성화도 동시에 발생한다는 점이 밝혀지게 되었다. 하지만, 이러한 면역세포가 형성하는 네트워크의 전반적인 메커니즘은 구체적으로 밝혀지지 않고 있다. 예를 들어, 만성염증질환인 “비만(Obesity)”의 초기단계에 활성화되는 세포는 어떠한 종류의 면역세포이며, trigger는 무엇이며, 그리고 어떠한 cascade로 “지방조직의 만성염증”이라는 상태로 이어지는가에 대해 충분히 밝혀지지 않고 있다. 또한 최근에는, 마크로파아지를 중심으로 세포내대사를 변환시키는 인자(mTOR pathway, HIF-1α pathway, 지방산합성을 조절하는 pathway 등)이 염증반응의 조절에 관여하고 있다는 점이 밝혀지게 되었다. 특히, 면역세포의 세포내대사상태가 면역세포의 활성화를 조절하여 “대사와 면역세포기능조절”에 관하여 다층적·다차원적인 이해와 해석이 필요하다고 생각된다. 이번 보고서에서는 현재까지 알려진 면역대사에 관여하는 주요한 대사경로에 대하여 대략적으로 정리하고, 각 대사상태가 어떻게 면역세포의 기능을 조절하는지에 대하여 서술하고자 한다.2. 본론
2.1 면역대사에 관여하는 주요한 대사경로
세포 내 대사경로는 복잡하게 얽혀져 있다. 면역대사에 관여하는 주요한 대사경로에 대하여 자세히 서술하고자 한다. 1) 해당계(Glycolysis) 세포질에는 glucose를 분해하는 대사경로가 존재하며, pyruvate가 최종산물로서 생산된다. 이후에 서술되는 citric acid cycle과 비교하여 ATP 생산에 대하여 비교적 비효율적이지만, glucose 한 분자에 대하여 2분자의 ATP가 생산된다. 하지만, 해당계는, 보조효소로서 중요한 NAD+를 NADH으로 환원시키는 동시에, 중간산물이 핵산, 아미노산, 지질산합성의 기질이 되는 등 에너지 생산 이외에도 중요한 역할을 담당하고 있다. 해당계를 유지하기 위하여, 세포는 pyruvate를 lactic acid으로 환원시키면서 NADH를 산화하여, NAD+를 일정량 유지시키는 데에 관여한다. PI3K 경로 및 MAPK 경로 등의 세포증식에 관여하는 시그널에 관여하는 대부분의 경로가 해당계의 활성화를 촉진시키는 것이 알려져 있다. 따라서, 해당계는 세포증식에 중요한 대사경로라고 생각된다.면역세포의 Metabolic reprogramming 강수진 Page 3 / 8 2) TCA 회로(Citric acid cycle)
Mitochondrial matrix에 발생하는 대사경로로서, glucose, 지질, 아미노산을 기질로서 이용한다. TCA 회로는 NADH와 FADH2를 생산하고, 이러한 효소를 통하여 미토콘드리아 내막의 전자전달계로 전자를 공여함으로써 oxidative phosphorylation을 촉진하며, 상당히 효율적인 에너지생산에 관여하는 대사경로이다.
3) Pentose phosphate pathway
세포질에는 해당계의 중간산물을 기질로서 사용하여 핵산 및 아미노산 전구체를 생산하는 대사경로이며, 세포증식을 촉진한다. NADPH의 생산원으로도 중요한 경로로 알려져 있으며, 생산된 NADPH는 지질산합성에 이용된다.
4) 아미노산대사
세포 내에 존재하는 아미노산은 주로 단백질 및 branched chain fatty acid 의 기질로서 중요한 세포내 영양소로서 알려져 있다. 아미노산 대사의 주요한 역할은 mTOR (mechanistic target of rapamycin) 시그널 경로이며, lysosome에 존재하는 mTOR complex가 각종 아미노산의 양을 감지하여, 하류시그널의 신호를 전달하는 기능을 담당하고 있다. Glutamine, aspartic acid는 de novo 핵산합성에 이용된다. 세포증식과정에서는 glutamine이 TCA 회로에 사용되며, 에너지생산 및 지방산합성에 관여한다.
5) 지방산산화
미토콘드리아에서 지방산으로부터 에너지를 생산하는 경로를 담당한다. 세포 내에 존재하는 지방산은 미토콘드리아의 외막에서 fatty acid acyl CoA으로 활성화된다. Mitochondrial matrix에 유송된 fatty acid acyl CoA는 지방산 산화 과정을 거쳐 acetyl CoA, NADH, FADH2을 생산한다. 이러한 산물은TCA회로 및 전자전달계에서 ATP생산에 이용된다.
6) 지방산합성
세포증식 시에는 세포구성성분으로서 지질의 수요가 높아지게 된다. 지방산합성은 주로 mTOR의 하류시그널로서 유도되는 SREBP (sterol regulatory element binding protein), FASN (fatty acid synthase), ACC (acetyl CoA carboxylase) 등의 효소에 의해 de novo 지질합성에 관여한다. 해당계, TCA 회로, pentose phosphate pathway의 중간산물이 지방산합성의 기질로서 사용된다.
면역세포의 Metabolic reprogramming 강수진 Page 4 / 8
2.2 각 대사경로에 의한 면역반응조절
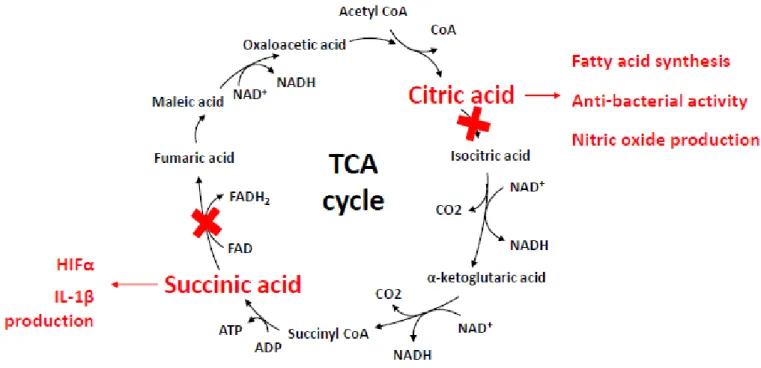
면역반응에는 상기에 서술된 대사경로에 대한 의존도의 변화가 발생한다 (그림 1). 이러한 변화는 적절한 면역반응에 필수 요소로서 작용하는 것이 최근 보고되었다. 이하의 각 대사경로의 면역반응의 조절작용에 대하여 서술하고자 한다. 그림 1. 면역세포 분화에 필요한 대사경로. 1) 해당계 염증성 마크로파아지(M1) 및 수지상세포, 활성화 NK 세포, 활성화 effector T 세포, 활성화 B 세포에는 해당계의 활성화(aerobic glycolysis)가 유도된다 [1, 2]. 해당계의 활성화는 패턴인식수용체, 사이토카인수용체, 항원수용체 등의 외부 자극에 대하여 급속히 면역세포 활성화를 유도한다. 해당계의 활성화에 의해 면역세포는 effector 기능을 획득하기 위한 에너지와 기질을 얻게 된다. 마크로파아지의 탐식작용(phagocytosis)이나 염증성 사이토카인의 생산, 수지상세포의 항원 제시, Th17 세포의 IL-17 (Interleukin-17) 생산 등에 해당계의 활성화가 필수적 요소로서 작용되는 것이 알려져 있다 [3]. 2) TCA 회로항염증성 마크로파아지(M2) 및 memory T 세포에는 oxidative phosphorylation의 활성화가 유도된다. 반면, 염증성 마크로파아지(M1)는 IDH (isocitrate dehydrogenase)의 발현이 현저히 감소되며, itaconic acid에 의한 SDH (succinate hydrogenase)의 저해를 유도하여, TCA 회로 산물인
면역세포의 Metabolic reprogramming 강수진 Page 5 / 8 citric acid 및 succinic acid의 직후에 TCA 회로의 분절이 발생하게 된다 (그림 2). TCA 회로의 분절에 의해 citric acid 및 succinic acid이 세포 내에 축적되면서, citric acid에 의해 지방산합성 및 itaconic acid, NO (nitric acid) 생산이 증진되며, succinic acid에 의해 HIFα (hypoxia inducible factor 1α)의 활성화 및 IL-1β (Interleukin-1β)의 생산에 이용된다 [4, 5].
그림 2. 염증성 면역세포의 TCA 회로의 분절현상. 3) 아미노산대사
3-1) Glutamine
Glutamine대사는 패혈증(Sepsis) 및 열상(Burns)의 발병시 유도되는 면역반응에 중요한 요소로서 알려져 있다. 최근에는 마크로파아지 분화에도 필수적으로 작용하는 것이 보고되었다. 염증성 마크로파아지(M1)의 분화에는 glutamine이 관여하지 않지만, 항염증성 마크로파아지(M2)의 분화에는 TCA 회로 및 Hexosamine 경로(Hexosamine biosynthesis pathway, HBP)로 glutamine의 유입이 필수적으로 작용된다는 것이 알려져 있다 [6]. Glutamine 대사는 림프구의 활성을 제어에도 관여한다. 특히, 항원수용체의 자극을 받은 림프구는 glutamine의 수요가 급격히 증가하게 된다. Glutamine 및 Leucine 등의 중성 아미노산의 uptake에 관여하는 transporter로서 ASCT2가 알려져 있다. ASCT2 결손 및 glutamine 결핍은 mTOR complex 1 (mTORC1)의 활성 저하를 유도하여, effector T 세포의 기능을 저하시킨다 [7].
3-2) Arginine
면역세포의 Metabolic reprogramming 강수진 Page 6 / 8 Arginine을 일산화산소합성 경로와 arginase 경로의 두가지 경로에서 이용한다. 일산화산소합성 경로에 필요한 arginine 유입은 염증성 마크로파아지의 기능을 활성화시킨다. 반면, arginase 경로에 의한 arginine 대사는 항염증성 마크로파아지의 기능을 활성화에 필수요소로 작용된다 [8]. 4) 지방산산화 Aerobic glycolysis가 염증성 면역세포 및 증식기세포에서 활성화되는 점에 비해, 지방산산화는 항염증성 마크로파아지 및 억제성 T 세포(Regulatory T, Treg cell), memory T 세포 등의 대사과정의 중추적 역할을 담당하고 있다. 최근 지방산산화과정이 마크로파아지의 분화에 필수적으로 작용하는 것이 보고 되었다. IFN-γ (Interferon γ) 및 LPS (Lipopolysaccharide)에 의해 유도되는 염증성 마크로파아지가 aerobic glycolysis에 의존적으로 분화되며, 반면, IL-4 (Interleukin-4) 에 의해 유도되는 항염증성 마크로파아지는 STAT6 및 PGC1β를 통한 지방산산화의 활성화에 의존적으로 분화가 유도되는 것이 알려져 있다 [9, 10]. T cell subset 중에서도 지방산산화에 대한 의존도가 각각의 T 세포에서 크게 다르다는 점이 주목을 받고 있다. Effector T 세포는 활성화 과정에서 지방산산화의 활성화저해가 요구되는 반면, 억제성 T 세포의 분화과정에서는 지방산산화의 활성화가 필수적으로 작용하는 것이 보고되었다 [2]. Memory T 세포의 분화 및 maintenance에도 지방산산화의 활성화가 요구된다 [11]. 5) 지방산합성 지방산산화와 대조적으로, 지방산합성은 염증성반응에 중요한 매개체로서 작용한다. LPS 및 사이토카인에 대한 자극을 받은 마크로파아지는 지방산합성과정을 활성화하여, 염증성 마크로파아지 분화를 촉진시킨다 [12]. 지방산합성은 수지상세포의 기능제어에도 중요한 역할을 한다. 수지상세포의 활성화 및 CD8+ T 세포에 항원제시과정에 지방산합성의 활성화가 필수요소로서 작용하는 점이 보고되었다 [13]. 최근, 림프구의 세포증식과정에 지방산합성과정이 요구되는 것이 보고되었다. 지방산합성은 Th17 세포의 분화에 필수요소이지만, 억제성 T 세포의 기능에는 불필요 요소인 점이 보고되었다 [14]. 따라서, 지방산산화와 같이 지방산합성도 Effector T 세포와 억제성 T 세포의 balance 제어에 중요한 역할을 담당하는 요소임을 시사한다.
2.3 암과 아미노산 대사
암세포는 대사과정의 reprogramming에 의해 세포의 생존, 증식, 전이 등을 조절한다. 이를 위하여, 충분한 에너지의 공급과 세포 증식에 필요한 단백질, 지질, 핵산 등의 고분자화합물의 공급이 중요하다. 이러한 대사의 reprogramming에 의해 암세포는 왕성한 세포 증식을 거쳐 세포사(apoptosis)를 억제한다.면역세포의 Metabolic reprogramming 강수진 Page 7 / 8 1) Glutamine
소장상피세포와 같이 증식이 빠른 세포에는 glutamine이 중요한 에너지원으로서 고분자화합물의 재료로서 사용된다. Glutamine 이 α-ketoglutamate (α-KG)를 통하여 lactic acid으로 분해되는 것을 “Glutaminolysis”라고 하며, 암세포는 glutaminolysis를 통하여 NAPDH 공급에 중요한 역할을 한다. 또한, glutamine은 TCA 회로 중간체를 공급하는 것에 의해 세포증식에 필요한 고분자화합물과 환원력을 제공하게 된다. 통상적으로, TCA 회로에 있어서 α-KG를 생산에 관여하는 효소는 IDH이지만, 이 유전자에는 빈번히 변이가 발생하는 것이 보고 된 바 있다. 변이가 발생한 IDH는 α-KG를 2-hydroxyglutarate (2-HG)으로 전환을 유도하여, 이 2-HG는 암세포에 의해 크게 변동하는 대사물 “oncometabolite”로 알려져 있다. 2-HG는α-KG와 산소를 기질로서 이용하여 대부분의 효소반응을 저해함으로써 암세포의 증식에 관여하게 된다. 2) Tryptophan 암세포는 정상세포와 비교하여 세포의 증식, 생존, 전이과정에 세포간 커뮤니케이션을 필요로 한다. 암세포는 면역계를 회피하기 위한 생존과정으로서, 암세포 내의 tryptophan의 대사산물인 kynurenine을 autocrine 또는 paracrine 형식으로 분비하여, 생존과 증식을 가능하게 하는 전략을 가지고 있다. 또한, tryptophan의 대사과정에 관여하는 indoleamine 2,3-dioxygenase을 저해하여 동물 모델의 암형성을 억제하게 되는 것이 보고되었다. 하지만, 분자 메커니즘에 대해서는 밝혀지지 않지만, 항암신약개발로 진행되고 있다.
3) 그 외의 아미노산
암세포에 있어 산화환원전위의 유지는, 산화스트레스에 대한 저항성을 증강하여 세포사로부터 회피하여 생존을 확보하기 위한 중요한 과정이다. 세포내의 주요한 환원제는 NAPDH와 glutathione이 있으며, NAPDH는 Warburg 효과에 의해 Pentose phosphate pathway 및 malic enzyme에 의해 공급된다. 한편, glutathione의 유지는 주로 cysteine의 공급에 의존한다. 따라서, 암세포의 생존, 악성화에 있어 sulfur-containing amino acid의 공급은 필수요소가 된다.
3. 결론
염증반응에는 활성화된 여러 면역세포의 네트워크형성에 의해 이루어진다. 이러한 면역세포의 활성화를 조절하는 여러 인자들이 알려져 있지만, 최근 여러 보고에 의해 각 면역세포의 세포 내 대사활성상태가 면역세포의 기능을 조절하는 것이 알려지게 되었다. 장래적으로 이러한 면역세포의 대사활성제어를 표적으로 한 영양소 및 약제가 개발된다면 신규의 영양요법 및 치료법으로 발전될 수 있는 가능성이 시사된다.면역세포의 Metabolic reprogramming 강수진 Page 8 / 8
4. 참고문헌
[1] Krawczyk CM and Pearce EJ, et al: Toll-like receptor-induced changes in glycolytic metabolism regulate dendritic cell activation. Blood, 115: 4742-4749, 2010.
[2] Michalek RD and Rathmell JC, et al: Cutting edge: distinct glycolytic and lipid oxidative metabolic programs are essential for effector and regulatory CD4+ T cell subsets. J Immunol, 186: 3299-3303, 2011.
[3] Shi LZ and Chi H, et al: HIF1α-dependent glycolytic pathway orchestrates a metabolic checkpoint for the differentiation of TH17 and Treg cells. J Exp Med. 208: 1367-1376, 2011.
[4] Michelucci A and Hiller K, et al: Immune-responsive gene 1 protein links metabolism to immunity by catalyzing itaconic acid production. Proc Natl Acad Sci U S A, 110: 7820-7825, 2013.
[5] Tannahill GM and O’Neill LA, et al: Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature, 496: 238-242, 2013.
[6] Jha AK and Artyomov MN, et al; Network integration of parallel metabolic and transcriptional data reveals metabolic modules that regulate macrophage polarization. Immunity, 42: 419-430, 2015.
[7] Nakaya M and Sun SC, et al: Inflammatory T cell responses rely on amino acid transporter ASCT2 facilitation of glutamine uptake and mTORC1 kinase activation. Immunity, 40: 692-705, 2014.
[8] Rath M and Munder M, et al: Metabolism via Arginase or Nitric Oxide Synthase: Two Competing Arginine Pathways in Macrophages. Front Immunol, 5: 532, 2014.
[9] Vats D and Chawla A, et al: Oxidative metabolism and PGC-1beta attenuate macrophage-mediated inflammation. Cell Metab, 4: 13-24, 2006.
[10] Huang SC and Pearce EJ, et al: Cell-intrinsic lysosomal lipolysis is essential for alternative activation of macrophages. Nat Immunol, 15: 846-855, 2014.
[11] van der Windt GJ and Pearce EL, et al: Mitochondrial respiratory capacity is a critical regulator of CD8+ T cell memory development. Immunity, 36: 68-78, 2012.
[12] Posokhova EN and Dushkin MI, et al: Lipid synthesis in macrophages during inflammation in vivo: effect of agonists of peroxisome proliferator activated receptors alpha and gamma and of retinoid X receptors. Biochemistry (Mosc), 73: 296-304, 2008.
[13] Everts B and Pearce EJ, et al: TLR-driven early glycolytic reprogramming via the kinases TBK1-IKKɛ supports the anabolic demands of dendritic cell activation. Nat Immunol, 15: 323-332, 2014.
[14] Berod L and Sparwasser T, et al: De novo fatty acid synthesis controls the fate between regulatory T and T helper 17 cells. Nat Med, 20: 1327-1333, 2014.
The views and opinions expressed by its writers do not necessarily reflect those of the Biological Research Information Center.
강수진(2019). 면역세포의 Metabolic reprogramming. BRIC View 2019-T04
Available from http://www.ibric.org/myboard/read.php?Board=report&id=3162 (Feb 28, 2019) Email: [email protected]