INTRODUCTION
When plant cells are exposed to stresses, such as drought, salinity and cold, a proline accumulation is a widespread phenomenon. Proline increase plays a key role for an osmo-tic adjustment in a large number of plant species under stress conditions (Delauney and Verma 1993). Mutants resistant to proline analogs are thought to be a useful material for studying a proline biosynthesis and a salinity, drought, and cold resistance in higher plants (Miflin et al. 1983). Azetidine-2-carboxylic acid (AZCA), a proline analog, is a natural
product found in some species of Liliaceae, which inhibited an irreversible cell growth by competition with proline for incorporation into protein (Cella et al. 1982; Hyun et al. 2003). This incorporation presumably leads to an altered protein conformation and function, and accounts for the cytotoxic effects of AZCA (Lodato et al. 1984; Song et al. 2007). In many cases, the mechanism of a resistance to amino acid analogs has been shown to be due to the insensitivity to a feedback inhibition (Widholm 1974). Mutants with an altered feedback mechanism express more resistant to a feedback inhibition and these result in a greatly elevated free proline accumulation. The decrease of uptake inhibitors by a change in cell membrane permeability and an inactivation of inhibi-tors have also been reported as mechanisms of an amino acid analog resistance.
─ ─ 179 ──
Selection of Gamma-ray Induced Salt Tolerant Rice
Mutants by in vitro Mutagenesis
Dong Sub Kim†, Jae Young Song1,†, Jaebeom Chun, Kyung Jun Lee, Jin-Baek Kim, Sang Hoon Kim, Song Joong Yun1and Si-Yong Kang*
Advanced Radiation Technology Institute, Korea Atomic Energy Research Institute, Jeongeup 580-185, Korea
1Department of College of Crop Agriculture & Life Science Agriculture & Life Sciences, Chonbuk University, Jeonju 561-756, Korea
Abstract -- The present study has been performed to select the salt tolerant rice mutant lines through an in vivo and in vitro mutagenesis with a gamma-ray. The physiological responses such as MDA and chlorophyll of the selected salt mutant lines were investigated under salt stress. For the selection of the salt tolerant rice mutants by in vitro mutagenesis with gamma-ray, we conducted a second selection procedure with 1,500 mutant lines induced from the original cv. Dongan (wild-type, WT): 1st, selection under a nutrient solution with 171 mM NaCl; 2nd, selection under in vitro conditions. Based on a growth comparison of the entries, out of mutant lines, the putative 2 salt tolerant rice mutant lines, 495 and 532, were selected. The 2 ST-lines had a lower malonaldehyde (MDA) contents than wild-type (WT) during salt stress. The survival rate of the WT, ST-495 and ST-532 were 36.6%, 70% and 50% in 171 mM NaCl, respec-tively. The chlorophyll and carotenoid contents were decreased more in a WT plant than the two selected mutant lines. These rice mutant lines will be released for cultivation at the reclaimed land and used as a control plot for genetic research about salt tolerance.
Key words : Gamma-ray, Mutant, Rice, Salt tolerance
†Kim DS and Song JY contributed equally to this work. * Corresponding authors: Si-Yong Kang, Tel. +82-63-570-3310,
Radiation induced mutagenesis has been studied extensive-ly for the identification and isolation of a mutated plant gene in plant breeding programs. Rapid progress in plant tissue culture methods is attributed to the availability of many mutants that can be generated for specific biochemical events that are difficult or impossible to use with intact plants (Carl-son 1970; Larkin and Scowcroft 1981; Schaeffer and Sharpe 1983; Kim et al. 2004). This may be achieved by co-cultur-ing plant tissues with pathogens or their toxins and by mani-pulating a medium composition, e.g. selection for a tolerance to salinity, drought and amino acid analogs. Although, it is possible to select mutants with a salt tolerance by a somaclo-nal variation, mutant frequencies are often lower in plant systems due to inefficient recovery techniques, as well as the presence of multiple gene copies located on different chromosomes, and by cell-to-cell interactions in even the most ideal suspension culture systems (Schaffer and Sharpe 1983). Therefore, in vitro mutation induction techniques pro-vide tools for a rapid creation and increase of the selection frequencies of cell types for which there is no obvious coun-terpart in field environments (Maluszynski et al. 1995). Such techniques enable a greater use of mutated genes for a crop improvement, the cumulative effect of which may result in a more desirable character than the original variety. Rice, an important staple food, is consumed by more than half of the world’s population. It is sensitive to a salt stress, therefore, development of varieties with an increased salt tolerance is required. The possibility of a screening for a salt stress in a culture makes in vitro methods attractive for developing stress tolerant plants (Lee et al. 2003). We have constructed AZCA resistant rice mutant population with altered proline feedback inhibition. This mutant population was selected via in vitro mutagenesis with AZCA, regeneration, and a
consecutive selection through each generation (M2~M3). In
the present study, in order to develop of salt tolerant rice variety, we described the selection procedures for the selec-tion of salt tolerant lines and physiological characterizaselec-tion of the selected mutant lines under salt stress.
MATERIALS AND METHODS
1. Plant materialsFor increasing the contents of proline in rice (Oryza sativa L.) cv. Dongan, the mutant cell lines resistant to growth
inhi-bition by Azetidine-2-carboxylic acid (AZCA) were selected from the callus irradiated with a gamma ray of 50, 70 Gy through embryo culture. The resistant mutant lines (M2~
M3) to AZCA were obtained from the consecutive selection.
These lines showed high level of free proline contents (Hyun et al. 2003). By a successive selfyng, we obtained M6rice
population including 1,500 mutant lines. This population used for the selection of salt tolerant mutants and physiologi-cal analysis.
2. In vivo screening of salt tolerant rice mutants To develop rice varieties to be used for saline paddy sites, the seeds of 1,500 mutant lines (M6generation) and the
origi-nal cv. Dongan (wild-type, WT) were planted in trays with 15 seeds per line were placed in each pot at a depth of 5 mm. The trays were filled with fine soil, which was commercially made for rice culture. The trays were watered with tap water until the 3 to 4 leaf stage. At that stage, excess water was drained, and the trays with rice seedlings were refilled with solution containing 171 mM (1%, E.C==16 mS) salt and 1 g l-1 fertilizer. The solution was circulated with an underwater rotator to maintain a uniform salt concentration for 7 days in a glasshouse. The E.C measurements were taken daily. After salinization with 171 mM, salinity symptoms were scored according to the Standard Evaluation System (1~3: tolerant, 5: moderate and 7~9: sensitive) developed at IRRI (International Rice Research Institute) (data not shown). We have investigated survival rate of the mutant lines and WT. To re-select the salt tolerance, each 30 seeds of the selected mutant lines (M7) and WT were germinated and the seedlings
were grown in soil trays (without salt) for 14 days. After 7 days salt treatment, for the investigation of the recovery of salt tolerant mutant lines, seedlings were transferred to tap water for 7 days under a growth condition (16 h light/8 h dark); the water was exchanged every day during this period.
3. In vitro screening of salt tolerant rice mutants The seeds (15 seeds per line) of the WT and mutant lines selected through in vivo screening were sterilized in 75% ethanol for 30 sec and followed with 5% sodium hypochlo-rite for 30 min. The seeds were rinsed 3~4 times with sterile distilled water and transferred to selection medium contain-ing the 1/2 MS medium, 171 mM (1.0%) NaCl and 15 g l-1 sucrose solidified with 8% agar powder in the glass bottle.
The seeds were germinated and cultured in a growth
cham-ber at 27±1�C under growth condition. The WT and mutant
lines were cultivated in three replications, respectively. Final-ly, two salt tolerant (ST) lines, ST-495 and ST-532, were selected after 40 days.
4. Measurement of lipid peroxidation
The WT and 2 mutant lines were imbibed in 171 mM NaCl solution at 28±1�C for 7 days at the seedling stage. After treatment, the shoots harvested at the indicated time, frozen in liquid nitrogen and stored at -80�C until physiological analysis. Lipid peroxidation in the fresh leaf tissue was deter-mined in terms of malondialdehyde (MDA) contents by trichloroacetic acid (TCA) reaction as described method by Dhindsa (1981) and Ding (2008). Fresh leaf (0.5 g) was ground with 5 ml of 0.1% TCA. The homogenate was centrifuged at 13,000 rpm for 10 min and the supernatant transferred into a new tube. The supernatant of 1 ml was mixed with 4 ml of 20% TCA buffer containing 0.5 thiobarbituric acid (TBA). The mixture was heated at 100�C for 30 min, immediately cooled on ice. The cooled mixture was centrifuged at 13,000 rpm for 10 min. Malondialdehyde (MDA) content was mea-sured spectrophotometrically at A532and corrected for
non-specific turbidity at A600. The concentration of MDA is
expressed as nmol MDA per gram fresh weight.
5. Chlorophyll and carotenoid measurements To determine total chlorophyll content, leaves of each
plant were weighed and frozen at -80�C 5 ml of extraction buffer, acetone (100%), was added to frozen tissues in a 15 ml tube and placed at 4�C in darkness for 24 hours. The samples were centrifuged at 10,000 rpm at 4�C for 10 min. The supernatant was transferred to new microcentrifuge tube. The absorption spectrum at 470, 644.8 and 661.6 nm of the extracted liquid (1 ml) in a quartz cuvette using 100% ace-tone as a blank was measured with a spectrophotometer (UVIKON 923, Bio-Tek Instruments, Winooski, Vermont, USA). Chlorophyll content was calculated as follows with using extinction coefficients reported by Lichtenthaler (1987).
RESULTS AND DISCUSSION
1. Screening of rice mutant lines under salt stress To screen the salt tolerance at the seedling stage, a total of 1,500 mutant lines (M6) were used and then applied undera 171 mM salt solution. The preliminary screening on the basis of seedlings survival to salt stress showed that 56 out of 1,500 mutant lines evidenced tolerance under the salt level.
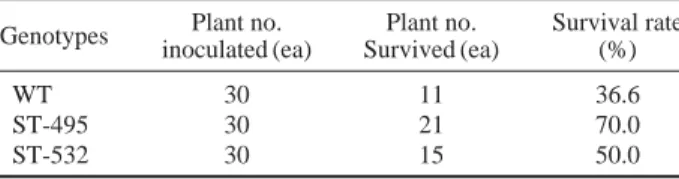
Table 1. Survival rate of the WT and 2 ST-lines after 14 days under
171 mM salt solution
Genotypes Plant no. Plant no. Survival rate inoculated (ea) Survived (ea) (%)
WT 30 11 36.6
ST-495 30 21 70.0 ST-532 30 15 50.0
Fig. 1. Comparison of the WT and 2 selected mutant lines. Seeds of control and 2 mutants were germinated and grown in soil trays (without
salt) for 14 days and then irrigated with 171 mM NaCl solutions for 7 days (A) and 7 days after recovery in without salt solution (B). (A) (B) 35.0 30.0 25.0 20.0 15.0 10.0 5.0 0.0 Shoot length (cm) WT ST-495 ST-532
7 days after salt-stress 7 days after recovery
The salt stress exposure resulted in reduction in plant growth, death of older leaves or whole plants and fewer numbers of tillers in most of the selected mutant lines as well as the WT. Based on a growth comparison of the entries, we have inves-tigated the survival rate. Out of the selected 56 mutant lines, the putative 2 ST-lines were finally selected. The survival rates of the WT, ST-495 and ST-532 were 36.6%, 70% and 50%, respectively (Table 1). To examine whether the enh-anced salt tolerance in the plants, seeds of 2 ST-lines and WT were germinated and grown in soil trays (without salt) for 14 days, then irrigated them with 171 mM salt solution for 7 days. Symptoms of major damage caused by salt stress, such as wilting and yellowing of old leaves, and death of older leaves or whole plants were moderated of delayed in the 2 ST-lines than those of the WT (Fig. 1A). Tested plants were transferred into tap water in order to recover and resume. The shoots of 2 ST-lines significantly grew greater than the WT plants during this recovery period. After 7 days, the growth length of WT was about 0.1 cm while ST-495 and ST-532 showed about 1.3 and 2.8 cm, respectively (Fig. 1B). To estimate whether their tolerance to high salinity at the seedling stage, 2 salt tolerant lines were initially cultured on selection medium containing 171 mM NaCl for 40 days. We found that salt stress significantly affected the growth and survival of both ST-lines and WT plants. Three weeks after a treatment with 171 mM NaCl, it was found that the seedlings had gradually wilted, but more severe in the WT than the ST lines. This result means that 2 mutant lines had significantly increased salt tolerance and these reasons were due to the alteration of genes related to the salt stress mech-anisms such as ion membrane transport and osmoprotectant biosynthesis, etc.
2. Measurement of lipid peroxidation
Lipid peroxidation contents in shoots of the WT and 2 ST-lines, measured as the content of malonaldehyde (MDA), are recorded in Fig. 2. Salt stress caused an increase in the MDA contents among the tolerant and sensitive lines. The MDA contents showed almost no difference (average 6.1
nmol g-1F.W.) in 2 ST-lines and WT plant under normal
growth conditions, while the mutant lines including WT plant were gradually increased in earlier days until 7 days after salt treatment and the difference of MDA contents be-came apparent after 7 days between mutant lines and WT.
The WT plant (23.6 nmol g-1F.W.) enhanced greater than
ST-495 (11.6 nmol g-1F.W.) and ST-532 (12.6 nmol g-1
F.W.) (Fig. 2). Luna et al. (2000) mentioned that MDA con-tents are higher in sensitive groups than in tolerant groups. The accumulation of MDA in seedlings increased with increasing salinity stress in the all plants including ST-lines and WT, but showed greater change in the WT plant than the tolerant lines. The low values of MDA content is impor-tant sign of oxidative damage limiting capacity under salinity as announced in different studies by other authors (Liang
et al. 2003; Ruiz et al. 2005). MDA is an end product of
lipid peroxidation in biomembranes, and the MDA content usually reflects the level of lipid peroxidation and indirectly reflects the extent of membrane injury (Wang et al. 2010). Our study showed that 2 ST-lines lines had significantly lower MDA contents than the WT lines.
3. Chlorophyll and carotenoid contents
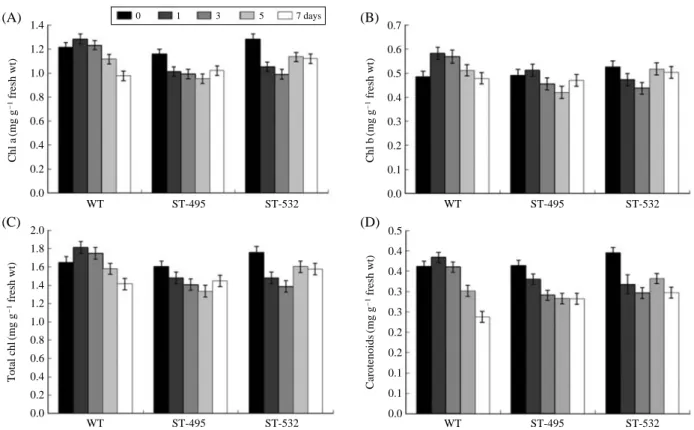
Decrease of the degree of chlorophyll and carotenoid content is a common phenomenon in salt stress conditions.
The chlorophyll a, b and a++b contents in WT plant were
gradually decreased until 7 days after salt treatment. However, salt tolerant lines did not show any notable change during the period. The carotenoid contents decreased more in a WT plant than two tolerant mutants during whole process after salt treatment (Fig. 3). Decreased chlorophyll and carotenoid contents after the salt treatment in WT were observed after 7 days treatment. The chlorophyll and carotenoid contents were decreased more in a WT plant than two tolerant mutants. Degradation of chlorophyll and carotenoid contents is a normal status in abiotic stress conditions (Sultana et al. 1999). The chlorophyll pigment in rice is very sensitive to salt Fig. 2. Malondealdehyde (MDA) content of the wild type and 2 salt
tolerant lines in 171 mM salt solution for 7 days. 25 20 15 10 5 0 MDA (nmol g -1fresh wt) WT 495 532 0 1 3 5 7 days
stress (Bahaji et al. 2002), especially in salt susceptible vari-ety (Asch et al. 2000). Chlorophyll degradation in salt
stres-sed rice is positively related to net photosynthetic rate (Pn)
reduction or CO2assimilation as well as stomatal conductance
and transpiration rate (Sultana et al. 1999). In addition, the
chlorophyll and Pnreduction percentages in salt tolerant
rice are lower than those in salt sensitive rice (Cha-um et al. 2007). The physiological changes in salt stressed rice are directly impaired on overall growth (Khan and Panda 2008).
In vitro culture in rice in combination with gamma-ray
induced mutations is an effective way to improve salt-toler-ance in rice. Variations in the progenies of in vitro mutage-nized plant materials have already been reported and probab-ly originated from the genetic instability created during plant tissue culture in combination with induced mutations (Malus-zynski et al. 1995). The mutation and selection frequency were greatly increased, and the selected mutants were stable. Such mutant lines could be utilized as a source of salt-tolerant germplasm. In this study, we introduced the selection proce-dures for the selection of salt tolerant rice mutants, and pro-mising 2 mutant lines, ST-495 and ST-532, were finally selected. These rice mutant lines will be released for
cultiva-tion at the reclaimed land and used as a control plot for genetic research about salt tolerance.
ACKNOWLEDGMENT
This work was supported by a grant (Code 308020052S B040) from Agricultural R&D Promotion Center, Ministry for Food, Agriculture, Forestry & Fisheries and a grant (Code 20070501034005) from BioGreen 21 Program, RDA (Rural Development Administration), Republic of Korea.
REFERENCES
Asch F, Dingkuhn M and Dorffling K. 2000. Salinity increase CO2assimilation but reduces growth infield-grown, irrigated
rice. Plant Soil 218:1-10.
Bahaji A, Mateu I, Sanz A and Cornejo MJ. 2002. Common and distinctive responses of rice seedlings to saline- and osmo-tically-generated stress. Plant Growth Regul. 38:83-94. Carlson PS. 1970. Induction and isolation of auxotrophic
mu-tants in somatic cell cultures of Nicotiana tabacum. Science
168:487-489.
Fig. 3. Chlorophyll a content (A), chlorophyll b content (B), total chlorophyll (C) and carotenoid content (D) of the wild type and 2 salt tolerant lines in 171 mM salt solution for 7 days.
0 1 3 5 7 days 1.4 1.2 1.0 0.8 0.6 0.4 0.2 0.0 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0.0 0.5 0.4 0.4 0.3 0.3 0.2 0.2 0.1 0.1 0.0 2.0 1.8 1.6 1.4 1.2 1.0 0.8 0.6 0.4 0.2 0.0 Chl a (mg g -1fresh wt) Chl b (mg g -1fresh wt) Carotenoids (mg g -1fresh wt) Total chl (mg g -1fresh wt) WT ST-495 ST-532 WT ST-495 ST-532 WT ST-495 ST-532 WT ST-495 ST-532 (A) (B) (C) (D)
Cella R, Parisi B and Nielsen E. 1982. Characterization of a carrot cell line resistant to azetidine-2-carboxylic acid. Plant
Sci. Lett. 24:125-135.
Cha-um S, Supaibulwatana K and Kirdmanee C. 2007. Glycine-betaine accumulation, physiological characterizations, and growth efficiency in salt tolerant and salt sensitive lines of indica rice (Oryza sativa L. spp. Indica) response to salt stress. J Agron Crop Sci. 193:157-166.
Delauney AJ and Verma DPS. 1993. Proline biosynthesis and osmoregulation in plants. Plant J. 4:215-223.
Dhindsa RS and Matowe W. 1981. Drought tolerance in two mosses correlated with enzymatic defense against lipid peroxidation. J. Exp. Bot. 32:79-91.
Ding YC, Chand CR, Luo W, Wu YS, Ren XL, Wang P and Xu GH. 2008. High potassium aggravates the oxidative stress induced by magnesium deficiency in rice leaves.
Pedosphere 18(3):316-327.
Hyun DY, Lee IS, Kim DS, Seo YW and Lee YI. 2003. Selec-tion of azetidine-2-carboxylic acid resistant cell lines by in
vitro mutagenesis in rice (Oryza sativa L.). J. Plant Biotech.
5(1):43-49.
Khan MH and Panda SK. 2008. Alterations in root lipid peroxida-tion and antioxidative responses in two rice cultivars under NaCl-salinity stress. Acta. Physiol. Plant 30:81-89. Kim DS, Lee IS, Jang CS, Hyun DY, Seo YW and Lee YI. 2004.
Selection of radiation induced 5-methyltryptophan resistant rice mutants through embryo culture. Euphytica 135(1):9-19. Larkin PJ and Scowcroft WR. 1981. Somaclonal variation - a novel source of variability from cell cultures for plant impro-vement. Theor. Appl. Genet. 60:197-214.
Lee IS, Kim DS, Lee SJ, Lim YP and Lee YI. 2003. Selection and characterizations of radiation-induced salinity tolerant lines via rice embryo culture. Breed Sci. 53(4):313-318. Liang Y, Chen Q, Liu Q, Zhang W and Ding R. 2003. Exogenous
silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 160(10):1157-1164.
Lichtenthaler HK. 1987. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Method Enzymol. 148:350-382.
Lodato RF, Smith RJ, Valle DL and Crane K. 1984. Mutant
cell lines resistant to azetidine-2-carboxylic acid: alterations in the synthesis of proline from glutamic acid. J. Cell
Phy-siol. 119:137-143.
Luna C, Garcia Seffino L, Arias C and Taleisnik E. 2000. Oxida-tive stress indicators as selection tools for salt tolerance in Chloris gayana. Plant Breeding 119:341-345.
Maluszynski M, Ahloowalia BS and Sigurbjornsson B. 1995. Application of in vitro mutation techniques for crop improve-ment. Euphytica 85:303-315.
Miflin BJ, Bright SW, Rognes SE and Kueh JHS. 1983. Amino acids, nutrition and stress: the role of biochemical mutants in solving problems of crop quality. In: Kosuge T, Meredith CP, Hollaender A (eds.), Genetic engineering of plants-an agricultural perspective, pp. 391-414. Plenum Press, London. Ruiz JM, Blasco B, Rivero RM and Romero L. 2005. Nicotine-free and salttolerant tobacco plants obtained by grafting to salinity-resistant rootstocks of tomato. Physiol. Plant. 124: 465-475.
Schaeffer GW and Sharpe FT. 1983. Mutations and selection: genetic variation for improved protein in rice. Genetic Engineering, Application to Agriculture; Invited papers presented at a symposium, May 16-19, 1982, Beltsville, pp. 237-254.
Song JY, Kim DS, Lee G-J, Lee IS, Kang KK, Yun SJ and Kang S-Y. 2007. Characterization of salt tolerant rice mutant lines derived from Azetidine-2-carboxylic acid resistant cell lines induced by gamma ray irradiation. J. Plant Biotech. 34(1):61-68.
Sultana N, Ikeda T and Itoh R. 1999. Effect of NaCl salinity on photosynthesis and dry matter accumulation in developing rice grains. Environ. Exp. Bot. 42:211-220.
Wang YC, Qu GZ, Li HY, Wu YJ, Wang C, Liu GF and Yang CP. 2010. Enhanced salt tolerance of transgenic poplar plants expressing a manganese superoxide dismutase from Tamarix androssowii. Mol. Biol. Rep. 37:1119-1124.
Widholm JM. 1974. Cultured carrot cell mutants: 5-methyltryp-tophan-resistant trait carried from cell to plant and back.
Plant Sci. Lett. 3:323-330.
Manuscript Received: June 8, 2010 Revision Accepted: June 14, 2010