Introduction
Introduction
Osteosarcoma is one of the most common primary ma-lignant bone tumors arising from mesenchymal stem cells, and occurs mainly in late childhood and early adolescence [1]. Chemotherapy treating patients with nonmetastatic osteo-sarcoma has dramatically improved 5-year survival rates from < 20 to 60–70% [2,3]. Understanding the underlying molecular mechanisms of osteosarcoma is the most important steps in the treatment process [4]. A new treatment strategy is highly desirable to increase the survival rate of osteosarcoma patients [4].
Most anticancer drugs act as chemotherapy agents for can-cer by causing cell apoptosis and inhibiting cancan-cer cell prolifer-ation [5-7]. Therefore, apoptosis of cancer cells due to the use of these anticancer drugs has become an important indicator of cancer treatment results [8,9]. Apoptosis can occur in can-cer through a death receptor-dependent extrinsic pathway or mitochondria-dependent intrinsic pathway that can be induced by chemotherapy treatment [10,11].
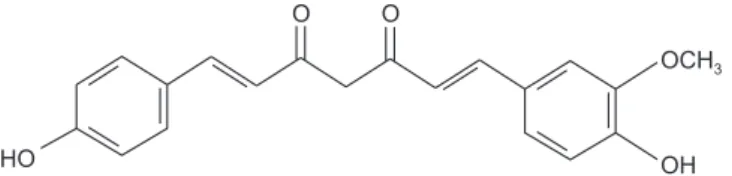
Demethoxycurcumin ((E,E)-1-(4-Hydroxy-3-methoxy-phenyl)-7-(4-hydroxyphenyl)-1,6-heptadiene-3,5-dione, DMC, Fig. 1) is a curcuminoid found in turmeric and is called curcumin II or desmethoxycurcumin [12]. Curcuminoids are
Int J Oral Biol 46:23-29, 2021 pISSN: 1226-7155 • eISSN: 2287-6618 https://doi.org/10.11620/IJOB.2021.46.1.23
Apoptotic activity of demethoxycurcumin in MG-63
human osteosarcoma cells
Kyeong-Rok Kang
1, Jae-Sung Kim
1, Tae-Hyeon Kim
1, Jeong-Yeon Seo
1,2, Jong-Hyun Park
1, Hong Sung Chun
2,
Sun-Kyoung Yu
1, Heung-Joong Kim
1, Chun Sung Kim
1, and Do Kyung Kim
1*
1The Institute of Dental Science, Chosun University, Gwangju 61452, Republic of Korea
2Department of Integrative Biological Sciences & BK21 FOUR Educational Research Group for Age-associated Disorder Control Technology, Chosun University, Gwangju 61452, Republic of Korea
Demethoxycurcumin (DMC), which is a curcuminoid found in turmeric, has anti-proliferative effects on cancer cells. However, the effect of DMC on osteosarcoma has not been established. The aim of this study was to examine the effects of DMC on cell growth and apoptosis induction in MG-63 human osteosarcoma cells. This study was investigated using 3-[4, 5-dimethylthiazol-2-yl]-2, 5 diphenyl tetrazolium bromid assay, Live/Dead cell assay, 4’, 6-diamidino-2-phenylindole staining, and immunoblotting in MG-63 cells. DMC induced MG-63 cell death in a dose-dependent manner, with an estimated IC50 value of 54.4 µM. DMC treatment resulted in nuclear condensation in
MG-63 cells. DMC-induced apoptosis in MG-63 cells was mediated by the expression of Fas and activation of caspase-8, caspase-3, and poly (ADP-ribose) polymerase. Immunoblotting results showed that Bcl-2 and Bcl-xL were downregulated, while Bax and Bad were upregulated by DMC in MG-63 cells. These results indicated that DMC inhibits cell proliferation and induces apoptotic cell death in MG-63 human osteosarcoma cells via the death receptor-mediated extrinsic apoptotic pathway and mitochondria-receptor-mediated intrinsic apoptotic pathway.
Keywords: Demethoxycurcumin, Cell death, Apoptosis, Anticancer effect
Received January 21, 2021; Revised March 4, 2021; Accepted March 4, 2021
*Correspondence to: Do Kyung Kim, E-mail: [email protected] https://orcid.org/0000-0001-6254-946X Copyright © The Korean Academy of Oral Biology
CC This is an open-access article distributed under the terms of the Creative Commons Attribution Non-Commercial License
(http://creativecommons.org/licenses/by-nc/4.0/), which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
linear diarylheptanoids with molecules such as curcumin, or curcumin derivatives with different chemical groups that in-crease the solubility of curcumin and make it suitable for drug formulations [13,14]. Curcumin has inflammatory, anti-oxidant and anti-cancer properties, but is easily degraded in vitro and in vivo [15-17]. DMC is a derivative of curcumin that does not have a methoxy group attached to the benzene ring, and has similar biological properties to curcumin, but is more chemically stable [17,18]. It is known to show anti-proliferative effects on cancer cells including prostate cancer, kidney can-cer, and breast cancer [17-21]. However, DMC effects on os-teosarcoma cells are not clearly established.
In this study, therefore, the effect of DMC on cell growth and the mechanism of cell death elicited by DMC were examined in MG-63 human osteosarcoma cells. Our results showed that DMC can inhibit cell viability and induce apoptosis in a dose-dependent manner in MG-63 human osteosarcoma cells.
Materials and Methods
Materials and Methods
1. Materials
DMC (Fig. 1), 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltet-razolium bromide (MTT) and 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI) were obtained from Sigma (St. Louis, MO, USA). Anti-cleaved caspase-3, -8, -9, Fas, anti-cleaved poly (ADP-ribose) polymerase (PARP), anti-Bcl-2, anti-Bcl-xL, anti-Bax, anti-Bad, and anti-β-actin antibodies were supplied by Cell Signaling Technology, Inc. (Danvers, MA, USA). The Live/Dead cell viability assay kit was purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
2. Cell line and cell cultures
The MG-63 human osteosarcoma cells obtained from the American Type Culture Collection (ATCC, Rockville, MD, USA). The MG-63 cells were grown in Eagle’s minimum essential medium (ATCC) containing 10% fetal bovine serum (Invitrogen, Carlsbad, CA, USA) at 37°C in an atmosphere containing 5% CO2.
3. Cell viability test (MTT assay)
The MG-63 cells were seeded at a concentration of 5 × 103 cells/well in 24-well plates. After 24 hours growth, the cells were treated with DMC at various concentrations for 24 hours. The cell viability test was evaluated using the MTT assay. At least 4 separate experiments were performed on each con-centration combination. The IC50 value was estimated using a fitted line after plotting x-y and fitting the data with a straight line (linear regression). The formula is as follows: y = a × x + b, IC50 = (0.5 – b)/a, (a; slope, b; intercept).
4. Live/Dead cell assay
The MG-63 cells (1 × 105 cells/well) were cultured in an 8-well chamber slide, and allowed to attach to the bottom of the chamber slide overnight. Thereafter, the cells were treated with 0, 12.5, 25, or 50 µM DMC for 24 hours at 37℃ and stained using the Live/Dead cell viability assay kit. The cells were imaged using a fluorescence microscope (Eclipse TE2000; Nikon Instruments, Melville, NY, USA). The Live/Dead cell viability assay kit uses green calcein AM to stain the live cells (green fluorescence) and ethidium homodimer-1 to stain the dead cells (red fluorescence).
5. DAPI staining
The MG-63 cells were cultured in 24-well plates at a seed-ing density of 5 × 103 cells/well. After 24 hours growth, the cells were treated with 0, 12.5, 25, or 50 µM DMC for 24 hours. DAPI staining was done according to the previously de-scribed method [7]. The stained cells examined by fluorescent inverted microscopy (Eclipse TE2000).
6. Immunoblotting
The MG-63 cells were treated with 0, 12.5, 25, or 50 µM DMC for 24 hours. Immunoblotting was done according to the previously described method with minor modifications [22]. The cleaved caspase-3, -8, -9, Fas, anti-cleaved PARP, anti-Bcl-2, anti-Bcl-xL, anti-Bax, anti-Bad, or anti-β-actin antibody was used as the primary antibody.
7. Data analysis
All experiments were performed at least 4 times. The
re-HO
OCH3
OH
O O
sults were presented as mean ± standard error of the mean. The statistical significance was analyzed by using Student’s t-test for two groups and one way analysis of variance for multi-group comparisons. All statistical analyses were performed us-ing SPSS version 12.0 (SPSS Inc., Chicago, IL, USA). A p-value < 0.05 was considered statistically significant.
Results
Results
1. Cytotoxic effect of DMC in MG-63 cells
To analyze the effect of DMC on the viabilities of MG-63 cells, the cells were treated with DMC at various concentra-tions for 24 hours. Treatment with 25–200 µM DMC decreased the viability of MG-63 cells compared with the control in a dose-dependent manner (Fig. 2). The IC50 value of DMC on the MG-63 cell viability was approximately 54.4 µM. This re-sult suggests that DMC induces MG-63 cell death in a dose-dependent manner. Hence, in subsequent experiments were conducted at concentrations of 12.5, 25, and 50 µM, which are less than IC50.
2. Induction of apoptosis by DMC in MG-63 cells
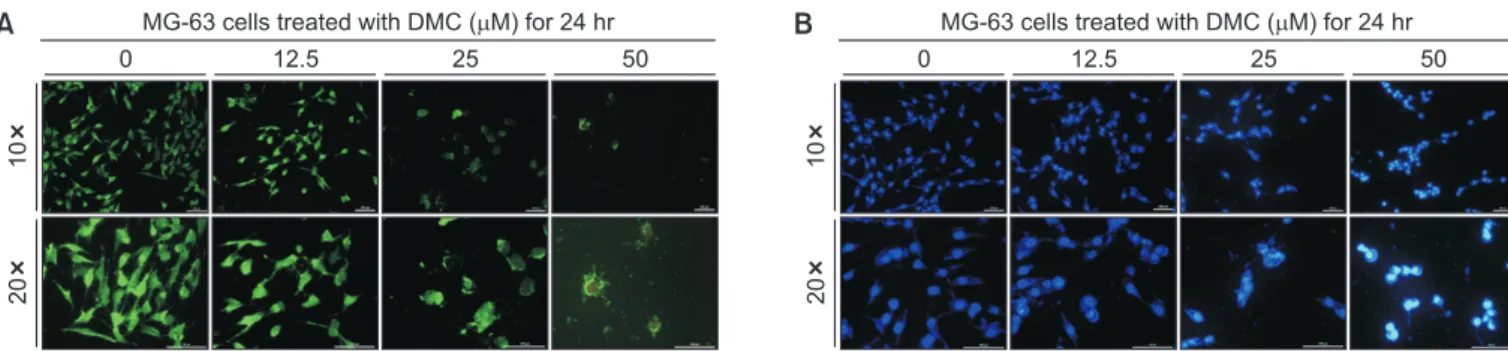
To confirm the viabilities of MG-63 cells with DMC, a Live/ Dead cell assay was performed. The MG-63 cells exposed to DMC emitted red fluorescence in a dose-dependent manner following staining with ethidium homodimer-1, which stains dead cells (Fig. 3A).
To determine whether DMC-induced MG-63 cell death is due to apoptosis, DAPI staining was performed to detect nuclear condensation, a typical feature of apoptosis. The
num-ber of MG-63 cells with condensed nuclei increased upon exposure to DMC in a dose-dependent manner, which are the characteristics of apoptosis (Fig. 3B). These results indicate that DMC induces apoptotic cell death in MG-63 cells.
3. Extrinsic and intrinsic apoptotic pathways induced
by DMC in MG-63 cells
To determine the cellular apoptotic pathways associated with DMC-induced MG-63 cell death, immunoblotting was
0 120 100 80 60 40 20 Relative cell viability (%)
MG-63 cells treated with DMC ( M) for 24 hr
0 12.5 25 50 100 200 ** *** *** ***
Fig. 2. Effects of demethoxycurcumin (DMC) on cell viability in MG-63 hu-man osteosarcoma cells. The MG-63 cells were treated with various con-centrations of DMC or without DMC for 24 hours. The cell viabilities were determined by the 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bro-mide assays. The percentage of cell viability was calculated as a ratio of A570nms of DMC treated cells and untreated control cells. Each data point
represents the mean ± standard error of the mean of four experiments. **p < 0.01 vs. control and ***p < 0.001 vs. control (the control cells mea-sured in the absence of DMC).
10
20
A
0 12.5 25 50
MG-63 cells treated with DMC ( M) for 24 hr
10
20
B
0 12.5 25 50
MG-63 cells treated with DMC ( M) for 24 hr
Fig. 3. Induction of cell apoptosis by demethoxycurcumin (DMC). (A) MG-63 cell death by DMC. The cells were treated with 0, 12.5, 25, or 50 µM DMC for 24 hours. DMC induced the death of MG-63 cells in the dose-dependent manner. Cells emitting green fluorescence are live cells stained by green calcein AM, whereas cells emitting red fluorescence are dead cells stained by ethidium homodimer-1. (B) Changes in nuclear morphology by DMC. The cells were treated with 0, 12.5, 25, or 50 µM DMC for 24 hours. 4′,6-diamidino-2-phenylindole dihydrochloride staining revealed that the number of MG-63 cells with nuclear condensation was increased by DMC.
performed. Fas, which is an apoptotic ligand that triggers the death receptor-dependent extrinsic apoptotic pathway in can-cer cells [23,24], was induced significantly by DMC in MG-63 cells (Fig. 4A). As shown in Fig. 4A, the level of cleaved caspase-8, the downstream target of pro-apoptotic factor Fas, increased following DMC treatment.
The expression levels of Bcl-2 and Bcl-xL, anti-apoptotic factors associated with the intrinsic mitochondria-dependent apoptosis pathway, were downregulated by DMC in MG-63 cells, while those of mitochondria-dependent pro-apoptotic factors such as Bax and Bad were upregulated by DMC in MG-63 cells (Fig. 4B). And, DMC treatment increased the expres-sion level of cleaved caspase-9 in MG-63 cells (Fig. 4B).
Both cleaved caspase-8 and caspase-9, acting in the ex-trinsic death receptor-mediated and inex-trinsic mitochondria-dependent apoptosis pathways, induced the expression of cleaved caspase-3 and PARP in MG-63 cells following DMC treatment (Fig. 4C).
Discussion
Discussion
DMC, which is found in turmeric, has similar biological prop-erties to curcumin, and is widely used in anticancer activity studies because of its greater chemical stability than curcumin [12,17-21]. In the present study, the cytotoxic activity and apoptotic activity of DMC were examined in MG-63 human osteosarcoma cells. The results of this study indicated that anti-proliferative activity of DMC against osteosarcoma cells was due to its ability to induce cell apoptosis.
In our cell viability test (Fig. 2) and Live/Dead cell assay (Fig. 3), DMC inhibited growth of MG-63 cells in a concentration-dependent manner. These results speculated that the DMC has cytotoxicity for osteosarcoma cells and potential value for anti-cancer drug discovery.
In this study, we examined the nuclear morphological chang-es with DAPI staining to confirm whether apoptosis is involved in the inhibition of MG-63 cell growth by DMC. The DMC in-duced the formation of nuclear condensation in MG-63 cells
Fig. 4. Intrinsic mitochondria-dependent and extrinsic death receptor-mediated apoptotic signaling pathways induced by demethoxycurcumin (DMC) in MG-63 cells. The MG-63 cells were treated with 0, 12.5, 25, or 50 µM DMC for 24 hours. The cell lysate was prepared and analyzed by immunoblotting as described in “Materials and Methods”. (A) Extrinsic death receptor-mediated apoptotic signaling pathway induced by DMC. DMC upregulated the expression level of the death receptor ligand Fas and subsequently activated the extrinsic death receptor-mediated apoptotic signaling pathway through the cleavage of caspase-8 in MG-63 cells. (B) Intrinsic mitochondria-dependent apoptotic signaling pathway induced by DMC. DMC downregulated anti-apoptotic factors Bcl-2 and Bcl-xL associated with the intrinsic mitochondria-dependent apoptotic pathway and upregulated the mitochondria-dependent pro-apoptotic factors Bax and Bad in MG-63 cells. (C) Extrinsic death receptor-mediated and intrinsic mitochondria-dependent apoptosis signaling pathways via the activation of caspase-3/-7, and poly (ADP-ribose) polymerase (PARP) induced by DMC. Cleaved caspase-8 and cleaved caspase-9 induced the activation of caspase-3 and PARP in MG-63 cells treated with DMC.
Fas Cleaved caspase-8 Cleaved caspase-8 Cleaved caspase-8 -actin 48 kDa 43 kDa 41 kDa Nonspecific band 18 kDa 45 kDa Procaspase-3 Cleaved caspase-3 Full length PARP Cleaved PARP -actin 33 kDa 17 kDa 116 kDa 45 kDa 89 kDa 0 12.5 25 50 0 12.5 25 50 DMC ( M) DMC ( M) A Bcl-2 Bcl-xL Bax Bad Procaspase-9 Cleaved caspase-9 -actin 26 kDa 30 kDa 20 kDa 47 kDa 45 kDa 37 kDa 20 kDa 0 12.5 25 50 DMC ( M) B C
(Fig. 3), suggesting apoptotic cell death by DMC. Caspase-3, -7, -8, and -9 can act as effector caspases of apoptotic cell death in eukaryotic cells [25-27]. In this study, the immunob-lotting results show that low levels of cleaved capase-3, -8, and -9 were present in DMC-untreated MG-63 cells, and the amount of cleaved enzymes was increased after DMC treat-ment in MG-63 cells (Fig. 4). These results suggested that DMC induce apoptotic cell death by the activation of caspas-es-3/-7/-8/-9 in MG-63 cells.
Fas, an important regulator of apoptosis, binds to the recep-tor FasR across the surface of the target cell, and then initi-ates the death receptor-mediated extrinsic apoptotic pathway through activation of caspase-8, -3, and PARP [23,24]. In our study, the amount of Fas protein was significantly increased by DMC in MG-63 cells (Fig. 4). In succession, the Fas stimulated by DMC triggered caspase cascade, which results in the acti-vation of apoptotic factors including cleaved caspase-8 and -3 (Fig. 4). Finally, activated caspase-3 by DMC cleaved the major substrate, PARP, leading to apoptosis in MG-63 cells (Fig. 4) [23,24]. These results indicate that DMC-induced apoptosis in MG-63 cells is mediated by the death receptor-mediated ex-trinsic apoptotic pathway through the Fas/PARP axis.
To the next, we examined the effect of DMC on the ex-pressions of Bax, Bad, Bcl-2, and Bcl-xL proteins in MG-63 cells. Pro-apoptotic proteins such as Bax and Bad, and anti-apoptotic mitochondrial proteins such as Bcl-2 and Bcl-xL are important regulators of cytochrome c release in mitochondria [27-29]. In our study, DMC treatment increased the levels of Bax and Bad protein expressions, but decreased the levels of Bcl-2 and Bcl-xL protein expressions in MG-63 cells (Fig. 4). Changes in the levels of these anti- and pro-apoptotic factors associated with the mitochondria-dependent intrinsic pathway subsequently induced the activation cascade of caspase-9, caspase-3, and PARP in MG-63 cells treated with DMC (Fig. 4). These results indicate that DMC induces apoptosis in MG-63 cells involving the death receptor- and mitochondrial-signal transduction pathways.
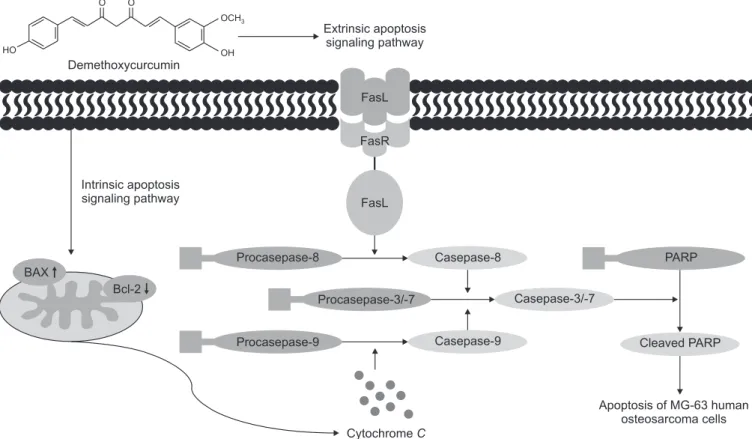
In conclusion, these results suggest that DMC inhibits cell proliferation and induces apoptotic cell death in MG-63 human osteosarcoma cells through both the death receptor-mediated extrinsic apoptotic pathway and the mitochondria-mediated intrinsic apoptotic pathway (Fig. 5). Additionally, our present findings suggest that DMC may provide a strategy to treat os-teosarcoma. HO O O OCH3 OH Demethoxycurcumin Extrinsic apoptosis signaling pathway Intrinsic apoptosis signaling pathway FasL FasR FasL CytochromeC Apoptosis of MG-63 human osteosarcoma cells Procasepase-8 Procasepase-9 Casepase-8 PARP Procasepase-3/-7 Casepase-9 Casepase-3/-7 Cleaved PARP BAX Bcl-2
Fig. 5. Apoptotic signaling pathway induced by demethoxycurcumin in MG-63 human osteosarcoma cells. PARP, poly (ADP-ribose) polymerase.
Acknowledgements
Acknowledgements
This study was supported by research fund from Chosun University (2020).
Conflicts of Interest
Conflicts of Interest
No potential conflict of interest relevant to this article was reported.
References
References
1. Eftekhari F. Imaging assessment of osteosarcoma inchild-hood and adolescence: diagnosis, staging, and evaluating re-sponse to chemotherapy. Cancer Treat Res 2009;152:33-62. doi: 10.1007/978-1-4419-0284-9_3.
2. Misaghi A, Goldin A, Awad M, Kulidjian AA. Osteosarcoma: a comprehensive review. SICOT J 2018;4:12. doi: 10.1051/ sicotj/2017028.
3. Suresh S, Saifuddin A. Radiological appearances of appen-dicular osteosarcoma: a comprehensive pictorial review. Clin Radiol 2007;62:314-23. doi: 10.1016/j.crad.2006.11.002. 4. Zhao Q, Wang C, Zhu J, Wang L, Dong S, Zhang G, Tian J.
RNAi-mediated knockdown of cyclooxygenase2 inhibits the growth, invasion and migration of SaOS2 human osteosarco-ma cells: a case control study. J Exp Clin Cancer Res 2011;30: 26. doi: 10.1186/1756-9966-30-26.
5. Cohen GM. Caspases: the executioners of apoptosis. Bio-chem J 1997;326(Pt 1):1-16. doi: 10.1042/bj3260001. 6. Hu W, Kavanagh JJ. Anticancer therapy targeting the
apop-totic pathway. Lancet Oncol 2003;4:721-9. doi: 10.1016/s1470- 2045(03)01277-4.
7. Jeong KI, Kim SG, Go DS, Kim DK. Inhibition of cell growth and induction of apoptosis by bilobalide in FaDu human pha-ryngeal squamous cell carcinoma. Int J Oral Biol 2020;45:8-14. doi: 10.11620/IJOB.2020.45.1.8.
8. Hengartner MO. The biochemistry of apoptosis. Nature 2000; 407:770-6. doi: 10.1038/35037710.
9. Kaufmann SH, Hengartner MO. Programmed cell death: alive and well in the new millennium. Trends Cell Biol 2001;11:526-34. doi: 10.1016/s0962-8924(01)02173-0.
10. Kaufmann SH, Earnshaw WC. Induction of apoptosis by cancer chemotherapy. Exp Cell Res 2000;256:42-9. doi: 10. 1006/excr.2000.4838.
11. Reed JC. Apoptosis-regulating proteins as targets for drug discovery. Trends Mol Med 2001;7:314-9. doi: 10.1016/ s1471-4914(01)02026-3.
12. Jäger R, Lowery RP, Calvanese AV, Joy JM, Purpura M, Wil-son JM. Comparative absorption of curcumin formulations. Nutr J 2014;13:11. doi: 10.1186/1475-2891-13-11.
13. Flórez KR, Shih RA, Martin MT. Nutritional fitness constructs,
measures, and links to health outcomes. In: Flórez KR, Shih RA, Martin MT, editors. Nutritional fitness and resilience: a review of relevant constructs, measures, and links to well-being. Santa Monica (CA): RAND Corporation; 2014. p. 7-24. 14. Lu Y, Wei C, Xi Z. Curcumin suppresses proliferation and
in-vasion in non-small cell lung cancer by modulation of MTA1-mediated Wnt/β-catenin pathway. In Vitro Cell Dev Biol Anim 2014;50:840-50. doi: 10.1007/s11626-014-9779-5. 15. Jurenka JS. Anti-inflammatory properties of curcumin, a
ma-jor constituent of Curcuma longa: a review of preclinical and clinical research. Altern Med Rev 2009;14:141-53.
16. Gupta SC, Patchva S, Koh W, Aggarwal BB. Discovery of curcumin, a component of golden spice, and its miraculous biological activities. Clin Exp Pharmacol Physiol 2012;39:283-99. doi: 10.1111/j.1440-1681.2011.05648.x.
17. Wu Y, Zhang P, Yang H, Ge Y, Xin Y. Effects of demethoxyc-urcumin on the viability and apoptosis of skin cancer cells. Mol Med Rep 2017;16:539-46. doi: 10.3892/mmr.2017.6666. 18. Tamvakopoulos C, Dimas K, Sofianos ZD, Hatziantoniou S,
Han Z, Liu ZL, Wyche JH, Pantazis P. Metabolism and anti-cancer activity of the curcumin analogue, dimethoxycurcumin. Clin Cancer Res 2007;13:1269-77. doi: 10.1158/1078-0432. CCR-06-1839.
19. Lin SS, Lai KC, Hsu SC, Yang JS, Kuo CL, Lin JP, Ma YS, Wu CC, Chung JG. Curcumin inhibits the migration and invasion of human A549 lung cancer cells through the inhibition of matrix metalloproteinase-2 and -9 and vascular endothelial growth factor (VEGF). Cancer Lett 2009;285:127-33. doi: 10.1016/ j.canlet.2009.04.037.
20. Chen HW, Lee JY, Huang JY, Wang CC, Chen WJ, Su SF, Huang CW, Ho CC, Chen JJ, Tsai MF, Yu SL, Yang PC. Curcumin inhibits lung cancer cell invasion and metastasis through the tumor suppressor HLJ1. Cancer Res 2008;68: 7428-38. doi: 10.1158/0008-5472.CAN-07-6734.
21. Ni X, Zhang A, Zhao Z, Shen Y, Wang S. Demethoxycurcumin inhibits cell proliferation, migration and invasion in prostate cancer cells. Oncol Rep 2012;28:85-90. doi: 10.3892/or. 2012.1783.
MicroR-NA-27 promotes odontoblast differentiation via Wnt1 signal-ing. Int J Oral Biol 2015;40:197-204. doi: 10.11620/IJOB. 2015.40.4.197.
23. Herrnring C, Reimer T, Jeschke U, Makovitzky J, Krüger K, Gerber B, Kabelitz D, Friese K. Expression of the apoptosis-inducing ligands FasL and TRAIL in malignant and benign human breast tumors. Histochem Cell Biol 2000;113:189-94. doi: 10.1007/s004180050438.
24. Li HJ, Wang CY, Mi Y, Du CG, Cao GF, Sun XC, Liu DJ, Shorgan B. FasL-induced apoptosis in bovine oocytes via the Bax signal. Theriogenology 2013;80:248-55. doi: 10.1016/j. theriogenology.2013.04.002.
25. Datta R, Kojima H, Yoshida K, Kufe D. Caspase-3-mediated cleavage of protein kinase C theta in induction of apoptosis. J Biol Chem 1997;272:20317-20. doi: 10.1074/jbc.272.33. 20317.
26. Liu X, Zou H, Slaughter C, Wang X. DFF, a heterodimeric pro-tein that functions downstream of caspase-3 to trigger DNA
fragmentation during apoptosis. Cell 1997;89:175-84. doi: 10. 1016/s0092-8674(00)80197-x.
27. Yang SJ, Lee SA, Park MG, Kim JS, Yu SK, Kim CS, Kim JS, Kim SG, Oh JS, Kim HJ, Chun HS, Kim YH, Kim DK. Induction of apoptosis by diphenyldifluoroketone in osteogenic sarcoma cells is associated with activation of caspases. Oncol Rep 2014;31:2286-92. doi: 10.3892/or.2014.3066.
28. Kluck RM, Bossy-Wetzel E, Green DR, Newmeyer DD. The release of cytochrome c from mitochondria: a primary site for Bcl-2 regulation of apoptosis. Science 1997;275:1132-6. doi: 10.1126/science.275.5303.1132.
29. Kluck RM, Esposti MD, Perkins G, Renken C, Kuwana T, Bossy-Wetzel E, Goldberg M, Allen T, Barber MJ, Green DR, Newmeyer DD. The pro-apoptotic proteins, Bid and Bax, cause a limited permeabilization of the mitochondrial outer membrane that is enhanced by cytosol. J Cell Biol 1999;147: 809-22. doi: 10.1083/jcb.147.4.809.