INTRODUCTION
It is a well-known fact that single cells among both survived and inactivated parts of a homogeneous popula-tion respond differently to the same dose of irradiapopula-tion. Inactivated cells produce microcolonies consisting of various numbers of cells (different inactivation forms) (Korogodin 1958). Survived cells produce on solid nutrient media colonies of various sizes and morphology which appear within different time interval after irradiation. It agrees with the modern theory of microdosimetry and classical hit-and-target models of radiobiology. The late
appearance of clones produced by cell survived after ioniz-ing radiation is expressed only for diploid and polyploid cells for which sigmoid survival curves after exposure by ionizing radiation are obtained. Each lesion is lethal only for haploid cells. In diploid cells, effects of several primary lesions (sublethal lesions) may be summarized and inherited resulting in various mitotic disorders. These disorder effects are the higher the greater number of primary sublethal lesions is produced in the originally irradiated cells. In the course of producing macrocolonies by the irradiated diploid cells, � lethal sectors� are often formed (Korogodin et al. 1968; Korogodin 1993).
It was shown before that different manifestation of cell radiation damage can be quantitatively described by the probability model (Kapultsevich and Petin 1977;
Kapult-─ ─ 59 ──
Yeast Cells Retain a Memory of Their Original
Radiation-Induced Insults
Vladislav G. Petin, Olga V. Pereklad, Mohammad Nili1and Jin Kyu Kim2,*
Medical Radiological Research Center, 249036 Obninsk, Kaluga Region, Russia 1Dawnesh Radiation Research Institute, 08007 Barcelona, Spain 2Korea Atomic Energy Research Institute, Jeongeup 580-185, Korea
Abstract -- In accordance with microdosimetry idea and classical hit-and-target models of radiobiology, various cells exposed to the same dose are damaged in a different manner in accor-dance with the hit-principle. It means that the survived cells can differ in the content of sublesions (hits) produced by the energy absorbed inside the cell and which is not enough to create the effec-tive damage responsible for cell inactivation. Taking into considerations these ideas, the growth rate of cells from 250 colonies of diploid yeast cells of various sizes which appeared in different time after irradiation (60Co, 600 Gy, survived 20%) was determined. Based on these results, the dis-tribution of clones grown from irradiated cells according to the number of sublesions in the origi-nal cell survived after irradiation was depicted. It allowed us to select cells with 0, 1, 2, 3, and 4 sublesions. Clones with various numbers of sublesions were shown to differ in their viability, radio-sensitivity, sensitivity to environmental conditions, frequency of recombination and respiratory-deficient mutations. It is concluded that the application of genetic and biochemical methods for identification of the nature of these sublesions will promote elucidation of the nature of effective primary radiation damage finally resulting in cell inactivation.
Key words: Ionizing radiation, Sublethal lesion, Probability model, Yeast cells
* Corresponding author: Jin Kyu Kim, Tel. +82-63-570-3130, Fax. +82-63-570-3139, E-mail. [email protected]
sevich 1978). The model is based on the supposition that clone formation is a probable process and the probability for the successful division of cell is determined by a num-ber of damages. This means that the probability model is a peculiar synthesis of hit-and-target principle and biological stochastic. The model qualitatively explains a wide variety of cell responses to irradiation and quantitatively describes survival curves and the yield of inactivation forms of diffe-rent yeast strains (Kapultsevich and Petin 1977; Kapult-sevich 1978). According to this model, single irradiated cells are damaged in a random fashion in accordance with the hit-principle, as in classical models (Timofeeff-Ressov-sky 1972; Alper 1979), and the number of damages defines the probability P for the successful division. If the proba-bility of damage expression (the probaproba-bility of refusal) is α,
the probability for the successful division of a cell with one damage is
P1==(1-α). (1)
For independent interaction of radiation damages, the probability for the successful division of a cell with i pri-mary damage (sublethal lesion, hit) may be presented as
Pi==(1-α)i. (2)
It was demonstrated that the reduced probability for the successful division can remain unchanged through the succes-sive division of cells (Kapultsevich and Petin 1977; Kapultse-vich 1978). Hence, the mechanism of clone formation is the well-known “birth-and-death” process. The process operates when an entity, in our case a single cell, either gives rise to progeny like itself (birth), or is removed in some way (death), and these two events occur in a random fashion. Taking into account this approach, it would be of interest to obtain ex-perimentally the distribution of survived cells in accordance with the number of primary lesions and after that, taking the population of cells with a known number of sublethal lesions, to determine various biological responses in the dependence of the primary sublethal lesions. These tasks were performed in this study.
MATERIALS AND METHODS
The following diploid yeast strains were used in this study-Saccharomyces ellipsoideus (vini), strain Megri
139-B, and Saccharomyces cerevisiae, strain 5a3bα that is het-erozygous for the ade2 mutation. Yeast cells were incuba-ted before irradiation for 3~5 days at 30�C on a complete nutrient agar layer to a stationary phase. Aliquots with 106 yeast cells ml-1 were exposed to ionizing radiation. We used a 60Co ray source (Gammacell 220, AECL). The γ-ray dose-rate, estimated by a Siemens ionization chamber, was 20 Gy min-1. The survival was assessed by counting colonies in platings. Primary colonies, which appear after irradiation, were used to obtain subclones. Colonies, sub-clones of which grew simultaneously with a control and did not differ from the control phenotypically, were identified as stable (normal cell clones). Colonies, which formed slowly growing subclones and/or subclones that differed in morphology, were believed to be unstable clones.
To determine the content of mitotic recombinants in diplo-ids that are heterozygous for the ade1 and ade2 mutations, colonies were replated on nutrient media YEPD and the number of white, red and sectored colonies was counted. When estimating culture growth rate, cells of separate clones were grown in liquid nutrient media in tubes under aeration; the number of cells, capable of forming colonies was deter-mined in plating. The growth rate was calculated according to the exponential region of growing curve. The quantitative evaluation of respiratory mutants was performed as was published by others (Ogur et al. 1957), and the content of nonviable cells was determined by detecting, under a micro-scope, budding and nonbudding cells on the surface of the nutrient agar after growing for one day. Other details have been published (Kim et al. 2001; Petin and Kim 2004).
EXPERIMENTAL RESULTS
AND DISCUSSION
We determined the growth rate of cells from colonies of various sizes appearing in different time after irradiation. Fig. 1 show the growth of control cells in liquid nutrient medium at 30�C (curves 1). Curves 2~4 represent growth curves of yeast cells from individual colonies, produced by cells survived after irradiation and having 1~3 primary damage (sublethal lesion, hit) respectively. These curves have much lower slopes (i.e., slower growth rate). The growth rate of cells from the same colony was retained unchanged during a lot of passages (Ka pultsevich and Petin
1977; Kapultsevich 1978).
It was shown before (Kapultsevich and Petin 1977) that for diploid strains Megry-139B and 5a3Bα the probability of refusal α==0.12. Hence, the following probabilities for
the successful division of cells with 0, 1, 2, 3 and 4 dama-ges can be obtained: Po==1, P1==0.88, P2==0.77, P3==0.68
and P4==0.60. It turned out that the reduced probability for
the successful division of damaged cell is retained for a large number of cell generation (Kapultsevich and Petin 1977; Kapultsevich 1978). It leads to the decrease of clone-formation rate immediately after irradiation and upon the successive plating.
If the reduction of growth rate is related with the decrea-sing of the probability Pifor the successful division, the
specific growth rate of cells μiwith i sulethal lesions may
be given as
μi==μc(2Pi-1), (3)
where μc is the specific growth rate of control cells. We
determined the specific growth rate of cells from 150 (Megry-139B) and 100 (5A×3B) colonies produced by the
survived irradiated diploid yeast cells (60Co, 600 Gy,
survi-ved ≈20%).
Knowing μiand μc, we calculated Pifor these clones.
Experimental results are presented in Fig. 2. It is of interest that the maxima of the distribution almost coincide with
theoretical values of Pipredicted for cells with 0, 1, 2, and
3 primary damage (sublethal lesion). It would be of interest to test whether clones with various values of damages differ in their radiosensitivity, viability, sensitivity to environmen-tal conditions, frequency of recombination and respiratory-deficient mutations.
When colonies of S. cerevisiae haploid and diploid strains,
0 10 20 30 40 50 60 70 0 10 20 30 40 50 60 70 Time, hours 108 107 106 105 104 103 102 Concentration, cells ml -1 (A) (B) 1 2 3 4 1 2 3 4
Fig. 1. Dynamics of the propagation of nonirradiated cells (curves 1) and distant progenies of cells survived after irradiation and generating unstable colonies (curves 2~4); strains: Megry 139-B (panel A) and 5a3bα (panel B).
40 30 20 10 0 Number of events 1 2 3 4 1 2 3 4 0
Number of sublethal lesions in the cells
(A) (B)
1.0 0.9 0.8 0.7 0.6 1.0 0.9 0.8 0.7 0.6 Probability of cell division
Fig. 2. Distribution of clones grown from irradiated cells accor-ding to the probability for successful division (upper scale of the abscissa), corresponding to the number of primary dam-age (sublethal lesion) in the original cell (lower scale of the abscissa); strains: Megry 139-B (panel A) and 5a3bα (panel B).
which were obtained after a single irradiation, were irradiat-ed repeatirradiat-edly, Tobias was the first (Tobias 1952) to state that radiosensitivity of repeatedly irradiated haploids does not differ from that of the original cultures. However, in diploids, cells of some colonies that appeared after irra-diation showed an increased radiosensitivity. These results were interpreted by mitotic inheritance of sublethal lesions occurring in diploid cells. No morphological differences between these colonies or any sign of instability were men-tioned in his work (Alper 1979). It was therefore of interest to know whether there was a difference between radiosen-sitivity of cells from normal clones and from that contain-ing the various number of sublethal lesions. Fig. 3 exhibits the results obtained. It can be seen that cells from unstable clones containing various number of primary sublethal le-sions (curves 2~5) have an increased radiosensitivity
com-pared to control cells (curves 1). This is expressed as a dec-rease in the survival curve shoulder and in an incdec-rease of its slope. In formal terms, the shoulder decrease means that cells initially had a number of primary sublethal lesions. The slope increase can indicate that the expression of the postradiation recovery process is somewhat weaker in these cells than in nonirradiated cells or cells from normal clones. As can be seen from Fig. 3, the radiosensitivity of distant
progenies of single cell, survived after irradiation, corre-lates with the number of primary sublethal lesions. Effects of separate lesions are summarized. Cells with a great num-ber of primary sublethal lesions are inactivated with a great probability while those with a lesser number of sublesions can give rise to clones differing in both growth rate and radiosensitivity, the degree of radiosensitivity being the higher the greater number of sublesions are in the original cell survived after irradiation.
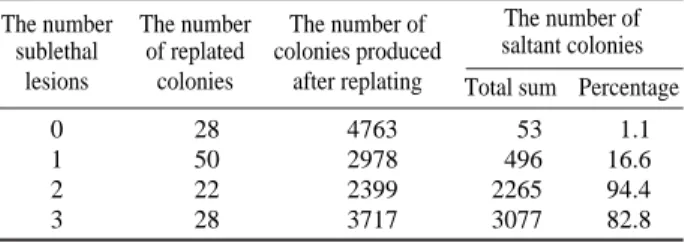
The appearance among macrocolonies produced by irradi-ated diploid yeast cells with various morphological changes may be attributed to the example of the expression of pri-mary sublethal lesions. The content of such kind of cells in colonies produced by survived diploid yeast cells (strain Megry 139-B, 60Co γ-rays, 600 Gy) is summarized in Table
1. One can see that the content of morphologically changed cells with two and three sublethal lesions is significantly greater compared to that for clones without or with one pri-mary sublethal lesion (hit).
The increased probability of unsuccessful division (re-fusal) should result in the existence of nonviable cells in clones produced by single cells survived after irradiation. Then there is a reasonable base to expect the increased number of nonviable cells in unstable clones. Experimental
100 10 1 Survival, % 0 200 400 600 800 0 200 400 600 800 1000 (A) (B) 3 5 4 3 2 1 2 1 Dose, Gy
Fig. 3. Survival curves of control cells (curves 1) and cells from unstable clones containing various number of primary sublethal lesions (curves 2~5); strains: Megry 139-B, i==0, 1, 2 for curves 1, 2, 3, respectively (panel A) and 5a3bα, i==0, 1, 2, 3, and 4 for curves 1, 2, 3, 4, and 5, respectively (panel B).
data concerning the content of cells incapable to prolifera-tion at optimal (30�C, standard nutrient media) and subopti-mal (37�C, standard nutrient media++7% NaCl) conditions
for distant progenies of diploid yeast cells (strain Megry 139-B, 600 Gy) with various number of primary sublethal radiation lesions are presented in Table 2.
It could be easily seen from the results presented in Table 2 that the relative yield of nonviable cells in clones produc-ed by survivproduc-ed diploid yeast cells is increasproduc-ed with the num-ber of inherited primary radiation sublethal lesions. Due to this fact, the growth effectiveness of cells under suboptimal conditions of culture was 76, 46 and 13% correspondingly to clones with 0, 2, and 3 primary sublethal lesions.
Some unstable clones differ by a high segregation rate of respiratory mutants. Although respiratory mutants are vir-tually not encountered among primary colonies of irra-diated diploid, these mutants can constitute more than 30% of all subclones in plating of some unstable clones. The data concerning the yield of respiratory mutants in clones with various numbers of primary sublethal lesions are pre-sented in Table 3. These experiments were performed with diploid yeast cells (strain Megry 139-B) which were irra-diated in the stationary phase of growth (60Co γ-rays, 600
Gy). To identify respiration deficiency in yeast we used the tetrazolium overlag technique. According to this method, cells with normal respiratory ability were colored in red
color while clones consisting of respiratory mutants stayed white. One can see that irradiation resulted in an increased content of respiratory mutants in clones produced by irradi-ated cells. The effect was particular expressed for clones with a greater number of primary sublethal lesions (hits).
CONCLUSIONS
It was shown in this paper that unstable clones were for-med among colonies grown from diploid yeast cells sur-vived after irradiation. Cells from unstable clones exhibited an increased (as compared with the control) content of mor-phologically changed cells, nonviable cells and respiration mutants. Cells from unstable clones are characterized by the enhanced radiosensitivity. The degree of expression of the foregoing effects was the higher the greater number of primary sublethal lesions was in the originally irradiated cell. The data obtained in this paper can be explained by the hypothesis in accordance with which the primary radiation lesion is not absolutely lethal for the diploid cell. With some probability which depends on the total number of primary sublethal lesions and conditions of culture, they result in the disturbance of cell division. Such a disturbance revealed by cell inactivation (nonviable cells among clones produced by irradiated cells), incorrect distribution of mitochondria between daughter cells (respiratory mutants). Thus, the suggested methodology of identification of cells with a definite number of primary sublethal lesions will promote further elucidation of the nature of primary radia-tion damage.
ACKNOWLEDGEMENTS
This work was carried out under the National R & D
Pro-Table 1. The content of morphologically changed cells among colonies produced by irradiated diploid yeast cells The number The number The number of The number of
sublethal of replated colonies produced saltant colonies lesions colonies after replating Total sum Percentage
0 28 4763 53 1.1
1 50 2978 496 16.6
2 22 2399 2265 94.4
3 28 3717 3077 82.8
Table 3. The content of respiratory mutants in clones produced by survived diploid yeast cells exposed to ionizing radiation The number of The percentage of clones containing primary radiation various number of respiratory mutants (%)
sub lethal lesions 0 ¤2 ¤10 ¤50
(hits) ~2% ~10% ~50% ~100%
0 100 - -
-1 80 16 2 2
2 53 12 20 15
3 32 16 28 14
Table 2. The content of nonviable cells in clones produced by di-ploid yeast cells survived after exposure to ionizing radi-ation
The content of
The number of The number nonviable cells, %
primary sublethal of tested
Optimal Suboptimal
lesions (hits) clones
condition condition
0 28 17±6 24±7
1 50 20±5 43±11
2 22 27±9 54±15
gram by the Ministry of Education, Science and Techno-logy of Korea. It was also supported by the International Cooperation Program and the Russian Fund of Humanity Sciences and Administration of Kaluga Region (Grant No. 08-06-59609).
REFERENCES
Korogodin VI. 1958. Inactivation forms of yeast cells after exposure to ionizing radiation, Biophysica 3:206-214. Korogodin VI, Kapultsevich YG, Myasnik MN, Mosin AF and
Gridnev VV. 1968. Cellular repair processes: survival of irra-diated yeast, bacteria, and phages under different postra-diation conditions, Adv. Biol. Med. Physics 12:253-274. Korogodin VI. 1993. The study of post-irradiation recovery of
yeast: the premolecular period, Mutat. Res. 289:17-26. Kapultsevich YG and Petin VG. 1977. Probability model for
cell responses to irradiation, Studia Biophysica 62:151-165.
Kapultsevich YG. 1978. Quantitative Regularities of Cell Radia-tion Lesions, Moscow: Atomizdat.
Timofeeff-Ressovsky NV, Ivanov VI and Korogodin VI. 1972. Die Anwendung des Trefferprinzip in der Strahlengbio-logie, Jena, VEB Gustav Fischer Verlag.
Alper T. 1979. Cellular Radiobiology. Cambridge: University Press.
Ogur H, John R and Nagai S. 1957. Tetrazolium overlag techni-que for population studies of respiration deficiency in yeast, Science 125:928-931.
Kim JK, Petin VG and Zhurakovskaya GP. 2001. Exposure rate as a determinant of synergistic interaction of heat com-bined with ionizing or ultraviolet radiations in cell killing, J. Radiat. Res. 42:361-365.
Petin VG and Kim JK. 2004. Survival and recovery of yeast cells after combined treatments with ionizing radiation and heat, Radiat. Res. 161:56-63.
Tobias CA. 1952. The dependence of some biological effects of radiation on the rate of energy loss, pp. 97-113, In: Sym-posium on Radiobiology. The Basic Aspects of Radiation Effects on Living Systems (Nickson JJ ed.), Oberlin Colle-ge, NY, John Wiley and Sons.
Manuscript Received: April 11, 2008 Revision Accepted: May 14, 2008