저작자표시-비영리-동일조건변경허락 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. l 이차적 저작물을 작성할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 동일조건변경허락. 귀하가 이 저작물을 개작, 변형 또는 가공했을 경우 에는, 이 저작물과 동일한 이용허락조건하에서만 배포할 수 있습니다.
이학
이학
이학
이학 석사학위
석사학위
석사학위
석사학위 논문
논문
논문
논문
TIM-3 발현
발현
발현
발현 조절에
조절에
조절에
조절에 중요한
중요한
중요한
중요한
DNA region 과
과
과
과
이에
이에
이에
이에 관여하는
관여하는
관여하는
관여하는 전사인자
전사인자
전사인자
전사인자 분석
분석
분석
분석
아
아
아
아 주
주
주 대
주
대
대
대 학
학
학 교
학
교
교
교
대
대
대
대 학
학
학
학 원
원
원
원
의생명과학과/분자의학전공
김
김
김
김
정
정
정
정
식
식
식
식
TIM-3 발현
발현
발현
발현 조절에
조절에
조절에
조절에 중요한
중요한
중요한
중요한
DNA region 과
과
과
과
이에
이에
이에
이에 관여하는
관여하는
관여하는
관여하는 전사인자
전사인자
전사인자
전사인자 분석
분석
분석
분석
지도교수
지도교수
지도교수
지도교수
박
박
박
박
선
선
선
선
이
이
이
이 논문을
논문을
논문을
논문을 이학
이학
이학 석사학위
이학
석사학위
석사학위 논문으로
석사학위
논문으로
논문으로
논문으로 제출함
제출함
제출함
제출함....
2009
2009
2009
2009
년
년
년
년 02
02
02
02
월
월
월
월
아
아
아
아 주
주
주 대
주
대
대
대 학
학
학 교
학
교
교
교
대
대
대
대 학
학
학
학 원
원
원
원
의생명과학과/분자의학전공
김
김
김
김
정
정
정
정
식
식
식
식
김정식의
김정식의
김정식의
김정식의
이학
이학
이학
이학
석사
석사
석사
석사
논문을
논문을
논문을
논문을
인준함
인준함
인준함
인준함
.
.
.
.
심사위원장
심사위원장
심사위원장
심사위원장
신
신
신
신
호
호
호
호
준
준
준
준
인
인
인
인
심사위원
심사위원
심사위원
심사위원
박
박
박
박
선
선
선
선
인
인
인
인
심사위원
심사위원
심사위원
심사위원
김
김
김
김
경
경
경
경
민
민
민
민
인
인
인
인
아
아
아
아 주
주
주 대
주
대
대
대 학
학
학 교
학
교
교
교
대
대
대
대 학
학
학
학 원
원
원
원
20
20
20
2008
08
08
08
년
년
년
년 12
12
12
12
월 22
월
월
월
22
22
22
일
일
일
일
- i - - 국문요약 -
TIM-3 발현
발현
발현
발현 조절에
조절에
조절에 중요한
조절에
중요한
중요한 DNA region 과
중요한
과
과
과
이에
이에
이에
이에 관여하는
관여하는
관여하는
관여하는 전사인자
전사인자
전사인자
전사인자 분석
분석
분석
분석
TIM-3 는 T 세포 immunoglobulin- and mucin-domain-containing molecules (TIMs) family 의 하나로서 보조 T 세포 1 형에서 특이적으로 발현하는 분자로
발견되었다. 이 분자는 비만세포에서도 발현이 되며 TGF-β 에 의해 발현이
증가한다고 알려졌으나 분자생물학적인 발현기작에 대해서는 연구되지 않았다. 본 연구에서는 TIM-3 의 분자생물학적인 발현조절기작을 밝히기 위해 TIM-3
upstream DNA 와 intron 1 과 2 를 각각 포함하는 luciferase reporter vector 를 제작한
후 사람 비만세포 주에서 luciferase 활성을 측정하였다. 그 결과 TIM3 upstream
-349/+142 부분이 전사활성에 있어서 중요한 역할을 하며 TGF-β 처리에 의한 TIM-3 발현 증가에서도 중요하게 작용함을 보였다. 또한 TGF-β 신호전달에
중요한 smad 2 와 smad 4 과발현 시스템을 이용하여 TGF-β 에 의한 TIM-3
발현증가에 smad 2 와 smad 4 가 관여함을 제시하였다. 본 연구의 결과는 TIM-3 전사조절에 중요한 upatream DNA region 을 밝힘으로써 TIM-3 발현 조절 기작을 이해하는데 중요한 초석이 된다.
핵심어 핵심어 핵심어
- ii -
차
차
차
차 례
례
례
례
국문요약 ··· i 차례 ··· ii 그림 차례 ··· iii 표 차례 ··· vi I. 서론 ··· 1 II. 재료 및 방법 ··· 5A. TIM-3 upstream DNA 와 intron 을 삽입한 luciferase reporter plasmid 제작 ··· 5
B. 세포배양과 plasmid 의 세포이입 ··· 6
C. Luciferase reporter assay ··· 8
D. RNA 추출과 Real-time PCR ··· 8
E. Site directed mutagenesis ··· 11
F. 통계처리 ··· 11
III. 결과 ··· 13
A. TGF-β 자극에 의한 TIM-3 mRNA 발현 증가 ··· 13
B. 인간과 생쥐의 TIM-3 regulatory DNA 의 상동성 분석 ··· 13
C. HMC-1 의 TIM-3 regulatory DNA 염기서열 분석 ··· 20
D. TIM-3 발현 조절 DNA 의 luciferase reporter vector 생산 ··· 20
E. Basal level 의 TIM-3 전사활성에 중요한 DNA region 분석 ··· 25
F. TGF-β 자극에 따른 TIM-3 전사증가에 관여하는 DNA region 분석 ··· 30
G. Smad 분자 과발현이 TIM-3 enhancer/promoter 활성에 미치는 영향 ··· 30
IV. 고찰 ··· 33
V. 결론 ··· 37
참고문헌 ··· 38
- iii -
그
그
그
그 림
림
림
림 차
차
차
차 례
례
례
례
Fig. 1. HMC-1 세포와 HEK 293 세포의 TGF-β 자극에 의한 TIM-3 mRNA 양의 변화 ··· 15Fig. 2. 사람 TIM-3 와 생쥐 tim-3 의 upstream DNA 와 intron 염기서열의
상동성 분석 ··· 17
Fig. 3. 인간 혈액/정자에서 보고된 TIM-3 유전자 염기서열과
HMC-1 에서 분리한 TIM-3 upstream DNA 의 염기서열 비교 ··· 22
Fig. 4. TIM-3 의 upstream DNA 와 intron 1 과 2 의 luciferase reporter vector 제작 ··· 23
Fig. 5. TIM-3 조절 유전자에 의한 luciferase 활성 분석 ··· 27
Fig. 6. TGF-β 자극이 TIM-3 upstream DNA 에 의한
luciferase 활성에 미치는 영향 ··· 31
Fig. 7. TIM-3 upstream DNA 에 의한 luciferase 활성에
- iv -
표
표
표
표 차
차
차 례
차
례
례
례
Table. 1. Primers for amplification of TIM-3 upstream DNA and introns ··· 7
Table. 2. Primers and probes for real-time PCR ··· 9
1
I. 서
서
서
서 론
론
론
론
TIM-3는 T 세포 immunoglobulin variable region (IgV)-like domain and mucin-domain-containing molecules (TIMs) family에 속하는 분자로서 보조 T 세포 1형 (Th1 세포)에서 특이적으로 발현되고 보조 T 세포 2형 (Th2 세포)에서는 발현되
지 않는 분자로 발견되었다 (Monney 등, 2002). TIM-3는 ligand와 결합하여 Th1 세 포 면역을 조절하고 다발 경화증이나 천식, 실험적 모델의 뇌 척수염 등의 면역 질병과 관련이 있음이 보고되고 있다 (Sabatos 등, 2003; Zhu 등, 2005).
TIM family를 구성하는 분자는 마우스 염색체 11B1.1에서 8 개, 사람 염색체 5q33.2에서 3 개가 보고되고 있다. 각각의 유전자는 서로 상동성을 보이는데, 특
히 마우스의 TIM-1과 TIM-2는 사람의 TIM-1과 각각 32 %, 42 %의 염기서열 상동 성을 보이고, 마우스의 TIM-3와 TIM-4는 사람의 TIM-3와 TIM-4와 각각 63 %,
49 %가 동일함이 확인되었다 (Kuchroo 등, 2003). 각각의 유전자는 signal sequence
에 이어 IgV-like domain과 mucin like domain, transmembrane domain과 intracellular tail 로 구성되어있다. 마우스의 TIM-1, TIM-2 그리고 TIM-3는 intrancellular tail에 타이 로신 인산화 모티프를 가지고 있는 반면 TIM-4는 타이로신이 없는 짧은
intracellular tail을 가지고 있고, 이 때문에 TIM-4는 decoy receptor로서 작용하거나
세포막에 신호 전달을 변환해주는 signaling partner를 가질 것이라 제시되고 있다
(Kuchroo 등, 2003).
TIM-3에 대한 ligand로서 galectin-9이 최근에 동정되었다. Galectin-9은 galectin family의 한 member로서 두 개의 carbohydrate-recognition domain을 가지고 있고,
이들이 carbohydrate를 인지하는데 있어서 매우 중요하다고 알려져 있다
(Matsushita 등, 2000). Galectin-9은 TIM-3와 결합을 하고 이 결합에는 TIM-3의 IgV-like domain이 중요할 것으로 생각되고 있다. 왜냐하면 TIM-3의 mucin-like domain이 없는 형태의 TIM-3가 galectin-9과 결합하였기 때문이다. 한편, TIM-3와 galectin-9의 결합이 α-lactose의 존재 하에서는 억제되므로 TIM-3의 당화가 galectin-9의 결합에 중요함이 밝혀졌다 (Zhu 등, 2005).
2
TIM-3는 비만세포에서 발현되어 이 세포의 기능을 조절한다. 비만세포에서
TGF-β에 의해 TIM-3 발현이 증가되었는데, 비만세포에서 TIM-3 ligand가 발현되
므로 TIM-3에 의한 autocrine signaling이 이 세포에서 가능하다고 제기되었다
(Wiener 등, 2007). 이를 뒷받침하는 보고는 아직 발표되지 않았다. 그러나, TIM-3
에 대한 항체 처리로 비만세포에서 interleukin-4 (IL-4), IL-6, IL-13의 생산이 증가 하고 세포 사가 억제되었으므로 TIM-3 경로가 비만세포 기능 조절에 관여하는 것 같다 (Nakae 등, 2007). TIM-3가 발견 된 이후 TIM-3 발현 조절에 대한 연구는 많이 진행되지 않았다. T 세포에서 이 세포의 활성 동안 TIM-3의 발현이 증가하고, 사람 수지세포에서 TIM-3의 발현이 이 세포의 성숙과 활성이 진행함에 따라 감소하며 정상 마우스 에 비해 TLR4 결핍 마우스의 비만세포와 대식세포에서 바이러스 감염에 의한 TIM-3 발현이 IgE 자극 혹은 TGF-β 처리에 의해 증가하였다는 보고가 있을 뿐이
다 (Yang 등, 2008; Sui 등, 2006; Frisancho 등, 2007; Wiener 등, 2007).
TGF-β는 면역반응에 있어서 다양한 역할을 한다. T 세포의 분화를 막을 뿐만
아니라, CD4+CD25- T 세포가 CD4+CD25+ regulatory T 세포로 분화가 되는 것을 촉진함으로써 대식세포, B 세포, 수지세포의 면역억제나 관용의 유지에 관여를
하고 있다 (Li 등, 2006). 또한 TGF-β는 인간 비만세포의 주화성을 자극하고 비만
세포의 분화와 기능에 영향을 미치며 FceRI의 발현을 조절하는 것으로 보고되었 다 (Olsson 등, 2000; Gebhardt 등, 2005; Gomez 등, 2005). 이 분자의 자극을 비만세 포에 오래 노출하면 세포사를 유도하고 생쥐의 점막 비만세포의 성숙에 영향을 미치는 것으로 보고되었다 (Norozian 등, 2006; Miller 등, 1999).
TGF-β의 세포 내 신호전달은 I 형과 II 형 receptor의 hetero-dimer를 이룬 복합
체에 의하여 매개된다. 7 개의 I 형 receptor와 (Activinreceptor like kinase (ALK) 1–7)
5 개의 II 형 receptor가 다양한 조합을 이루며 TGF-β sub-family member의 세포 내
신호전달에 관여한다 (Massagu´e 등, 1998). TGF-β receptor는 ligand binding에 의해
hetero-dimer를 형성하는 serine/threonine kinase로서 특히 II 형 receptor는 kinase 활
3
분자를 receptor로 유도한다 (de Caestecker 등, 2004). 인산화가 된 I 형 receptor는
TGF-β signal을 매개하는 분자로 알려진 receptor-regulated smads (smad 2와 3)를 인
산화한다 (Graff 등, 1996; Hoodless 등, 1996; Liu 등, 1996; Zhang 등, 1996).
이런 R-smad와 I 형 receptor의 상호작용에 scaffold 단백질인 smad anchor for
receptor activation (SARA)와 adaptor 단백질인 Dab2가 관여한다고 알려져 있다 (Hocevar 등, 2001; Tsukazaki 등, 1998). TGF-β에 의하여 활성이 된 receptor 복합체
는 clathrin에 의해 internalisation이 되고 SARA를 포함하는 early endosome으로 이 동한다 (Di Guglielmo 등, 2003). Receptor에 의해서 인산화 된 smad 2와 smad 3는
homomeric 또는 heteromeric 복합체를 형성하고 형성된 복합체는 다시 common mediator Smad (Co-Smad)로 알려진 Smad 4와 복합체를 형성하여 핵 안으로 이동하
고 다른 전사인자와 상호작용하여 유전자 발현에 관여하게 된다 (Lagna 등, 1996;
Zhang 등, 1996; Feng 등, 2005). R-smad와 함께 상호작용하는 전사인자로서 activating protein-1 family (Zhang 등, 1998), nuclear factor-kappaB (Lopez-Rovira 등, 2000), runt-related transcription factor-2 (Hanai 등, 1999), signal protein-1 (Moustakas 등, 1998)등이 밝혀져 있다.
Smad는 MH 1, MH2, linker domain으로 구성된 분자이다. MH 1 domain은 DNA
결합에 중요하며 nuclear localization에 중요하다 (Dennler 등, 1998; Kim 등, 1997;
Liu 등, 1997; Shi 등, 1998; Zawel 등, 1998). MH 1 domain은 MH 2의 기능을 저해하
는 역할을 한다 (Schmierer 등, 2006). Smad의 MH 2 domain은 linker domain과 함께 전사활성에 중요한 부분으로 알려져 있다 (Liu 등, 1996, 1997; Wu 등, 1997). 또한
MH 2 domain은 R-smad와 receptor의 상호작용과 smad간의 복합체 형성, smad와 SARA 복합체 형성, smad와 다양한 전사인자의 복합체 형성에 중요한 역할을 한
다 (Bai 등, 2000; Chen 등, 1998; Hayashi 등, 1997; Lo 등, 1998; Chacko 등, 2001; Shi 등, 1997; Wu 등, 2001, 1997; Wu 등, 2000; Massague 등, 2005). 한편 smad 3와 smad 4 가 DNA에 biding하는 element (smad-binding element: SBE)는 AGAC 혹은 그에 상 보적인 GCTC이다. Smad의 SBE binding affinity는 단독으로 SBE가 존재하는 것보 다 국소적으로 여러 개의 SBE가 모여있을 때 affinity가 높다 (Zawel 등, 1998).
4
Smad 2는 DNA에 binding을 할 수 있는 motif가 없기 때문에 smad 3 혹은 smad 4
와 complex를 형성해야 전사인자로서의 기능을 할 수 있다 (Dennler 등, 1999).
TIM-3 분자의 발현 조절 기전을 이해하는 것은 면역반응 이해에 중요함에도
불구하고 깊이 연구되지 않았으므로 본 연구에서는 비만세포에서 TGF-β에 의해
TIM-3 발현이 증가할 때 이에 관여하는 TIM-3 DNA region과 전사인자를 분석하
5
II. 실험
실험
실험
실험 재료
재료
재료 및
재료
및
및
및 방법
방법
방법
방법
A. TIM-3 upstream DNA와와와 intron 을와 을을 삽입한을 삽입한삽입한삽입한 luciferase reporter plasmid 제작제작제작 제작
G-DEX Genomic DNA extraction kit (Intron, Sungnam, Korea)를 이용하여 1 x 107
개의 HMC-1 (Human mast cell-1)에서 genomic DNA를 분리하였고 이를 주형으로
TIM-3에 대한 primer를 이용하여 중합효소 연쇄반응 (polymerase chain reaction; PCR)을 시행하였다. TIM3 +1과 TIM3 -2000 primer를 이용하여 TIM-3 upstream DNA -1677 ~ +144 부분을, TIM3 +1과 TIM3 -1000 primer를 이용하여 TIM-3 upstream DNA -872 ~ +144 부분을, TIM3 -2500과 anti TIM3 -2000 primer를 이용하여 TIM-3 upstream DNA -2238 ~ -1634 부분을 각각 증폭하였다. 또한 TIM3-intron1 (5)과 intron1 (3) primer를 이용하여 TIM-3 intron 1을, intron2 (5)와 TIM3-intron2 (3) primer를 이용하여 TIM-3 intron 2를 각각 증폭하였다 (Table 1). rTaq (Bioneer, Daejeon, Korea)을 이용하여 다음과 같은 조건으로 PCR을 수행하였다. 94℃에서 5 분 정치 후에, 94℃ 30초, 67℃ 30초, 72℃ 1분으로 25 cycles를 수행하
였다. PCR 산물을 Topo TA cloning vector (Invitrogen, Carsbade, CA, U.S.A.)에 클로닝 하고 sequence를 확인하였다. TIM-3 upstream DNA -349 ~ +144 부분을 증폭하기 위 해서 확보된 TIM-3 upstream DNA -1677 ~ +144 부분을 주형으로 TIM3 +1과 sense
TIM3 -500 primer를 이용하여 rTaq (Bioneer, Daejeon, Korea)을 이용하여 다음과 같
은 조건으로 PCR을 수행하였다. 94 ℃에서 5 분 정치 후에, 94 ℃ 30초, 67 ℃
30초, 72 ℃ 1분으로 25 cycles를 수행하였다. PCR 산물을 Topo TA cloning vector (Invitrogen, Carsbade, CA, U.S.A.)에 클로닝하고 sequence를 확인하였다. 각 TIM-3 DNA 조각을 firefly luciferase reporter vector인 pGL-Basic vector (Promega, Madison, WI, U.S.A.)와 pGL-Promoter vector (Promega, Madison, WI, U.S.A.)에 삽입하였다.
TIM-3 upstream DNA -1677 ~ +144 부분과 TIM-3 upstream DNA -872 ~ +144 부분, TIM-3 upstream DNA -349 ~ +144 부분을 pGL-basic vector의 Nhe I / Bgl II site에 삽입 하여 각각 T3U(1.8)-luc, T3U(1.0)-luc, T3U(0.5)-luc으로 명명하였다. T3U(1.8)-luc의
6
Kpn I/Xho I site에 Intron 1과 Intron 2를 삽입하여 각각 T3U(1.8/I1)-luc,
T3U(1.8/I2)luc으로 명명하였다. pGLPromoter vector의 Kpn I/Xho I site에 TIM3 upstream DNA -2238 ~ -1634 부분과 Intron 1, Intron 2를 삽입하여 각각 T3U(e0.6)-luc-p, T3(I1)-luc-p, T3(I2)-luc-p으로 명명하였다.
B. 세포배양과세포배양과세포배양과세포배양과 plasmid의의의의 세포이입세포이입세포이입세포이입
HMC-1 세포의 배양에는 Iscove’s modified Dulbecco’s medium (IMDM), HEK293
세포의 배양에는 Dulbecco’s modified Eagle’s medium (DMEM)을 사용하였다. 각 배
지에 10%의 우태아혈청 (fetal bovine serum; FBS)과 1%의 penicillin/streptomycin
(PS)이 포함되며 세포는 37℃, 5% CO2 배양기에서 배양하였다.
HMC-1 세포에 luciferase reporter vector의 한시적 이입 (transient transfection)을
전기천공기 (Digital Bio Technology, Seoul, Korea)를 이용한 electroporation으로 수행
하였다. 1 x 106
개의 세포를 1 ml의 PBS로 세척을 한 후, 100 ul Resuspension buffer (Digital Bio Technology, Seoul, Korea)로 부유시키고, 여기에 3.6 ug의 luciferase reporter vector와 400 ng의 pEGFP-N1 plasmid (Clonetech, Mountain View, CA, U.S.A.)
를 넣은 후, 전기충격을 가하였다. 이를 2 ml의 배지가 들어있는 24 well plate에
다시 부유시킨 다음 37 , 5% CO℃ 2 배양기에서 48 시간 배양하였다.
HEK293 세포에 luciferase reporter vector의 이입에는 Lipofectamin 2000 (Invitrogen, Carsbade, CA, U.S.A.)이 이용되었다. 먼저 5 x 105 개의 세포를 0.5%의 FBS를 포함하는 DMEM에 부유하여 6 well plate에 넣고 밤새 배양하였다. 한편 luciferase reporter vector 3.6 ug과 pEGFP-N1 400 ng을 1 mg/ml 농도의 Lipofectamin 2000 reagent 13 ul와 혼합한 후, 100 ul의 Opti-MEM에 넣고 잘 섞어준 다음 30 분
간 실온에서 정치하였다. 이를 HEK293 세포에 넣고 6 시간 정치 한 후, 배지를 교환하고 42 시간 배양하였다.
7
Table 1. Primers for amplification of TIM-3 upstream DNA and introns
Primer Nucleotide sequence
TIM3 +1 GGAGCTTGCAGAAGAAAAGTCAGAGGACACCTCTGTTAGG TIM3 -1000 CTTTTGCTTTTAAGGTGTCCAGATAAAGGTCACACTCCCAG TIM3 -2000 AGAGCCTTGACCAAGTTCATGCTGCTAATAAAAATAACCCCA G TIM3 -2500 GGAAATAAAAGTTAAGTGAACATGAAAATGTTCAGAACCAG CTCC
anti TIM3 -2000 CTGGGGTTATTTTTATTAGCAGCATGAACTTGGTCAAGGCTCT sense TIM -500 CTGTGACCAAAGTTTATGAAGCC
TIM3-intron1 (5) GTAAGTCTCGGCATGGATATTTACAATGACATAATGGTGCTG TIM3-intron1 (3) CTGCATAGAGAGAGAAGGAGAGCCAAGACTCAAGAGG TIM3-intron2 (5) GTGAGTGGACATTTGCATGCCATCTTTATGAATAAGATTTATCT G TIM3-intron2 (3) CTAATGTCAGAAACAACATAAGGATGAAAATTATCTGAGAGC AGAAAGC
8 C. Luciferase reporter assay
Luciferase reporter plasmid가 이입된 세포를 PBS로 세척한 다음 1x Lysis buffer (Promega, Madison, WI, U.S.A.) 100 ul를 넣고 3분간 정치시켰다. 세포 용해액을 96 well luminometer plate에 넣은 후 100 ul의 luciferin (Promega, Madison, WI, U.S.A.)을
넣고 반응시킨 후 luminometer (Molecular devices, Sunnyvale, CA, U.S.A.)에서
luciferase activity를 측정하였다. Luciferase activity를 측정하기 전에 plasmid의 세포
이입 정도를 보정하기 위해 GFP 발현 vector의 이입에 의한 GFP 발현 빈도를 형 광활성 세포분류기 (fluorescence activated cell sorter; FACS)를 통하여 측정하였다.
D. RNA 추출과추출과추출과추출과 real-time PCR
시간 별로 TGF-β의 자극을 준 세포에서 RNA를 추출하기 위해서 RNA STAT-60TM
(Tel-Test, Friendswood, TA, U.S.A.)을 사용하였다. 300 ul의 RNA STAT-60TM를 세포에
첨가하여 5 분간 상온에서 정치하여 세포를 lysis한 후 60 ul의 chloroform을 첨가
하여 5 분간 상온에서 정치하였다. 13000 rpm, 4 ℃, 15 분의 조건으로 원심분리를
하여 상층액을 새로운 tube에 옮기고 isopropanol을 사용하여 RNA를 침전시켰다.
75 % ethanol으로 RNA를 세척하고 diethyl pyrocabonate (DEPC)가 처리된 증류수를
사용하여 RNA를 녹여 정량하였다. 추출 된 1.4 ug의 RNA를 주형으로 oligo-dT
(Invitrogen, Carsbade, CA, U.S.A.)와 역 전사 효소 (Invitrogen, Carsbade, CA, U.S.A.)
를 이용하여 cDNA를 생산하였다. 0.5 ug의 oligo-dT와 10 mM dNTP 1 ul, DEPC 증 류수 13.7 ul을 RNA와 혼합한 후 65 ℃에서 5 분, 4 ℃에서 5 분간 각각 정치시켰 다. 이 혼합액에 4 ul의 5x first standard buffer (Invitrogen, Carsbade, CA, U.S.A.)와 2
ul의 0.1 M DTT, 0.3 ul의 역 전사효소를 첨가하여 42 ℃에서 52 분간 반응을 시켜 cDNA를 합성하였다. 생산 된 cDNA를 주형으로 TIM-3 mRNA의 발현 정도를 real-time PCR과 ABI PRISM 7000 system (Applied Biosystems, Foster city, CA, U.S.A.)을
9 Table 2. Primers and probes for real-time PCR
Primer and probe Nucleotide sequence TIM-3 real-time forward TCCAAGGATGCTTACCACCAG TIM-3 real-time reverse GCCAATGTGGATATTTGTGTTAGATT TIM-3 FAM probe ACATGGCCCAGCAGAGACACAGACACT GAPDH real-time forward TATTGTTGCCATCAATGACCCCTTCATTGA GAPDH real-time reverse CATATTGGAACATGTAAACCATGTAGTTG GAPDH FAM probe TATTGTTGCCATCAATGACCCCTTCATTGA
10
18 pmole의 TIM-3 real-time PCR forward/reverse primer, 2x gene expression mastermix (Applied Biosystems, Foster city, CA, U.S.A.) 10 ul와 0.5 ul의 FAM으로 표지 된 TIM-3 probe가 사용되었으며 Two-step PCR protocol (95 ℃에서 15 초, 60 ℃에서 1분: 40 cycles)이 사용되었다.
11 E. Site directed mutagenesis
TIM-3 upstream region -1094 bp에 변이 (A → G)가 생긴 T3U(1.8)-luc 9번 클론의
변이교정을 위해서 QuikChange® II Site-Directed Mutagenesis Kit site (Stratagene,
Cedar Creek, TA, U.S.A.)를 사용하여 site directed mutagenesis를 수행하였다. T3U(1.8)-luc 9번 클론을 주형으로 TIM-3 UTR mutation(5) primer와 TIM-3 UTR mutation(3) primer를 이용하여 중합효소 연쇄반응 (polymerase chain reaction; PCR)을
시행하였다 (Table 3). Pfu polymerase (Stratagene, Cedar Creek, TA, U.S.A.)를 이용하여 다음과 같은 조건으로 PCR을 하였다. 95 ℃에서 30 초 정치 후에, 95 ℃ 30 초,
65 ℃ 1 분, 72 ℃ 5 분으로 12 cycles를 수행하였다. PCR 산물을 Dpn I (Stratagene, Cedar Creek, TA, U.S.A.)으로 37 ℃에서 1 시간 반응시킨 후 transformation을 하였
다. DH5α competent cell에 PCR 산물 1 ul를 넣고 ice에서 30 분 정치시킨 후 42 ℃
에서 45 초간의 heat shock을 주었다. 다시 ice에 2 분간 정치시킨 다음 1 ml의 LB
broth를 넣어준 후 1 시간 배양을 하였다. 배양액을 LB plate (Amp+)에 spreading
한 다음 37 ℃에서 밤새 배양을 하고 집락을 LB broth에서 배양을 하여 교정 된
T3U(1.8)-luc을 준비하였다. 준비 된 plasmid의 변이교정을 sequencing을 통해 확인
하였다.
F. 통계처리통계처리통계처리 통계처리
실험군과 대조군의 통계학적인 분석은 paired-t test를 사용하여 p값이 0.05 미만
12 Table 3. Primers for site directed mutagenesis
Primer and probe Nucleotide sequence
TIM3 UTR mutation(5) GAATGACAAAGAAACTCAGGAAGGCAAGACATAGAGGA AG
13
III. 결
결
결
결 과
과
과
과
A. TGF-β 자극에자극에자극에 의한자극에 의한의한 TIM-3 mRNA 발현의한 발현발현발현 증가증가증가증가 사람에서 분리한 비만세포에서 TGF-β 처리에 의해 TIM-3 발현이 증가되었다 고 보고되어 있으나, 사람 비만 세포 주 HMC-1에서 이러한 현상이 조사되지 않 았으므로 먼저 HMC-1에서 TIM-3의 발현을 분석하였다. HMC-1 세포에서 TIM-3mRNA가 발현되고 있으며 TGF-β (2 ng/ml)를 4 시간 처리하였을 때 TIM-3 mRNA
의 발현이 약 3.5 배 정도 증가 하는 것을 관찰하였다 (Fig. 1A). HMC-1 세포에서
TIM-3가 발현되고 TGF-β에 의해서 발현이 증가되므로 HMC-1 세포가 TIM-3 발
현 조절 연구에 적합한 세포로 판단되었다. 또한 사람 신장세포에서 유래된 세 포주인 HEK293 세포의 TIM-3 발현을 분석하였다. HEK293 세포에서는 TIM-3
mRNA가 발현하지만 TGF-β (2 ng/ml) 처리에 의한 TIM-3 mRNA 발현증가는 관찰
되지 않았다 (Fig. 1B).
B. 인간과인간과인간과인간과 생생생쥐의생쥐의 TIM-3 regulatory DNA의쥐의쥐의 의의의 상동성상동성상동성 분석상동성 분석분석분석
TIM-3 유전자 전사활성 부분을 예측하기 위하여 인간 TIM-3 (NW_001838954)와
생쥐 tim-3 (NW_001030461)의 upstream DNA 염기서열과 intron 염기서열의 상동성 을 분석하였다. TIM-3 transcription start site를 기점으로 약 5 Kbp upstream DNA와
1.9 Kbp의 intron 1과 1.8 Kbp의 intron 2를 NCBI bl2seq BLAST program을 사용하여
상동성 분석을 한 결과, upstream DNA와 intron 1에서 생쥐의 tim-3와 상동성이 높 은 부분이 관찰되었다. 인간 TIM3 upstream DNA의 370 ~ 310의 60 bp와 632 ~
584의 48 bp DNA가 생쥐의 tim3 upstream DNA의 432 ~ 377의 55 bp와 1000 ~ -951의 49 bp의 염기서열과 각각 83 %, 86 %의 상동성을 보였다 (Fig. 2A). 또한 인
간 TIM-3 intron 1의 +247 ~ +304의 57 bp의 염기서열이 생쥐의 tim-3 intron 1의
14 3은 인간의 TIM-3 유전자의 +5625 ~ +5688 부분과 생쥐의 tim-3 유전자 +4308 ~ +4362 부분이 87 %, 인간의 TIM-3 유전자의 +7907 ~ +7961 부분과 생쥐의 tim-3 유전자 +4314 ~ +4367 부분이 85 %, 인간의 TIM-3 유전자의 +7954 ~ +8001 부분과 생쥐의 tim-3 유전자 +4314 ~ +4360 부분이 8 7%, 인간의 TIM-3 유전자의 +4655 ~ +4710 부분과 생쥐의 tim-3 유전자 +4308 ~ +4362 부분이 83 %, 인간의 TIM-3 유 전자의 +4805 ~ +4857 부분과 생쥐의 tim-3 유전자 +4308 ~ +4359 부분이 84 %, 인 간의 TIM-3 유전자의 +8033 ~ +8083 부분과 생쥐의 tim-3 유전자 +4308 ~ +4356 부분이 88 %의 상동성이 관찰되었다 (Fig. 2C). Intron 4는 인간의 TIM-3 유전자의
+12888 ~ +12947 부분과 생쥐의 tim-3 유전자 +12484 ~ +12542 부분이 85 %, 인간
의 TIM-3 유전자의 +12335 ~ +12385 부분과 생쥐의 tim-3 유전자 +12484 ~ +12533 부분이 86 %, 인간의 TIM-3 유전자의 +12888 ~ +12943 부분과 생쥐의 tim-3 유전 자 +14104 ~ +14158 부분이 83 %의 상동성이 관찰되었다 (Fig. 2D). Intron 6는 인간 의 TIM-3 유전자의 +20271 ~ +20342 부분과 생쥐의 tim-3 유전자 +21192 ~ +21260 부분이 79 %의 상동성이 관찰되었다 (Fig. 2E). 그러나 intron 2와 intron 5는 상동성 이 관찰되지 않았다. 비록 intron 2는 종간의 상동성을 관찰하지 못하였지만
TIM-3 regulatory DNA에 GATA가 binding할 가능성을 제시한 이전의 보고를 근거로 (Chae 등, 2004) TIM-3 regulatory DNA의 전사인자 binding motif를 분석한 결과, intron 2에 GATA가 binding을 할 가능성이 높기 때문에 intron 2의 luciferase reporter vector를 제작하였다. 이 분석을 바탕으로 추측해볼 때 TIM-3 upstream의 -310 ~ - 632 부분, intron 1 의 +247 ~ +304 부분이 TIM-3 전사에 중요하게 작용할 가능성
15 A.
16
Fig. 1. HMC-1 세포세포세포와세포와와와 HEK 293 세포의세포의세포의 TGF-세포의 β 자극에자극에 의한자극에자극에 의한의한의한 TIM-3 mRNA 양양양양의의의 변의 변변변
화 화화
화. (A) HMC-1 세포에 4 시간의 TGF-β (2 ng/ml) 자극을 준 후, TIM-3 mRNA 양의
변화를 real-time PCR로 측정하였다. (B) HEK293 세포에 24 시간의 TGF-β (2 ng/ml)
17 A.
18 C.
19 E.
Fig. 2. 사람사람사람 TIM-3와사람 와와 생쥐와 생쥐생쥐생쥐 tim-3의의 upstream DNA와의의 와와와 intron 염기서열의염기서열의염기서열의염기서열의 상동성상동성상동성상동성 분분분분
석 석석
석. (A) 인간과 생쥐의 TIM-3 upstream DNA의 상동성 분석. (B) 인간과 생쥐의 TIM-3 intron 1 DNA의 상동성 분석. (C) 인간과 생쥐의 TIM-3 intron 3 DNA의 상동 성 분석. (D) 인간과 생쥐의 TIM-3 intron 4 DNA의 상동성 분석. (E) 인간과 생쥐의 TIM-3 intron 6 DNA의 상동성 분석.
20
C. HMC-1의의의의 TIM-3 regulatory DNA 염기염기염기염기서열서열서열 분석서열 분석분석분석
HMC-1의 TIM-3 upstream DNA와 intron 1과 intron 2를 각각 클로닝 한 후 이의
염기서열을 genebank에 등록되어 있는 인간 혈액/정자의 TIM-3 유전자
(NW_001838954)와 비교하였다. 먼저 각 10개의 클론의 염기서열을 비교하여 서
로 일치하지 않는 염기서열을 클로닝 과정에서 발생한 변이로 간주하였고, 10개 클론에서 모두 일치하는 염기서열을 genebank에 등록된 TIM-3 염기서열과 비교 하였다. 2.2 Kbp의 TIM3 upstream DNA의 염기서열 중 1636 bp, 1189 bp, 782 bp,
574 bp에서 HMC1과 혈액/정자의 TIM3 염기서열에 차이가 있었으며 HMC1의 -1471 bp, -1188 bp에서 각각 C와 A nucleotide의 삽입이 관찰되었다. 이 중 -574 bp (T > G)는 이미 보고된 TIM-3 polymorphism (rs10515746)으로 분석되었다 (Fig. 3A).
한편, 1.9 Kbp의 intron 1에서 혈액/정자의 TIM-3 유전자와 HMC-1의 TIM-3 유전자 의 염기서열이 모두 일치하였다. 한편 1.8 Kbp의 intron 2 염기서열 중 +2776 bp,
+2821 bp, +2850 bp, +3043 bp, +3067 bp, +3630 bp, +3759 bp, +3760 bp, +4400 bp에서 HMC-1과 혈액/정자의 TIM-3 염기서열이 차이가 있었으며 HMC-1의 +2849 bp에
서 G nucleotide의 삽입과 +4173 bp, +4174 bp에서 nucleotide 결실이 관찰되었다
(Fig. 3B).
D. TIM-3 발현발현발현발현 조절조절조절조절 DNA의의의의 luciferase reporter vector 생산생산생산 생산
TIM-3의 전사에 TIM-3 genomic DNA의 어느 부분이 중요한 역할을 하는지 알
아보기 위해 TIM-3 upstream DNA와 intron 1, intron 2를 포함하는 luciferase reporter
vector를 제작하였다 (Fig. 4A). 클로닝이 된 luciferase reporter vetor의 확인을 위해 vector DNA를 제한효소 처리한 후 DNA 절편의 크기를 분석하였다. EcoR I을 처리
한 T3U(1.8)-luc, T3U(1.0)-luc, T3U(0.5)-luc, T3U(e0.6)-luc-p, T3(I1)-luc-p, T3(I2)-luc-p 는 각각 약 4.8 Kbp의 DNA band와 삽입한 TIM-3 DNA 크기의 band로 분리되었으 므로 luciferase reporter vector가 제대로 만들어졌음을 알 수 있었다. T3U(1.8/I1)-luc
21
와 T3U(1.8/I2)-luc는 약 5.8 Kbp DNA 절편과 2.8 Kbp DNA 절편으로 자르는 Kpn I 과 Sal I을 사용하여 분석하였고, 예상크기의 band를 관찰함으로써 T3U(1.8)-luc
vector에 intron 1과 intron 2가 각각 삽입된 T3U(1.8/I1)-luc과 T3U(1.8/I2)-luc이 만들
22 A.
B.
Fig. 3. 인간인간인간 혈액인간 혈액혈액혈액/정자정자정자정자에서에서에서 보고된에서 보고된 TIM-3 유전자보고된보고된 유전자유전자유전자 염기서열과염기서열과염기서열과 HMC-1에서염기서열과 에서에서에서 분리한분리한분리한 분리한 TIM-3 upstream DNA의의의의 염기서열염기서열염기서열염기서열 비교비교비교비교. (A) TIM-3 upstream DNA의 약 2.2 Kbp내의 인간 혈액/정자와 HMC-1의 TIM-3 염기서열 상동성 분석 (TIM-3 GenBank
accession no. NW_001838954) (B) TIM-3 intron 2 약 1.8 Kbp내의 인간 혈액/정자와 HMC-1의 TIM-3 염기서열 상동성 분석. * : 결실
23 A.
24
Fig. 4. TIM-3의의의의 upstream DNA와와와와 intron 1과과과과 2의의의의 luciferase reporter vector 제작제작제작제작 (A)
여러가지 TIM-3 luciferase reporter vector의 지도. (B) 제한효소 처리에 의한 TIM-3
luciferase reporter vector의 분석. T3U(1.8/I1)-luc과 T3U(1.8/I2)-luc은 Kpn I과 Sal I으
25
E. Basal level의의의 TIM-3 전사활성에의 전사활성에전사활성에전사활성에 중요한중요한중요한중요한 DNA region 분석분석분석분석
HMC-1 세포와 HEK293 세포는 자극이 없는 조건에서 TIM-3 mRNA를 발현
하므로 이러한 basal level의 전사에 중요한 regulatory DNA region을 알아보고자 하 였다. 먼저 DNA 이입이 비교적 쉬운 HEK293 세포에 TIM-3 luciferase reporter
vector를 이입하여 luciferase 활성을 분석하였다. pGL-Basic vector가 이입된 대조군
에 비해 T3U(0.5)-luc가 이입된 경우 약 10.5 배의 luciferase activity가 관찰되었으 며, T3U(1.0)-luc가 이입된 경우 약 5.6 배, T3U(1.8)-luc가 이입된 경우 약 2.5 배의
luciferase activity가 관찰되었다. 이는 T3U(0.5)-luc에 삽입된 TIM-3 upstream의 -349 ~ +144 부분이 basal level의 TIM-3 전사활성에 중요함을 의미한다. 한편 T3U(1.8)-luc이 이입된 경우에 비해 T3U(1.8/I1)-luc가 이입된 경우 luciferase activity 증가는
약 3 배, T3U(1.8/I2)-luc가 이입된 경우 약 3.6 배가 관찰되었다. 그렇지만
T3U(e0.6)-luc-p와 T3(I1)-luc-p, T3(I2)-luc-p가 이입된 경우의 lucifease activity는 pGL-Promoter vector가 이입된 대조군에 비해서 크게 차이가 없었다 (Fig. 5A). 이
는 TIM-3 전사에 intron 1과 intron 2가 작용하며 이들이 작용하기 위해서는 TIM-3
upstream DNA region이 필요함을 제시한다. 다음으로 이러한 결과가 HMC-1 세포
에서도 유사하게 나타나는지 알아보고자 실험하였다. HMC-1 세포에 TIM-3
luciferase reporter vector를 이입하여 luciferase 활성을 분석하였다. pGL-Basic vector
가 이입된 대조군에 비해 T3U(0.5)-luc가 이입된 경우 약 5.5 배의 luciferase
activity가 관찰되었으며, T3U(1.0)-luc가 이입된 경우 약 3 배, T3U(1.8)-luc가 이입
된 경우 약 1.7 배의 luciferase activity가 관찰되었다. 이는 HMC-1 세포에서도
T3U(0.5)-luc에 삽입된 TIM-3 upstream의 -349 ~ +144 부분이 basal level의 TIM-3 전
사활성에 중요함을 의미한다. 한편 T3U(1.8)-luc이 이입된 경우에 비해
T3U(1.8/I1)-luc가 이입된 경우 luciferase activity 증가는 약 1.3 배, T3U(1.8/I2)-luc가
이입된 경우 luciferase activity는 약 1.7 배가 관찰되었다. 또한 T3U(e0.6)-luc-p와
T3(I1)-luc-p, T3(I2)-luc-p가 이입된 경우의 lucifease activity는 pGL-Promoter vector가
26
level TIM-3 전사에 intron 1과 intron 2가 의미 있게 작용하지 않을 가능성을 제시
27 A.
28 B.
29
Fig. 5. TIM-3 조절조절조절조절 유전자에유전자에유전자에 의한유전자에 의한 luciferase 활성의한의한 활성활성활성 분석분석분석 (A) 5 x 10분석 5 개의 HEK293 세
포 혹은 (B) 1 x 106
HMC-1 세포에 각 luciferase reporter vector와 pEGFP-N1 vector(Total plasmid : pEGFP-N1 = 10 : 1)를 이입하고 48 시간 후 luciferase 활성을
측정하였다. pGL-Basic 또는 pGL-Promoter가 이입된 세포의 luciferase 활성을 기준 으로 하여 각 TIM-3 조절유전자의 luciferase reporter vector가 이입된 세포의
luciferase 활성을 계산하였다. DNA 이입효율보정을 위해 GFP 발현세포빈도를 FACS로 분석하였다. 결과는 5회 이상 실험하여 얻은 값을 평균 ± 표준편차로 표시하였다. *P<0.001, **P<0.01, ***P<0.0001
30
F. TGF-β 자극에자극에자극에 따른자극에 따른따른따른 TIM-3 전사증가에전사증가에 관여하는전사증가에전사증가에 관여하는관여하는 DNA region분석관여하는 분석분석분석
HMC-1 세포에서 TGF-β 자극에 의한 TIM-3 전사증가에 관여하는 DNA region
을 알아보고자 HMC-1 세포에 각각의 luciferase reporter vector를 이입하고 6 시간
의 TGF-β (2 ng/ml) 자극을 준 후에 luciferase 활성을 측정하였다. TGF-β 자극에 의
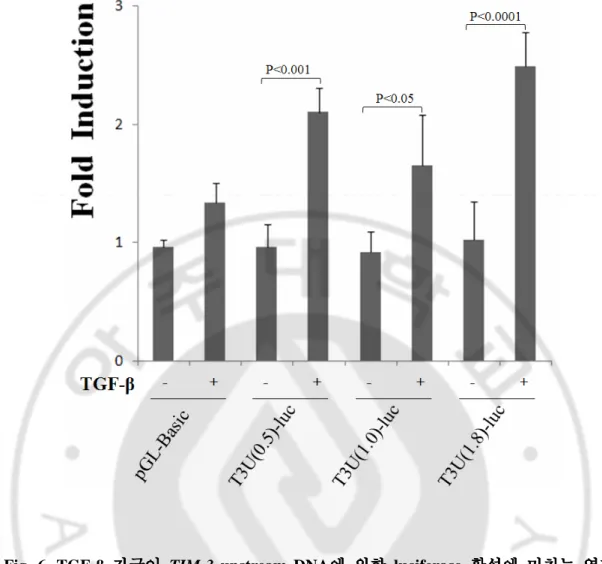
해 T3U(0.5)-luc의 luciferase activity는 약 2.3 배, T3U(1.0)-luc의 luciferase activity는 약 1.8 배, T3U(1.8)-luc의 luciferase activity는 약 2.6 배 증가하는 것을 관찰하였다
(Fig. 6). 따라서 HMC-1에서 TGF-β 자극에 의한 TIM-3 전사증가가 TIM-3 upstream
의 -349 ~ +144 부분으로도 충분하다고 생각된다.
G. Smad 분자분자분자분자 과발현이과발현이과발현이 TIM-3 enhancer/promoter 활성에과발현이 활성에활성에 미치는활성에 미치는미치는미치는 영향영향영향영향
TGF-β에 의하여 활성이 되는 smad 2와 3은 homomeric complex 혹은
heteromeric complex를 형성하여 smad 4와 복합체를 형성한 후 TGF-β의 target gene
의 발현에 관여한다는 보고가 있다 (Lagna 등, 1996; Zhang 등, 1996). 이에 TGF-β
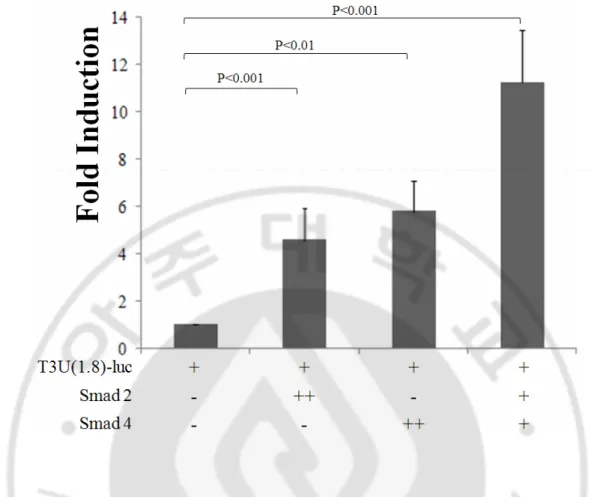
자극에 의한 TIM-3 전사증가에 smad 분자가 관여하는지 알아보고자 smad를 과 발현 하도록 한 HEK293 세포에서 luciferase reporter assay를 진행하였다. 빈 vector 를 이입한 대조군에 비해 smad 2 를 과발현 시킨 경우 luciferase activity는 4.5 배,
smad 4 를 과발현 시킨 경우 luciferase activity는 5.8 배 증가하였다. Smad 2와 smad 4를 함께 과발현 시킨 경우 대조군에 비해서 11 배, smad 2 단독 혹은 smad 4 단독을 이입한 경우보다 각각 2.4 배, 1.8 배 높은 luciferase activity가 관찰되었
다 (Fig. 7). 이 결과는 smad 2와 smad 4가 TIM-3 발현에 관여하며 smad 2와 smad 4 가 협동적으로 TIM-3 전사에 역할을 할 가능성을 보여준다.
31
Fig. 6. TGF-β 자극자극자극이자극이이이 TIM-3 upstream DNA에에에 의한에 의한 luciferase 활성에의한의한 활성에활성에 미치는활성에 미치는미치는미치는 영향영향영향영향.
1 x 106 HMC-1 세포에 각 luciferase reporter vector와 pEGFP-N1 vector (Total plasmid : pEGFP-N1 = 10 : 1)를 이입하여 6 시간의 TGF-β (2 ng/ml) 자극을 주고 48 시간 후 luciferase 활성을 측정하였다. TGF-β가 처리되지 않은 세포의 luciferase 활성을 기 준으로 하여 TGF-β가 처리된 경우의 luciferase 활성을 계산하였다. DNA 이입효율 보정을 위해 FACS로 분석한 GFP 발현세포빈도로 luciferase 활성을 보정하였다. 결과는 5회 이상 실험하여 얻은 값을 평균 ± 표준편차로 표시하였다.
32
Fig. 7. TIM-3 upstream DNA에에에에 의한의한의한 luciferase 활성에의한 활성에 smad 과발현이활성에활성에 과발현이과발현이과발현이 미치는미치는미치는미치는 영향영향영향영향.
1 x 106 HEK293 세포에 각 T3U(1.8)-luc과 smad 2 또는 smad 4를 발현하는 plasmid, pEGFP-N1 vector를 이입하고 48 시간 후 luciferase 활성을 측정하였다. DNA 이입
효율보정을 위해 pEGFP-N1 vector를 Total plasmid : pEGFP-N1 = 10 : 1의 비로 세포 에 이입하였으며 GFP 발현세포빈도를 FACS로 분석하였다. T3U(1.8)-luc과 빈 벡
터가 이입된 세포의 luciferase 활성을 기준으로 하여 smad 2와 smad 4가 발현하는
세포의 luciferase 활성을 계산하였다. 결과는 5회 이상 실험하여 얻은 값을 평균 ± 표준편차로 표시하였다.
33
IV. 고
고
고
고 찰
찰
찰
찰
본 연구에서는 TIM-3 전사에 중요한 TIM-3 upstream DNA와 그에 관여하는 전사인자를 알아보고자 하였다. Basal level의 TIM-3 전사에 -349 ~ +144 부분이 중
요한 것으로 관찰되었으며 이 부분이 TGF-β 자극에 의한 TIM-3 전사활성에도
중요한 것으로 관찰되었다. 또한 smad를 발현하는 plasmid를 이입한 세포의
luciferase reporter assay 결과 smad 2와 smad 4가 TIM-3 전사에 관여할 것이라고 예
측된다. 따라서 TIM-3의 전사에 -349 ~ +144 부분이 중요하게 작용하고 TGF-β에
의한 TIM-3 전사활성에 smad 2와 smad 4가 협동적으로 관여할 것으로 생각된다.
사람 혈액에서 분리한 단핵세포로부터 분화된 비만세포는 TGF-β 자극에 의
해 TIM-3의 발현이 증가된다는 보고가 있다. (Wiener 등, 2007). 앞의 보고와 동일
하게 비만 세포성 백혈병 환자에서 유래된 비만세포주인 HMC-1에서 역시 TGF-β
에 의한 TIM-3 mRNA의 발현증가를 관찰하였고, 이는 primary 세포와 마찬가지로
HMC-1에서도 TIM-3 발현증가에 TGF-β의 신호전달이 관여하는 것을 보여준다 (Fig. 1A). 사람 혈액에서 추출할 수 있는 비만세포의 수는 제한적이고 추출된 세
포의 배양에 있어서도 stem cell factor나 IL-6 같은 배양 인자가 충족되어야 하는 등 까다로운 실험조건을 요한다. 하지만 HMC-1는 배양이 훨씬 수월할 뿐만 아니 라 비만세포가 가지는 과립화 등과 같은 비만세포의 특성을 가지고 있어 비만세 포의 기능적 연구에 널리 사용되는 세포이다 (Butterfield 등, 1988). HMC-1 세포에 서 TGF-β에 의한 TIM-3 발현 증가가 관찰되었으므로 비만세포에서 TIM-3 발현 조절 기전을 밝힐 때 이 세포가 유용하게 이용될 수 있다고 생각되었다. 사람의 신장조직에서 TIM-3가 발현한다는 이전의 보고와 상응하게 (Ponciano
emd, 2006) 사람의 신장세포에서 유래된 HEK293 세포에서도 역시 TIM-3가 발현
하는 것을 확인하였다 (Fig. 1B). 그렇지만 TGF-β에 의해 TIM-3의 발현이 증가되
지 않았는데, 이것은 신장세포에서 비만세포와 TIM-3 발현조절이 다르게 일어남 을 보여준다.
34
의 상동성을 보인다 (Kuchroo 등, 2003). 한편 TIM-3 발현에 관여하리라 예측되는
upstream DNA 약 5 Kbp와 intron 염기서열은 사람과 생쥐간에 최고 87 %의 상동
성을 보이는 부분이 있는 반면 (intron 3 5625 ~ 5688 bp) intron 2와 intron 5에서는 상동성이 관찰되지 않았다.
본 연구에서는 TIM-3 upstream DNA와 intron 1과 intron 2가 TIM-3 발현조절에 어느 정도 관여하는가 luciferase reporter assay로 살펴보았으며, 3 전사에 TIM-3 upstream DNA의 -TIM-349 ~ +144 부분이 중요함을 밝혔다. 즉, TTIM-3U(1.8)-luc과
T3U(1.0)-luc의 luciferase activity는 T3U(0.5)-luc보다 상대적으로 낮게 관찰되었으며 (Fig. 3, Fig. 4), T3U(1.8)-luc에 비해 각각 1.9 Kbp, 1.8 Kbp가 더 긴 T3U(1.8/I1)-luc, T3U(1.8/I2)-luc의 luciferase activity가 T3U(1.8)-luc에 비해 더 높게 관찰되었다. 이
는 T3U(0.5)-luc의 luciferase activity가 가장 높은 이유가 T3U(0.5)-luc의 길이가 가 장 짧기 때문에 발생할 수 있는 실험적 오류일 가능성을 배제할 수 있게 하며, 대신 TIM-3 upstream DNA의 -349 ~ -1677 bp 사이에 TIM-3 발현억제에 관여하는 부분이 있을 가능성을 제기한다. 한편, TIM-3 upstream DNA -349 ~ +144 부분에서는
3 개의 smad binding site와 4 개의 GATA-1 binding site, 2 개의 AP-1 binding site, 2
개의 C/EBP binding site가 예측되었다. 따라서 이러한 전사인자가 TIM-3의 발현에 작용할 가능성이 있다고 생각된다.
TIM-3 upstream DNA와 intron이 같이 클로닝 된 vector의 luciferase activity는
HEK293 세포에서 intron이 단독으로 클로닝 된 vector의 luciferase activity 보다 높
았고 TIM-3 upstream DNA이 단독으로 클로닝 된 vector의 luciferase activity보다 높 았다 (Fig. 3). 이것은 HEK293 세포에서 basal level의 3 발현 시 intron이
TIM-3 promoter 활성을 증가할 가능성을 보여준다. 그렇지만 HMC-1의 luciferase reporter assay 결과는 다른 양상으로 관찰되었다. TIM-3 upstream DNA와 intron이 같
이 클로닝 된 vector의 luciferase activity는 intron이 단독으로 클로닝 된 vector의
luciferase activity 보다 높았지만 TIM-3 upstream DNA이 단독으로 클로닝 된 vector
의 luciferase activity와는 큰 차이가 없었다 (Fig. 4). 이것은 HMC-1의 basal level의
35
는 것이라고 생각된다. 그렇지만 luciferase reporter vector upstream DNA와 intron의 상대적 위치가 chromosomal DNA와 차이가 있으므로 HMC-1에서 이러한 결과가 나타났을 가능성을 배제할 수 없다.
비만세포에서 TGF-β 자극에 의해 다양한 길이의 TIM-3 upstream DNA를 포함
하는 vector의 luciferase activity와 TIM-3 upstream DNA의 -349 ~ +144 부분을 포함 하는 vector의 luciferase activity가 비슷한 수준으로 증가되고 또한 TIM-3 upstream
DNA의 -349 ~ +144 부분의 smad binding motif를 분석한 결과 TIM-3 upstream DNA
의 -215 ~ 204 bp, -120 ~ -106 bp, +108 ~ +139 bp의 3 군데에서 smad binding motif가
존재하기 때문에 TGF-β에 의한 TIM-3 발현증가는 TIM-3 upstream DNA의 -349 ~
+144 부분내의 smad binding motif를 통할 가능성이 클 것이라고 생각된다. 이것은
TGF-β에 의한 TIM-3 전사활성이 TIM-3 upstream DNA의 -349 ~ +144 내에서도 충
분할 것이라고 판단할 수 있다. 그렇지만 -349 ~ -319 부분은 인간 TIM-3와 생쥐 tim-3 간 상동성이 높은 반면, smad binding motif는 존재하지 않는 것으로 분석되 었기 때문에 -319 ~ +144 부분이 smad에 의한 신호전달에 관여할 것이라고 생각 된다. Smad에 대한 신호전달을 보다 확실히 알기 위해 TIM3 upstream DNA의
-349 ~ +144 부분의 smad binding motif에 대한 point mutation을 수행하여 luciferase activity를 비교해보는 실험이 뒤따라야 할 것이다. 한편 TGF-β 자극에 의해서 비
만세포에서 T3U(1.8/I1)-luc의 luciferase activity 또한 T3U(1.8)-luc의 luciferase
activity보다 약 1.8배 증가하는 것을 관찰하였다 (Data not shown). 이것은 비만세
포에서 TGF-β 자극에 의한 TIM-3 발현증가에 intron 1이 관여할 가능성을 제시한
다. 그렇지만 이 결과로 최소한 TIM-3 upstream DNA의 -349 ~ +144 부분이 TGF-β
에 의한 TIM-3 발현에 중요하게 작용한다는 것을 확인할 수 있다.
TGF-β에 의한 신호전달에 smad 분자가 관여한다는 논문이 보고 되어있다
(Lagna 등, 1996; Zhang 등, 1996). 또한 fibroblasts에 smad가 과발현 된 시스템에서
α-SMA promoter에 작용하는 smad의 영향을 확인한 논문을 근거로 판단해 볼 때
(Gu 등, 2007), HEK293 세포의 smad 과발현 시스템은 TGF-β에 의한 TIM-3 전사
36
TIM-3 발현 증가에 smad가 관여하는지 알아보기 위해서 smad가 과발현 된 HEK293 세포에서 luciferase activity를 측정한 결과, TGF-β에 의한 TIM-3 발현증가
에 smad 2와 4가 작용할 가능성이 있고 이러한 작용에는 smad 2와 4가 협동적으 로 관여할 것으로 생각된다 (Fig. 6). 이를 확인하기 위해서는 EMSA, ChIP assay를 통해서 smad가 TIM-3 regulatory DNA에 binding을 하는지에 대한 후속실험이 뒤따 라야 할 것이다.
결론적으로 본 연구에서는 HMC-1 세포에서 TIM-3 upstream DNA의 -349 ~
+144 부분이 basal level의 TIM-3 전사활성에 있어서 중요하게 작용할 것이라고
생각이 되고, TGF-β 자극에 의한 TIM-3 발현 증가에도 역시 -349 ~ +144 부분이
중요하게 작용을 하며 이러한 TGF-β에 의한 TIM-3 발현 증가에 smad 2와 smad 4
37
V. 결
결
결
결 론
론
론
론
TIM-3는 type I transmembrane protein으로서 비만세포에서 TGF-β에 의해서 발
현이 증가된다고 알려진 분자이다. 그렇지만 TGF-β에 의한 TIM-3의 분자생물학
적인 발현조절기작은 보고되어 있지 않다. 그렇기 때문에 본 연구는 TIM-3 발현 의 분자생물학적인 조절 기작을 밝히고자 TIM-3 upstream DNA와 intron 1, intron 2 를 각각 포함하는 TIM-3 luciferase reporter vector를 제작한 후 TIM-3 luciferase
reporter vector가 이입된 세포의 luciferase 활성을 측정하였다. 우선 비만 세포주인 HMC-1 세포와 신장 세포주인 HEK293 세포에서 TIM-3를 발현하고 HMC-1 세포
에서 TGF-β 자극에 의해서 TIM-3 mRNA가 증가하는 것을 확인하였다. Basal level
의 TIM-3 발현에 중요한 TIM-3 regulatory DNA 부분을 알아보고자 TIM-3 luciferase
reporter vector가 이입된 HMC-1 세포와 HEK293 세포의 luciferase 활성을 측정한
결과, TIM-3 upstream DNA의 -349 ~ +144 부분이 basal level의 전사활성에 있어서
HMC-1 세포와 HEK293 세포 둘 다에서 중요하게 작용할 것이라고 생각이 된다.
또한 intron 1과 intron 2가 HEK293 세포에서 basal level의 TIM-3 promoter 활성을
up-ragulation 할 것이라고 생각이 된다. TGF-β 자극에 의한 TIM-3 발현 증가에도
역시 -349 ~ +144 부분이 중요하게 작용을 하고 이러한 TGF-β에 의한 TIM-3 발현
38
참
참
참
참 고
고
고
고 문
문
문
문 헌
헌
헌
헌
1. Abdollah S, Macías-Silva M, Tsukazaki T, Hayashi H, Attisano L, Wrana JL: TbetaRI phosphorylation of Smad2 on Ser465 and Ser467 is required for Smad2-Smad4 complex formation and signaling. J Biol Chem. 272: 27678-27685, 1997
2. Anderson AC, Anderson DE: TIM-3 in autoimmunity. Curr Opin Immunol. 18: 665-669, 2006
3. Brück P, Ramos-Lopez E, Bartsch W, Böhme A, Badenhoop K: TIM-3 polymorphisms in type 1 diabetes families. J Hum Genet. 53:559-564, 2008
4. Butterfield JH, Weiler D, Dewald G, Gleich GJ: Establishment of an immature mast cell line from a patient with mast cell leukemia. Leuk Res. 12:345-55, 1988
5. Chae SC, Park YR, Lee YC, Lee JH, Chung HT: The association of TIM-3 gene polymorphism with atopic disease in Korean population. Hum Immunol. 65:1427-1431, 2004
6. Chae SC, Park YR, Shim SC, Yoon KS, Chung HT: The polymorphisms of Th1 cell surface gene Tim-3 are associated in a Korean population with rheumatoid arthritis.
Immunol Lett. 95:91-95, 2004
7. Chen D, Zhao M, Mundy GR: Bone morphogenetic proteins. Growth Factors. 4:233-241, 2004
8. de Caestecker M: The transforming growth factor-beta superfamily of receptors.
39
9. Dennler S, Huet S, Gauthier JM: A short amino-acid sequence in MH1 domain is responsible for functional differences between Smad2 and Smad3.Oncogene. 18:1643-1648, 1999
10. Derynck R, Zhang YE: Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature. 425: 577-584, 2003
11. Di Guglielmo GM, Le Roy C, Goodfellow AF, Wrana JL: Distinct endocytic pathways regulate TGF-beta receptor signalling and turnover. Nat Cell Biol. 5:410-421, 2003
12. Duan X, Liang YY, Feng XH, Lin X: Protein serine/threonine phosphatase PPM1A dephosphorylates Smad1 in the bone morphogenetic protein signaling pathway. J Biol
Chem. 281:36526-36532, 2006
13. Frisancho-Kiss S, Davis SE, Nyland JF, Frisancho JA, Cihakova D, Barrett MA, Rose NR, Fairweather D: Cutting edge: cross-regulation by TLR4 and T cell Ig mucin-3 determines sex differences in inflammatory heart disease. J Immunol. 178: 6710-6714, 2007
14. Feng XH, Derynck R: Specificity and versatility in tgf-beta signaling through Smads.
Annu Rev Cell Dev Biol. 21:659-693, 2005
15. Gebhardt T, Lorentz A, Detmer F, Trautwein C, Bektas H, Manns MP, Bischoff SC: Growth, phenotype, and function of human intestinal mast cells are tightly regulated by transforming growth factor beta1. Gut. 54: 928-934, 2005
16. Gomez G, Ramirez CD, Rivera J, Patel M, Norozian F, Wright HV, Kashyap MV, Barnstein BO, Fischer-Stenger K, Schwartz LB, Kepley CL, Ryan JJ: TGF-beta 1 inhibits mast cell Fc epsilon RI expression. J Immunol. 174: 5987-5993, 2005
40
17. Graff JM, Bansal A, Melton DA: Xenopus Mad proteins transduce distinct subsets of signals for the TGF beta superfamily. Cell. 85:479-487, 1996
18. Hanai J, Chen LF, Kanno T, Ohtani-Fujita N, Kim WY, Guo WH, Imamura T, Ishidou Y, Fukuchi M, Shi MJ, Stavnezer J, Kawabata M, Miyazono K, Ito Y: Interaction and functional cooperation of PEBP2/CBF with Smads. Synergistic induction of the immunoglobulin germline Calpha promoter. J Biol Chem. 274:31577-31582, 1999
19. Hocevar BA, Smine A, Xu XX, Howe PH: The adaptor molecule Disabled-2 links the transforming growth factor beta receptors to the Smad pathway. EMBO J. 20:2789-2801, 2001
20. Hoodless PA, Haerry T, Abdollah S, Stapleton M, O'Connor MB, Attisano L, Wrana JL: MADR1, a MAD-related protein that functions in BMP2 signaling pathways. Cell. 85:489-500, 1996
21. Inman GJ, Hill CS: Stoichiometry of active smad-transcription factor complexes on DNA. J Biol Chem. 277:51008-51016, 2002
22. Kearley J, McMillan SJ, Lloyd CM: Th2-driven, allergen-induced airway inflammation is reduced after treatment with anti-Tim-3 antibody in vivo. J Exp Med. 204: 1289-1294, 2007
23. Knockaert M, Sapkota G, Alarcón C, Massagué J, Brivanlou AH: Unique players in the BMP pathway: small C-terminal domain phosphatases dephosphorylate Smad1 to attenuate BMP signaling. Proc Natl Acad Sci U S A. 103:11940-11945, 2006
24. Koguchi K, Anderson DE, Yang L, O'Connor KC, Kuchroo VK, Hafler DA: Dysregulated T cell expression of TIM3 in multiple sclerosis. J Exp Med. 203: 1413-1418, 2006
41
25. Kuchroo VK, Umetsu DT, DeKruyff RH, Freeman GJ: The TIM gene family: emerging roles in immunity and disease. Nat Rev Immunol. 3: 454-462, 2003
26. Lagna G, Hata A, Hemmati-Brivanlou A, Massagué J: Partnership between DPC4 and SMAD proteins in TGF-beta signalling pathways. Nature. 383:832-836, 1996
27. Lehmann K, Janda E, Pierreux CE, Rytömaa M, Schulze A, McMahon M, Hill CS, Beug H, Downward J: Raf induces TGFbeta production while blocking its apoptotic but not invasive responses: a mechanism leading to increased malignancy in epithelial cells.
Genes Dev. 14:2610-2622, 2000
28. Lin X, Duan X, Liang YY, Su Y, Wrighton KH, Long J, Hu M, Davis CM, Wang J, Brunicardi FC, Shi Y, Chen YG, Meng A, Feng XH: PPM1A functions as a Smad phosphatase to terminate TGFbeta signaling. Cell. 125:915-928, 2006
29. Liu F, Hata A, Baker JC, Doody J, Cárcamo J, Harland RM, Massagué J: A human Mad protein acting as a BMP-regulated transcriptional activator. Nature. 381:620-623, 1996
30. Massagué J: How cells read TGF-beta signals. Nat Rev Mol Cell Biol. 1: 169-178, 2000
31. Massagué J: TGF-beta signal transduction. Annu Rev Biochem. 67:753-791, 1998
32. Matsushita N, Nishi N, Seki M, Matsumoto R, Kuwabara I, Liu FT, Hata Y, Nakamura T, Hirashima M: Requirement of divalent galactoside-binding activity of ecalectin/galectin-9 for eosinophil chemoattraction. J Biol Chem, 275: 8355-8360, 2000
33. Miller HR, Wright SH, Knight PA, Thornton EM: A novel function for transforming growth factor-beta1: upregulation of the expression and the IgE-independent extracellular release of a mucosal mast cell granule-specific beta-chymase, mouse mast cell protease-1. Blood. 93: 3473-3486, 1999
42
34. Monney L, Sabatos CA, Gaglia JL, Ryu A, Waldner H, Chernova T, Manning S, Greenfield EA, Coyle AJ, Sobel RA, Freeman GJ, Kuchroo VK: Th1-specific cell surface protein Tim-3 regulates macrophage activation and severity of an autoimmune disease. Nature. 415: 536-541, 2002
35. Moustakas A, Kardassis D: Regulation of the human p21/WAF1/Cip1 promoter in hepatic cells by functional interactions between Sp1 and Smad family members. Proc
Natl Acad Sci U S A. 95:6733-6738, 1998
36. Nakae S, Iikura M, Suto H, Akiba H, Umetsu DT, Dekruyff RH, Saito H, Galli SJ: TIM-1 and TIM-3 enhancement of Th2 cytokine production by mast cells. Blood. 110: 2565-2568, 2007
37. Nicolás FJ, De Bosscher K, Schmierer B, Hill CS: Analysis of Smad nucleocytoplasmic shuttling in living cells. J Cell Sci. 117:4113-4125, 2004
38. Norozian F, Kashyap M, Ramirez CD, Patel N, Kepley CL, Barnstein BO, Ryan JJ: TGFbeta1 induces mast cell apoptosis. Exp Hematol. 34: 579-587, 2006
39. Oikawa T, Kamimura Y, Akiba H, Yagita H, Okumura K, Takahashi H, Zeniya M, Tajiri H, Azuma M: Preferential involvement of Tim-3 in the regulation of hepatic CD8+ T cells in murine acute graft-versus-host disease. J Immunol. 177: 4281-4287, 2006
40. Olsson N, Rak S, Nilsson G: Demonstration of mast cell chemotactic activity in bronchoalveolar lavage fluid collected from asthmatic patients before and during pollen season. J Allergy Clin Immunol. 105: 455-461, 2000
43
41. Ponciano VC, Renesto PG, Nogueira E, Rangel EB, Cenedeze MA, Franco MF, Câmara NO, Pacheco-Silva A: Tim-3 expression in human kidney allografts. Transpl
Immunol. 17:215-222, 2007
42. Sabatos CA, Chakravarti S, Cha E, Schubart A, Sánchez-Fueyo A, Zheng XX, Coyle AJ, Strom TB, Freeman GJ, Kuchroo VK: Interaction of Tim-3 and Tim-3 ligand regulates T helper type 1 responses and induction of peripheral tolerance. Nat Immunol. 4: 1102-1110, 2003
43. Sánchez-Fueyo A, Tian J, Picarella D, Domenig C, Zheng XX, Sabatos CA, Manlongat N, Bender O, Kamradt T, Kuchroo VK, Gutiérrez-Ramos JC, Coyle AJ, Strom TB: Tim-3 inhibits T helper type 1-mediated auto- and alloimmune responses and promotes immunological tolerance. Nat Immunol. 4: 1093-1101, 2003
44. Schmierer B, Hill CS: Kinetic analysis of Smad nucleocytoplasmic shuttling reveals a mechanism for transforming growth factor beta-dependent nuclear accumulation of Smads. Mol Cell Biol. 25:9845-9858, 2005
45. Shi Y, Massagué J: Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. 113: 685-700, 2003
46. Sui L, Zhang W, Chen Y, Zheng Y, Wan T, Zhang W, Yang Y, Fang G, Mao J, Cao X: Human membrane protein Tim-3 facilitates hepatitis A virus entry into target cells. Int J
Mol Med. 17: 1093-1099, 2006
47. Tsukazaki T, Chiang TA, Davison AF, Attisano L, Wrana JL: SARA, a FYVE domain protein that recruits Smad2 to the TGFbeta receptor. Cell. 95:779-791, 1998
48. Watanabe M, Masuyama N, Fukuda M, Nishida E: Regulation of intracellular dynamics of Smad4 by its leucine-rich nuclear export signal. EMBO Rep. 1:176-182, 2000