저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
이학 석사학위 논문
G2 기에서 ATP 결핍이
세포분열 개시에 미치는 영향
아 주 대 학 교 대 학 원
의생명과학과/ 종양생물학전공
이 진 삼
G2 기에서 ATP 결핍이
세포분열 개시에 미치는 영향
지도교수 이 재 호
이 논문을 이학 석사학위 논문으로 제출함.
2018년 2월
아 주 대 학 교 대 학 원
의생명과학과/ 종양생물학전공
이 진 삼
i -국문요약-
G2 기에서 ATP 결핍이 세포분열 개시에 미치는 영향
진핵세포는 다양한 스트레스에 반응하여 세포주기를 조절하는 checkpoint를 가지고 있다. 진핵세포의 세포주기에서 이용 가능한 ATP의 양에 따라 2 가지 주요 energy checkpoint를 갖는 것으로 보고되었다. 이 checkpoint는 G1 기 동안 종양 세포주기의 진행과 G2 / M transition의 진행을 조절한다. G1 energy checkpoint에서는 AMPK에 의해 cyclin E가 억제되는 molecular mechanism이 보고되었지만 G2 energy checkpoint는 아직 완전히 설명되지 않았다. 본 저자는 G2 기에서 ATP 결핍이 세포주기에 어떤 영향을 미치는지 알아보고자 하였다. ATP를 결핍 시키는데 사용한 약물은 glycolysis를 억제하는 2-DG와 mitochondrial Cytochrome C oxidase를 억제하는NaN3를 사용하였다. Drugs를 처리하였을 때 ATP가 asynchronous 수준으로
떨어지는 것을 확인하였다. 그 다음 mitotic entry와 cyclin B nuclear translocation을 확인한 결과 mitotic entry와 cyclin B nuclear translocation이 차단된 것을 관찰할 수 있었다. 이러한 현상이 drugs을 wash out 해 주었을 때 정상수준으로 회복되는지 알아보고자 하였다. 그 결과 drugs를 wash out 했을 때 ATP와 mitotic index가 정상 수준으로 회복하였다. 그 다음 세포주기가 멈추는 현상이 정말로 ATP결핍에 의한 현상인지 알아보고자 또 다른 energy stress를 가해주었다. 2-DG와 mitochondrial complex I 을 억제하는 phenformin을 동시에 처리한 결과 세포주기가 정지되는 것을 확인하였다. 그 다음 glucose free media로 glucose starvation 시켰을 때에도 이러한 현상이 일어나는지 확인하였다. 그 결과 drugs 처리 때보다는 약한정도 였으나 어느정도 세포분열기로의 이행이 억제되는 것을 볼 수 있었다. 그 다음 세포내 ATP를 측정한 결과 2-DG와 phenformin을 처리하였을 때는
ii
asynchronous 수준으로 떨어지는 것을 확인하였다. 하지만 glucose starvation 조건에서는 ATP가 조금 감소하는 정도였다. 따라서 세포내 ATP의 양에 따라 세포주기가 조절된다는 것을 알 수 있었다. 이러한 현상이 일어나는 기전으로 먼저 AMPK에 의한 것인지 알아보고자 하였다. siRNA와 inhibitor를 사용하여 AMPK를 억제하였지만 세포주기 진행억제가 극복되지 않았다. 따라서 G2 energy checkpoint에서 AMPK는 큰 영향을 미치지 않는다는 것을 확인하였다. 그 다음 DNA 손상에 의한 신호전달 경로를 확인한 결과 Chk1이 인산화 되는 것을 확인하였다. 다양한 inhibitor를 통해 Chk1의 인산화를 억제하였지만 세포주기 진행억제를 극복하지 못했다. 따라서 Chk1도 별다른 영향을 미치지 않는다는 것을 알 수 있었다. 그 다음 ATP가 결핍되었을 때 Wee1 kinase에 의해 인산화 되는 p-Cdk1(Y15)이 줄어들지 않는 것을 확인하였다. Cdk1의 인산화를 조절 하는 Wee1과 Cdc25C의 protein level을 western blot을 통해 확인한 결과 ATP가 결핍되었을 때 protein level이 변화하지 않고 drugs 처리 전 수준으로 유지되는 것을 확인하였다. 따라서 Wee1의 kinase activity를 측정하고자 in vitro kinase assay를 수행하였다. 그 결과
2-DG와 NaN3를 처리하였을 때 Wee1의 kinase activity가 높은 것을
확인하였다. 따라서 ATP를 결핍 시킨 다음 Wee1 inhibitor로 알려진 PD166285를 처리하고 mitotic index와 western blot을 수행한 결과 P-Cdk1(Y15)가 줄어들고 mitotic entry가 일어나는 것을 확인하였다. 그리고 Time-lapse를 통해서도 확인한 결과 세포주기가 전행 되는 것을 확인하였다. 그런데 Wee1 억제에 의해 mitosis에 들어간 세포들이 mitosis에 세포주기가 멈추는 것을 확인하였다. 세포가 mitosis에 계속 머무르게 되면 세포 사멸이 일어난다는 보고가 있기 때문에 Trypan-blue staining을 통해 cell death를 확인하였다. 그 결과 Wee1 억제를 통해 mitosis로 들어간 세포에서 대부분 세포사멸이 일어나는 것을 확인하였다. 이러한 결과들은 G2 기에서 energy stress가 Wee1에 의한 Cdk1의 억제를 통해 세포주기를 정지시켰음을 시사한다. 나아가 Wee1 억제를 통해 이 세포들이 분열하도록 유도할 수
iii
있는데 이렇게 mitosis로 들어간 세포들은 세포사멸을 일으킨다는 점에서 새로운 항암화학요법의 가능성을 제시한다고 하겠다.
iv
차 례
국문 요약 ... i
차례 ... iv
그림 차례 ... vi
Ⅰ. 서론 ... 1
Ⅱ. 실험재료 및 방법 ... 6
A. 세포배양 ... 6
B. 항체와 시약 ... 6
C. ATP assay ... 6
D. Mitotic index ... 7
E. Immunocytochemistry(ICC) ... 7
F. 단백질 전기 영동 ... 7
G. 면역 침강법 ... 8
H. In vitro kinase assay ... 8
I. siRNA ... 9
J. Time-lapse analysis ... 9
K. 통계적 분석 ... 10
Ⅲ. 결과 ... 11
A. G2 phase 에서 ATP 결핍에 의한 세포주기 정지 확인 ... 11
B. ATP 결핍에 의한 cyclin B 의 핵으로의 이동억제 확인 ... 14
C. Energy-stress 가 세포주기에 미치는 영향 ... 17
D. AMPK 가 G2/M transition 에 미치는 영향 ... 22
v
F. ATP 결핍이 Wee1 과 Cdc25C 에 미치는 영향 ... 28
G. Wee1 의 억제를 통한 G2-M transition 확인 ... 31
H. Wee1 의 억제를 통해 mitosis 로 들어간 세포의 운명 확인 ... 35
Ⅳ. 고찰 ... 39
Ⅴ. 결론 ... 42
참고 문헌 ... 44
ABSTRACT ... 48
vi
그림 차례
Figure 1. Cell cycle checkpoint regulates cell cycle progression ... 1
Figure 2. Energy checkpoints in cell cycle ... 3
Figure 3. G1 energy checkpoint ... 4
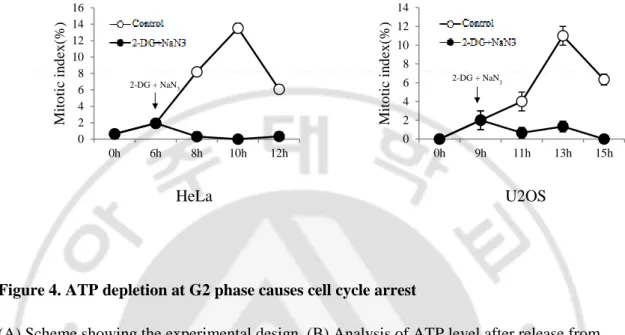
Figure 4. ATP depletion at G2 phase causes cell cycle arrest ... 13
Figure 5. ATP depletion inhibits nuclear translocation of Cyclin B ... 16
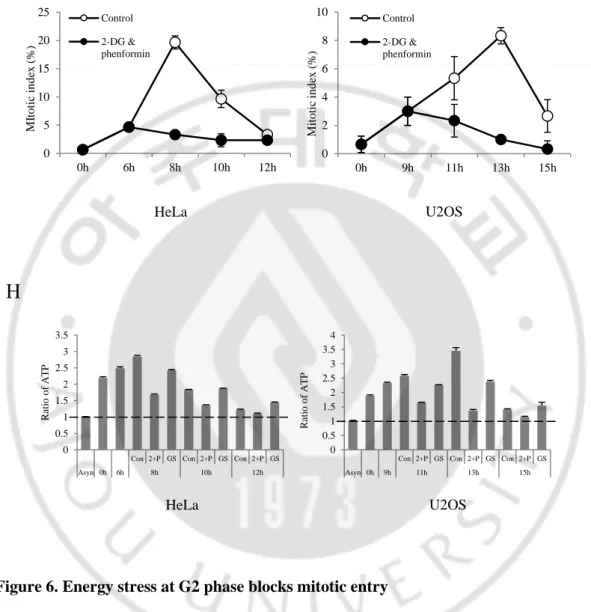
Figure 6. Energy stress at G2 phase blocks mitotic entry ... 21
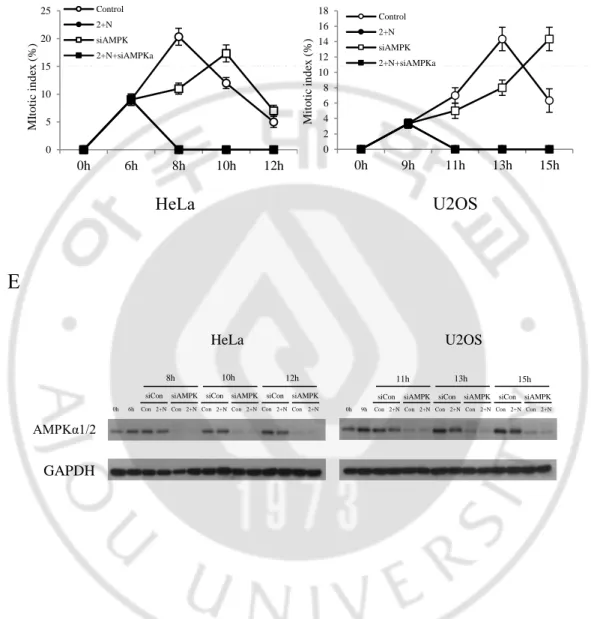
Figure 7. AMPK did not affect G2/M transition during ATP depletion ... 24
Figure 8. DNA damage did not affect G2/M transition during ATP depletion .. 27
Figure 9. ATP depletion affect Cdk1 regulator ... 30
Figure 10. PD166285 dramatically overcome G2 arrest by ATP depletion ... 34
1
I. 서론
진핵세포는 세포내 외부에서 일어나는 스트레스에 반응하여 세포주기 진행을 조절하는 조절 기전을 가지고있다. 이러한 조절 경로를 checkpoint 라고 부른다 (Paulovich et al., 1997). Checkpoint 는 여러가지 단백질들에 의해 조절되며 가장 잘 알려진 것으로는 G1, G2, mitosis 에 존재하는 checkpoint 이다.
Figure 1. cell cycle checkpoint regulates cell cycle progression
G1 phase 에서는 Rb, p107, p130 이 G1-S transition 을 억제한다. Rb 는 E2F 에 결합하여 E2F 를 억제하고, p107 및 p130 은 E2F4 및 E2F5 와 각각 결합하여 G1-S transition 을 촉진하는 단백질의 전사를 억제한다 (Hurford et al., 1997). 그리고 G1 phase 일 때 주변환경에 성장인자가 존재할 경우 Akt 는 FoxO1/3 를
인산화 하는데 이것으로 인해 FoxO1/3 가 핵 밖으로 나가는 기능을 G1 S G2 M G1 Checkpoint G2 Checkpoint
Spindle Assembly Checkpoint
Check for: - Cell size - DNA replication Check for: - Chromosome attachment to spindle Check for: - Cell size - Nutrients - Growth factors - DNA damage DNA synthesis
2
억제함으로써 세포의 생존과 증식을 가능하게 한다 (Tzivion et al., 2011). 이러한 조절 기전을 G1 checkpoint 라고 한다. TGF-β 나 DNA 손상 등 다양한 자극에 의해서도 G1 checkpoint 가 활성화 된다 (Petritsch, 2000; Bartek and Lukas, 2001a, b). 이러한 자극은 cyclin dependent kinase inhibitor (CKIs)인 INK4 또는 Cip/Kip 계열의 구성원을 유도하는 전사인자를 통해 작용한다 (Reynisdottir et al., 1995; Boxem and van den Heuvel, 2001; Zhang et al., 2009). 특히 Bmi1 은 암에서 INK4 A/B 발현의 음성 조절인자로서 작용한다 (Vonlanthen et al., 2001). CKI 를 조절하는 것 말고도 TGF-B 는 CDK 의 활성에 직접적으로 관여하는 Cdc25A 의 전사를 억제한다 (Bhowmick et al., 2003).
G2 phase 에서는 DNA 손상으로 인한 신호전달 경로를 통해 Cdk1 의 인산화를 통해 G2-M transition 을 억제한다 (O'Connell et al., 1997). DNA 가 손상되면 ATM/ATR kinase 가 활성화되고, 활성화된 ATM/ATR 은 Chk1/2 를 인산화 한다 (Falck et al., 2005). 인산화 된 Chk1/2 는 cdc25C 를 억제함으로써 Cdk1 의 활성을 억제한다 (Reinhardt and Yaffe, 2009). 또다른 신호전달 경로는 p53 과 MDM 의 결합을 억제함으로 p53 을 활성화 하는 것이다. 이로 인해 p53 이 활성화 되고, 이는 14-3-3 를 통해 인산화 된 Cdk1-cyclin B 를 핵으로부터 내보낸다 (Hermeking et al., 1997). 결과적으로 G2 phase 에서는 DNA 의 손상으로 인해 발생한 신호전달 경로를 통해 Cdk1 – cyclin B complex 가 억제된다. 이러한 세포주기 조절을 G2 checkpoint 라고 한다.
Mitosis 에서는 spindle assembly checkpoint 가 존재하여 각 염색체의
kinetochore 에 spindle 이 제대로 부착될 때까지 염색체의 분리를 억제한다. 세포가 정상적으로 기능하기 위해서는 세포분열 이후에 적절한 염색체 수를 가져야한다. 염색체 수가 정상보다 적거나 많을 경우, 세포사멸을 유발할 수 있다 (Musacchio and Salmon, 2007). 만약 세포사멸이 일어나지 않을 경우에는 치명적인 표현형 결과를 초래할 수 있다.
3
많은 암세포에서는 이러한 checkpoint 를 조절하는 여러 유전자가 돌연변이 되어있다 (Squire et al., 1986; Cahill et al., 1998). 따라서 정상세포와는 달리 세포주기가 진행될 상황이 아님에도 불구하고 세포 분열이 진행됨으로써 무한정 증식할 수 있다. 이러한 암세포의 비정상적인 증식을 막기 위해서는 암세포의 세포주기를 이해하고 이를 조절할 수 있는 연구가 필요하다.
암세포가 분열하기 위해서는 많은 양의 에너지와 세포분열에 필요한 재료들이 필요하다. 이러한 결과로 인해 초기 암세포는 어느 정도 성장하다가 결국 energy stress 에 빠지게 된다. 이렇게 되면 세포내 ATP 양이 줄어들게 되고 결국 AMP 의 양이 증가하게 된다. 이렇게 많아진 AMP 는 AMPK 에 결합하여 AMPK 의 입체구조를 바꾸게 되고, 이로 인해 LKB1/CaMKK 에 의해 α subunit 의 Threonine 172 site 가 인산화 되어 활성화 하게 된다 (Carling et al., 2008; Hardie et al., 2012). 활성화된 AMPK 는 세포내에서 fatty acid synthesis 를 억제하고 fatty acid oxidase 를 촉진함으로써 세포내 에너지 균형을 맞춘다. 또한 angiogenesis 를 유도하여 혈관을 신생하고 영양소를 공급받을 수 있게 해준다 (Nagata et al., 2003).
4
세포가 성장하고 분열하기 위해서는 세포주기를 조절하는 CDK 와 Cyclin 복합체의 활성이 필수적이다. 이 복합체는 ATP 를 사용하여 여러 기질단백질을 인산화하기 때문에 암세포가 성장하려면 많은 양의 ATP 가 필요하게 된다 (Arellano and Moreno, 1997). 암세포가 증식하면서 세포내 ATP 의 양을 세포주기 별로 나타낸 것을 보면 G1 에서 S 로 넘어가지 전까지 증가하 감소한다. 이후에 G2 까지 ATP 의 양이 다시 증가하다가 mitosis 로 넘어갈 때 감소한다. 이렇게 각 G1 과 G2 에 ATP threshold 가 존재하는데 이것을 energy checkpoint 라고 한다 (Chignola and Milotti, 2005).
G1 energy checkpoint 에서 G1 에서 S 로 넘어가면서 미토콘드리아의 membrane potential 이 증가하게 된다. 이로 인해 ATP 생성이 증가하고 cyclin E 의 번역이 증가하여 G1-S transition 이 일어난다. 그런데 만약 ATP 가 부족할 경우 AMPK 가 활성화 되고, 이로 인해 p53 과 p21 이 활성화되어 cyclin E 의 level 이 낮아지게 되어 세포주기가 멈추게 된다 (Finkel and Hwang, 2009). 이렇게 G1 energy checkpoint 에 대해서는 잘 알려졌지만, 아직 G2 energy checkpoint 에 대해서는 많이 알려지지 않았다.
5 G2 energy checkpoint 에 대해서 연구된 것으로는 분열 효모에서 주변 환경의 포도당 농도의 급격한 변화에 대한 반응으로 세포주기의 진행을 조절하는 것이 밝혀졌다. 포도당이 부족할 때 야생형 효모는 G2 단계에서 세포주기를 멈춘다. 이러한 과정은 Wee1 이 Cdk1 을 억제하는 과정을 통해 이루어진다고 밝혀졌다. Wee1 이 결핍된 돌연변이의 경우에는 포도당을 제한했을 때 G2 에서 세포주기가 멈추지 않아 야생형 세포보다 빠르게 생존율이 감소하는 것을 보였다. 결과적으로 이러한 연구 결과는 암 치료를 위한 좋은 분자 표적이 될 수 있는 세포내 에너지를 모니터링하는 새로운 checkpoint 의 존재를 나타낸다 (Masuda et al., 2016). G2 기 에서 Mitosis 로 진행하는데 있어 가장 중요한 단백질은 Cdk1 과 Cyclin B 이다. Cdk1 과 Cyclin B 는 서로 결합하여 mitotic substrate 들을 인산화 시킴으로써 G2 에서 mitosis 로의 진행을 조절한다. G2 기 말기에 Cdk1 과 cyclin B 의는 centrosome 에 모여 결합한다. Cyclin B 에는 nuclear import sequence 가 존재하여 Cdk1 과 결합하면 이 sequence 가 노출되고 이로 인해 Cdk1-Cyclin B 복합체는 핵 안으로 들어간다. 그런데 핵안으로 들어갔는데 핵 안에 Wee1 이 활성화 되어있으면 Cdk1 의 tyrosine 15 site 가 인산화 되고, 복합체가 핵 밖으로 나가게 된다. Cdk1 의 Y-15 의 인산화는 Cdc25C 에 의해 탈 인산화 된다. 따라서 mitosis 에 진입에 있어 Cdk1 의 탈 인산화가 중요하다고 할 수 있다 (Lindqvist et al., 2009). 따라서 본연구에서는 G2 phase 에서 ATP 를 결핍 시키기 위해 몇가지 방법을 사용하였다. 먼저 glycolysis 를 억제하는 2-DG 와 mitochondrial cytochrome C
oxidase 를 억제하는 NaN3 를 동시에 처리하였다 (Ralser et al., 2008). 또 다른
방법으로는 2-DG 와 mitochondrial complex I 을 억제하는 phenformin 도 동시에 처리하였다 (Wheaton et al., 2014). 마지막으로 glucose starvation 을 통해서 ATP 결핍을 유도 하였다. 이를 통해 G2 기 에서 ATP 결핍이 세포주기에 어떤 영향을 미치는지 알아보고 이에 대한 메커니즘에 대하여 알아보고자 하였다.
6
II. 실험 재료 및 방법
A. 세포배양
HeLa 세포는 10% fetal bovine serum (Young-in frontier, USA) 과 1% antibiotics (GIPCO)가 첨가된 DMEM/F-12 (GIPCO) 배지를 이용하여 37℃, 5% 이산화탄소 조건에서 배양하였다. U2OS 세포는 10% fetal bovine serum (Young-in frontier, USA) 과 1% antibiotics (GIPCO)가 첨가된 DMEM(high glucose) (GIPCO) 배지를 이용하여 37℃, 5% 이산화탄소 조건에서 배양하였다.
B. 항체와 시약
1 차 항체인 Cyclin B, GAPDH, Wee1, Cdc25C, IgG 는 Santa Cruz 에서 구입하였으며, AMPKα1/2, phospho-Chk1, phospho-Cdk1(Y15), Cell signaling 에서 구입하였다. Lamin B1 은 Abcam 에서 구입하였다. γ–H2AX 는 EMD
Millipore 에서 구입하였다. Thymidine, Phenformin, Mannitol, H2O2, Caffeine,
PD166285 는 SIGMA-ALDRICH 에서 구입하였다. Compound C 는
CalBiochem 에서 구입하였다. D-glucose 는 AMRESCO 에서 구입하였다. Trypan blue 는 ACROS 에서 구입하였다.
C. ATP assay
세포 내 ATP 는 ATP 분석 키트 (Perkin Elmer, 6016941)를 사용하여 측정 하였다. HeLa 와 U2OS 세포를 60 mm culture dish 에 seeding 하고 thymidine 을 HeLa 는 1 mM 로 20 시간, U2OS 는 2.5 mM 로 22 시간 처리하여 세포주기를 G1/S 기 경계에 정지시킨 후 PBS 로 두 번
세척하였다. HeLa 는 6 시간 후, U2OS 는 9 시간 후에 2-DG 6 mM, NaN3 10
mM 처리하였다. 시간별로 세포를 얻은 다음 cell counting 하였다. 그 다음 동일한 수의 세포 가 있는 세포용액 100μl 에 cell lysis solution
7
50μl 을 넣고 5 분간 반응시켰다. 그 다음 substrate solution 50μl 을 넣고 5 분간 반응시켰다. 이후 암조건에서 10 분간 incubation 한 다음 Victor 3
Model 1420-012 Multi-label Microplate Reader 를 사용하여 발광을
측정하였다.
D. Mitotic index
HeLa 와 U2OS 세포를 60 mm culture dish 에 seeding 하고
thymidine 을 HeLa 는 1 mM 로 20 시간, U2OS 는 2.5 mM 로 22 시간 처리하여 세포주기를 G1/S 기 경계에 정지시킨 다음 PBS 로 두 번 세척하였다. 이후 시간 및 조건 별로 얻은 세포를 얻은 세포를 슬라이드 글라스에 aceto-orcein 용액과 함께 섞은 후 커버 글라스를 덮어주었다. 그 다음 광학현미경을 이용하여 체세포 분열중인 세포의 수를 측정하였다.
E. Immuno-cyto chemistry (ICC)
커버 글라스에 poly-L-lysine 을 30 분 동안 처리하여 코팅하였다. 그 다음 6 well plate 에 코팅된 커버 글라스를 넣고 HeLa 와 U2OS 세포를 seeding 하였다. 이후 세포를 PBS 로 세척한 다음 100% methanol 로 -20℃에서 10 분간 고정하였다. 그 다음 PBS 로 두 번 세척하고 3% BSA 가 들어 있는 PBS 로 상온에서 1 시간 동안 blocking 하였다. 그 다음 4℃에서 Lamin B1, Cyclin B 1 차 항체로 하루 동안 반응 시켰다. 반응 후 PBS 로 세 번 세척하고 상온에서 3% BSA 가 들어있는 2 차 항체로 암 조건에서 1 시간 동안 반응 시켰다. 그 다음 PBS 로 두 번 세척 후 DAPI 염색을 하고 형광 현미경으로 관찰 하였다. F. 단백질 전기 영동
세포를 얻어 PBS 로 세척 후 RIPA cell lysis buffer (10 mM Tris-HCl (pH 7.4), 150 mM NaCl, 1 % Triton X-100, 1 µg/ml aprotinin, 1 µg/ml leupeptin, 1
mM NaF, 5m M Na3VO4)로 30 분 동안 ice 에서 incubation 하였다. 그 다음
8
그 다음 SDS-PAGE 를 이용해 분리하고 PVDF 에 전기이동(electro transfer)하였다. 그 다음 5% nonfat dry milk 가 들어있는 TBST 로 상온에서 1 시간동안 blocking 후 4℃에서 1 차 항체로 하루 동안 반응 시켰다. 반응 후 TBST 로 10 분씩 3 번 세척하고 2 차 항체로 상온에서 1 시간동안 반응 시켰다. 그 다음 TBST 로 10 분씩 3 번 세척하고 ECL 용액을 이용하여 단백질의 발현을 X-ray film 에 노출시켜 확인하였다.
G. 면역 침강법
세포를 얻어 PBS 로 세척 후 IP lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 0.5 % NP-40, 5 mM EDTA)에 10 분동안 incubation 한 후 10 초씩 3 번 30% Amplification 으로 sonication 하였다. 그 다음 microcentrifuge 로 13000 rpm 에서 원심분리하고 정량 하였다. 400μg 의 세포용액에 Wee1 항체 1 μg 을 넣고 4 ℃에서 overnight 하였다. 그 다음 20μl 의 protein G Sepharose (GE Healthcare Bio-Science, SWEDEN)을 넣고 4℃에서 한시간 동안 incubation 한다. 이 후 IP buffer 를 이용하여 bead 를 3 번 세척하였다. 상층액은 모두 버리고 bead 에 no-reducing sample buffer 를 50μl 넣은 후 10 분 동안 끓여주었다. Sample 은 SDS-PAGE 를 이용해 분리한 후 Nitrocellulose 에 전기이동(electro transfer)하고, 5% nonfat dry milk 가 들어있는 TBST 로 상온에서 1 시간동안 blocking 후 4℃에서 1 차 항체로 하루 동안 반응 시켰다. 반응 후 TBST 로 10 분씩 3 번 세척하고 2 차 항체로 상온에서 1 시간동안 반응 시켰다. 그 다음 TBST 로 10 분씩 3 번 세척하고 ECL 용액을 이용하여 단백질의 발현을 X-ray film 에 노출시켜 확인하였다.
H. In vitro kinase assay
세포를 얻어 PBS 로 세척 후 IP lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 0.5 % NP-40, 5 mM EDTA)에 10 분동안 incubation 한 후 10 초씩 3 번 30% Amplification 으로 sonication 하였다. 그 다음 microcentrifuge 로 13000
9
rpm 에서 원심분리하고 정량 하였다. 그 다음 400μg 의 세포용액에 Wee1 항체 1 μg 을 넣고 4 ℃에서 overnight 하였다. 그 다음 20μl 의 protein G Sepharose (GE Healthcare Bio-Science, SWEDEN)을 넣고 4℃에서 한시간 동안 incubation 하였다. 그 다음 IP buffer 로 2 번 세척한 다음 kinase buffer (25 mM Tris-Hcl (pH7.5), 5 mM β-glycerophosphate, 2 mM
dithiothreitol(DTT), 0.1 mM NaVO4, 10 mM MgCl2)로 1 번 세척하였다.
상층액을 모두 제거한 다음 kinase buffer 40 μl 에 200 μM ATP 와 substrate 0.5 ~ 1 μg 을 넣고 bead 와 섞어준 다음 30 ℃에서 반응시켰다. 그 다음 microcentrifuge 로 4500 rpm 에서 원심분리 하여 bead 를 down 시키고 상층액을 SDS-PAGE 를 이용해 분리하고 PVDF 에 전기이동(electro transfer)하였다. 그 다음 5% nonfat dry milk 가 들어있는 TBST 로 상온에서 1 시간동안 blocking 후 4℃에서 1 차 항체로 하루 동안 반응 시켰다. 반응 후 TBST 로 10 분씩 3 번 세척하고 2 차 항체로 상온에서 1 시간동안 반응 시켰다. 그 다음 TBST 로 10 분씩 3 번 세척하고 ECL 용액을 이용하여 substrate 의 인산화를 X-ray film 에 노출시켜 확인하였다.
I. siRNAs
AMPK siRNA 는 Santa Cruz Biotechnology 에서 구입하였으며
Oligofectamine Reagent (Invitrogen)을 이용하여 제조회사의 protocol 로
transfection 하였다.
J. Time-lapse
HeLa 세포를 4-well Glass-based dish (NuncTM Lab-Tak II Chambered
Coverglass, 155382)에 seeding 하고 thymidine 1 mM 을 20 시간동안 처리한
다음 release 하였다. 6 시간 후에 2-DG 와 NaN3를 처리하여 ATP 를 결핍
시키고, 2 시간 후에 PD166285 를 처리하여 Wee1 을 억제하였다. 이미지는 5 시간 동안 Nikon eclipse Ti 의 plan-Apochromat objective 로
10
10 분마다 촬영되었고 NIS elements Ar microscope imaging software 를 통해 얻었다.
K. 통계적 분석
그래프는 평균 값 ±표준 편차로 나타냈다. 그리고 그룹 간의 유의성 검증을 위해 Student’s t-test 를 사용하였다. P 값이 0.05 이하일 때 통계적으로 유의 하다고 판단하였다.
11
III. 결과
A. G2 phase 에서 ATP 결핍에 의한 세포주기 정지 확인
G2 phase 에서 ATP 가 결핍되면 G2-M 전이가 어떻게 되는지 확인 위하여 실험을 진행하였다. HeLa 세포는 thymidine 1 mM, U2OS 세포는 thymidine 2.5 mM 로 각각 20 시간, 22 시간동안 처리하여 G1/S phase 에 arrest 시켰다. 그
다음 PBS 로 세척 후 HeLa 는 6 시간, U2OS 는 9 시간 후에 2-DG 6 mM, NaN3 10
mM 농도로 처리하여 세포내 ATP 와 mitotic index 를 측정하였다 (Figure 4A). 정상적인 조건에서는 세포주기가 진행될수록 ATP 의 level 이 증가했다가
감소하는 것을 볼 수 있었다. 반면에 2-DG 와 NaN3 를 처리 하였을 때는
ATP 가 증가하지 못하고 asynchronous 수준으로 떨어지는 것을 볼 수 있었다 (Figure 4B). 그 다음 mitotic index 를 측정한 결과 정상적인 조건에서는 mitosis 에 들어간 세포의 수는 증가했다가 감소하는 것을 볼 수 있었다. 반면에 ATP 결핍을 유도한 경우에는 mitosis 로 들어가지 못하고 세포주기가 정지되는 것을 볼 수 있었다 (Figure 4C).
12 0 1 2 3 4
Con 2+N Con 2+N Con 2+N Asyn 9h 11h 13h 15h R ati o of A T P l ev el 0 0.5 1 1.5 2 2.5 3 3.5
Con 2+N Con 2+N Con 2+N Asyn 6h 8h 10h 12h R ati o of A T P l ev el Seeding Thymidine treatment Thymidin e release 2-DG + NaN3 22h 9h 2h 2h 2h
U2OS
Seeding Thymidine treatment Thymidine release 2-DG + NaN3 20h 6h 2h 2h 2hHeLa
A
B
HeLa U2OS13
Figure 4. ATP depletion at G2 phase causes cell cycle arrest
(A) Scheme showing the experimental design. (B) Analysis of ATP level after release from thymidine block. (C) Cells were stained with aceto-orcein solution to count mitotic cells. (D) Scheme showing the experimental design. (E) Cells were stained with aceto-orcein solution to count mitotic cells. (F) Analysis of ATP level after release from thymidine block and drugs wash out.
0 2 4 6 8 10 12 14 16 0h 6h 8h 10h 12h Mito tic in d ex (%) 0 2 4 6 8 10 12 14 0h 9h 11h 13h 15h Mito tic in d ex (%) 2-DG + NaN3 HeLa U2OS 2-DG + NaN3
C
14
B. ATP 결핍에 의한 Cyclin B 의 핵으로의 이동억제 확인
Cyclin B 와 Cdk1 은 세포질에서 결합하여 핵 안으로 들어가 G2
phase 에서 Mitosis 로의 진행을 조절하는 단백질 복합체이다. 따라서 ICC(immuno cyto-chemistry) 를 통해 ATP 결핍이 세포 내에서 cyclin B 의 위치를 변화시키는지 확인하였다. 실험 결과 정상적인 세포에서는 cyclin B 가 핵으로 이동하여 mitosis 가 진행되는 것을 볼 수 있지만, ATP 가 결핍되었을 때는 핵 안으로 들어가지 못하고 세포질에 위치하여 있는 것을 볼 수 있었다 (Figure 5A). 핵으로 이동한 cyclin B 의 수를 측정한 결과 ATP 가 결핍 되면 핵으로 이동하지 못하는 것을 볼 수 있었다 (Figure 5B). 결과적으로 ATP 가 결핍되면 Cdk1-cyclin B 복합체가 핵 안으로 들어가지 못한다는 것을 알 수 있었다.
15
B
A
0h Control Cyclin B Lamin B DAPI Merge 8h 2-DG + NaN 3 Control 2-DG + NaN3 6h 10h ControlCyclin B Lamin B DAPI Merge
0h Control 11h 2-DG + NaN3 Control 2-DG + NaN 3 9h 2-DG + NaN3 12h Control 2-DG + NaN3 13h 15h
HeLa
U2OS
0 2 4 6 8 10 12 0h 9h 11h 13h 15h C y cl in B n u cl ea r tr an sl oc ati on (% ) Control 2-DG+NaN3 0 2 4 6 8 10 12 14 16 0h 6h 8h 10h 12h C y cl in B n u cl ea r tr an sl oc ati on (% ) Control 2-DG+NaN3HeLa
U2OS
2-DG + NaN3 2-DG + NaN316
Figure 5. ATP depletion inhibits nuclear translocation of Cyclin B
(A) Immunocytochemistry after release from thymidine block and ATP depletion. Cyclin B(red), Lamin B(green) and DAPI. (B) The nuclear translocation of cyclin B was analyzed by Immunocytochemistry in which we counted the percentage of cells showing an overlapped cyclin B among DAPI-positive cells.
17
C. Energy stress 에 의한 세포주기 정지 확인
그렇다면 이러한 현상이 정말로 ATP 에 의한 현상인지 알아보고자
하였다. 먼저 ATP 결핍을 유도하기 위해 사용한 2-DG 와 NaN3 를 wash
out 해준다음 mitotic index 와 ATP 를 측정하였다. HeLa 와 U2OS 에 thymidine 을 각각 1 mM, 2.5 mM 로 20 시간, 22 시간 처리 하여 G1/S phase 에 세포 주기를 정지 시킨 다음 PBS 세척을 통하여 thymidine 을 release 하였다. 이후 HeLa 는 6 시간,
U2OS 는 9 시간 후에 2-DG 와 NaN3를 처리하였다. 그 다음 2 시간 후에 PBS 로
세척을 한 후 normal media 로 갈아주었다 (Figure 6A). 그 결과 wash out 을 했을 때 ATP 가 다시 증가하는 것을 볼 수 있었다. 또한 세포주기가 다시 진행하는 것을 볼 수 있었다 (Figure 6B). 그 다음 세포내 ATP 를 측정한 결과 media change 를 했을 때 ATP 가 다시 정상 수준으로 올라가는 것을 볼 수 있었다 (Figure 6C).
2-DG 와 NaN3 가 ATP 의 생성을 막는 역할 외에 다른 경로를 통해 G2
arrest 를 일으킬 수도 있다고 예상하였다. 따라서 또 다른 energy stress 를 통해 G2-M 전이가 억제 되는지 확인하였다. ATP 의 생성은 glycolytic metabolism 에 의해 일어나기 때문에 G2 phase 에서 glucose 를 starvation 하고 mitotic index 확인하였다. HeLa 와 U2OS 에 thymidine 을 각각 1 mM, 2.5 mM 농도로 20 시간, 22 시간 처리 하여 G1/S phase 에 세포 주기를 정지 시킨 다음 PBS 세척을 통하여 thymidine 을 release 하였다. 이후 HeLa 는 6 시간, U2OS 는 9 시간 후에 normal media 에서 glucose free media 로 바꿔주었다 (Figure 6D). 그 결과 두 세포 모두 glucose 가 starvation 됐을 때 mitosis 로 잘들어가지 못하고 세포주기가 지연되는 것을 확인하였다 (Figure 6E). 그 다음 이러한 현상이 glucose 결핍으로 인한 것인지 알아보기 위해 glucose free media 를 처리한지 4 시간 후에 D-glucose 25 mM 을 다시 넣어주고 mitotic index 를 측정하였다. D-glucose 를 넣어주게 되면 삼투압이 증가된다고 알려져 있기 때문에 mannitol 을 negative control 로 사용하였다. 그 결과 D-glucose 를 다시 넣어주면 mitotic index 가 control 만큼
18
증가하는 것을 볼 수 있었다 (Figure 6E). 따라서 G2 phase 에서 glucose 의 결핍은 세포주기를 지연시킨다는 것을 알 수 있었다.
NaN3 는 mitochondrial cytochrome C oxidase 를 억제함으로써 ATP 생성을
막을 수 있지만 독성이 있어 사람에게 항암치료용 약물로는 사용할 수 없다.
따라서 당뇨병치료제로 알려졌고, 항암치료에도 사용되는 phenformin 을
사용하였다. Phenformin 은 mitochondrial complex I 을 억제한다고 알려져 있기 때문에 2-DG 와 함께 처리하면 ATP 생성을 억제할 수 있을 것이라고 예상하였다. HeLa 와 U2OS 에 thymidine 을 각각 1 mM, 2.5 mM 로 20 시간, 22 시간 처리 하여 G1/S phase 에 세포 주기를 정지 시킨 다음 PBS 세척을 통하여 thymidine 을 release 하였다. 이후 HeLa 는 6 시간, U2OS 는 9 시간 후에 2-DG 와 phenformin 을 처리하여 mitotic index 를 확인하였다 (Figure 6F). 그 결과 cell 들이 mitosis 로 들어가지 못하고 세포주기가 정지되는 것을 확인할 수 있었다 (Figure 6G).
그 다음 energy stress 를 가했을 때 세포내 ATP 를 확인하고자 하였다. 그 결과 2-DG 와 phenformin 을 처리한 세포에서 ATP 가 Asynchronous 수준으로 감소하는 것을 볼 수 있었다. 반면에 glucose starvation 조건에서는 ATP 가 많이 떨어지지 않는 것을 볼 수 있었다 (Figure 6H). 즉, 2-DG 와 phenformin 을 처리했을 때와 glucose starvation 을 했을 때 서로 다른 현상이 일어난 이유가 세포내 ATP 양의 감소정도가 다르기 때문이라는 것을 알 수 있었다. 결과적으로 G2 기에서 mitosis 로 진행하는데 있어 세포내 ATP 의 양이 중요하다는 것을 알 수 있었다.
19 Seeding Thymidine treatment Thymidine release 2-DG + NaN3 20h 6h 2h 2h 2h HeLa Wash out Seeding Thymidine treatment Thymidine release 2-DG + NaN3 22h 9h 2h 2h 2h U2OS Wash out
HeLa
0 5 10 15 0h 6h 8h 10h 12h M it o ti c in d ex ( %) Control 2-DG + NaN3 2-DG + NaN3 Wash outU2OS
0 5 10 15 0h 9h 11h 13h 15h M it o ti c in d ex ( %) Control 2-DG + NaN3 2-DG + NaN3 Wash out 0 1 2 3 4Con 2+N Con 2+N wash out Con 2+N wash out Asyn 0h 9h 11h 13h 15h R at io o f A T P l ev el 0 0.5 1 1.5 2 2.5 3 3.5
Con 2+N Con 2+N wash out Con 2+N wash out Asyn 0h 6h 8h 10h 12h R at io o f A T P l ev el HeLa U2OS
A
B
C
20
E
Seeding Thymidine treatment Thymidine release 2-DG + phenformin 20h 6h 2h 2h 2hHeLa
Seeding Thymidine treatment Thymidine release 22h 9h 2h 2h 2hU2OS
2-DG + phenforminF
D
Seeding Thymidine treatment Thymidine release Glucose starvation 20h 6h 2h 2h 2hHeLa
Seeding Thymidine treatment Thymidine release 22h 9h 2h 2h 2hU2OS
D-glucose or mannitol Glucose starvation D-glucose or mannitol 0 5 10 15 0h 6h 8h 10h 12h M It o ti c in d ex ( %) Control Glucose starvation D-glucose Mannitol 0 2 4 6 8 10 12 0h 9h 11h 13h 15h M it o ti c in d ex ( %) Control Glucose starvation D-glucose Mannitol HeLa U2OS21
Figure 6. Energy stress at G2 phase blocks mitotic entry
(A), (D), (F) Scheme showing the experimental design (B) Cells were stained with aceto-orcein solution to count mitotic cells. (C), (H) Analysis of ATP level after release from thymidine block. (E) Cells were stained with aceto-orcein solution to count mitotic cells. Cells treated with glucose-free media, D-glucose 25 mM and mannitol 25 mM, (G) Cells were stained with aceto-orcein solution to count mitotic cells. Cells treated 2-DG 6 mM, phenformin 100 μM. 0 5 10 15 20 25 0h 6h 8h 10h 12h M It o ti c in d ex ( %) Control 2-DG & phenformin 0 2 4 6 8 10 0h 9h 11h 13h 15h M it o ti c in d ex ( %) Control 2-DG & phenformin HeLa U2OS
G
HeLa U2OSH
0 0.5 1 1.5 2 2.5 3 3.5Con 2+P GS Con 2+P GS Con 2+P GS
Asyn 0h 6h 8h 10h 12h R ati o of A T P 0 0.5 1 1.5 2 2.5 3 3.5 4
Con 2+P GS Con 2+P GS Con 2+P GS
Asyn 0h 9h 11h 13h 15h R ati o of A T P
22
D. AMPK 가 G2/M transition 에 미치는 영향
G2 phase 에서 mitosis 로 넘어가기 위해서는 많은 양의 ATP 가 필요하다.
그런데 2-DG 와 NaN3 로 인해 ATP 의 생성이 억제되는 경우 세포 내에 AMP 의
농도가 높게 유지된다. AMP 가 AMPK 의 γ subunit 에 결합하게 되면 α subunit 의 구조적 변화가 일어나면서 LKB1 에 의해 인산화 되어 활성화된다. 따라서 AMPK 의 활성화로 인해 G2/M arrest 가 일어나는 것인지 알아보기 위해 실험을 수행하였다. HeLa 와 U2OS 에 thymidine 을 처리 하여 G1/S phase 에 세포 주기를 정지 시킨 다음 PBS 세척을 통하여 thymidine 을 release 하였다. 이후 HeLa 는 6 시간, U2OS 는 9 시간 후에 ATP 를 결핍 시켰다. ATP 를 결핍 시키고 2 시간 후에 AMPK 의 inhibitor 로 알려진 Compound C 를 HeLa 는 5 μM, U2OS 는 10 μM 농도로 처리하였다 (Figure 6A). 각 시간 별로 얻은 세포로 mitotic index 를 측정하였다. 그 결과 Compound C 를 단독으로 처리한 경우에는 정상적인 조건보다 세포주기가 지연되는 것을 볼 수 있었다. 그리고 ATP 가 결핍된 상황에서는 mitotic entry 가 일어나지 않았고, 이때 Compound C 를 처리해도 mitotic entry 는 일어나지 않았다 (Figure 6B). 그 다음엔 AMPK siRNA 를 통해 endogenous AMPK 를 knockdown 하고 mitotic index 를 확인하였다 (Figure 6C). 그 결과 AMPK 를 knockdown 했음에도 불구하고 ATP depletion 했을 때 mitotic entry 가 일어나지 않는 것을 확인하였다 (Figure 6D and 6E). 결과적으로 ATP 결핍으로 인해 활성화된 AMPK 를 억제해도 mitotic entry 는 일어나지 않았다. 따라서 ATP 결핍으로 인한 세포주기 정지가 AMPK 에 의한 것이 아니라는 것을 알 수 있었다.
23 Seeding
Thymidine
treatment Thymidine release
2-DG + NaN3 Compound C 20h 6h 2h 2h 2h HeLa Seeding Thymidine treatment Thymidine release 2-DG + NaN3 Compound C 22h 9h 2h 2h 2h U2OS
B
A
0 2 4 6 8 10 12 14 0h 6h 8h 10h 12h M it o ti c in d ex (% ) Control 2DG + NaN3 Compound C 2DG + NaN3 + Compound C 0 2 4 6 8 10 0h 9h 11h 13h 15h M it o ti c in d ex (% ) Control 2DG + NaN3 Compound C 2DG + NaN3 + Compound CHeLa
U2OS
2-DG + NaN3 Compound C 2-DG + NaN3 Compound C siAMPK Thymidinetreatment Thymidine release 2-DG + NaN3
20h 6h 2h 2h 2h
HeLa Seeding
siAMPK
Thymidine
treatment Thymidine release 2-DG + NaN 3
22h 9h 2h 2h 2h
U2OS Seeding
C
24
Figure 7. AMPK did not affect G2/M transition during ATP depletion
(A) Scheme showing the experimental design (B) Cells were stained with aceto-orcein solution to count mitotic cells (C) Scheme showing the experimental design (D) Cells were stained with aceto-orcein solution to count mitotic cells (E) Cells were lysed, and immunoblot analysis using indicated antibodies. Cells transfected with siRNA AMPKα1/2
D
0 5 10 15 20 25 0h 6h 8h 10h 12h M It o ti c in d ex (% ) Control 2+N siAMPK 2+N+siAMPKa 0 2 4 6 8 10 12 14 16 18 0h 9h 11h 13h 15h M it o ti c in d ex ( %) Control 2+N siAMPK 2+N+siAMPKaHeLa
U2OS
E
AMPKα1/2 GAPDH0h 6h Con2+NCon2+NCon2+NCon2+NCon2+NCon 2+N
siCon siAMPK siCon siAMPK siCon siAMPK
8h 10h 12h
0h 9h Con2+NCon2+NCon2+NCon2+NCon2+NCon2+N
siCon siAMPK siCon siAMPK siCon siAMPK
11h 13h 15h
25
E. DNA 손상 신호전달 경로에 의한 G2 phase arrest 확인
G2 phase 에서는 DNA 손상에 대한 checkpoint 가 존재하여 손상된 DNA 가 있을 경우 세포주기를 멈추고 손상된 DNA 를 고친 후에 세포주기를
진행시킨다. 따라서 ATP 를 결핍 시킬 때 사용되는 2-DG 와 NaN3로 인해 DNA
damage 가 생기는지 확인하고자 실험을 진행하였다. DNA damage 신호전달 경로에서 활성화되는 Chk1 의 인산화와 γ-H2AX 를 western blot 을 통해
확인하였다. Positive control 은 HeLa 세포에 H2O2 를 100μM 처리한 것을
사용하였다. 그 결과 2-DG 와 NaN3 를 처리해도 γ-H2AX 는 발현하지 않았다
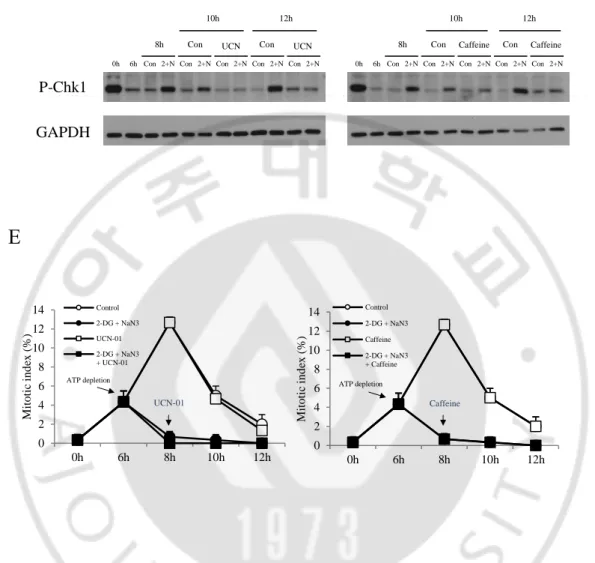
(Figure 7A). 그런데 ATP 를 결핍 시켰더니 DNA 가 손상되었을 때 활성화되는 Chk1 의 인산화가 증가하였다 (Figure 7B). 따라서 ATP 결핍에 의한 G2 phase arrest 는 DNA damage 에 의한 것이 아니라는 것을 알 수 있었다.
ATP 를 결핍 시켰을 때 DNA 의 손상은 없었지만 Chk1 의 인산화가 증가하는 것을 볼 수 있었다. 따라서 Chk1 의 활성으로 인해 G2 phase 에서 세포주기가 멈추는 것인지 알아보기 위해 실험을 진행하였다. HeLa 세포에 thymidine 을 처리 하여 G1/S phase 에 세포 주기를 정지 시킨 다음 PBS 세척을 통하여 thymidine 을 release 하였다. 이후 6 시간 후에 ATP 를 결핍 시켰다. ATP 를 결핍 시키고 2 시간 후에 Chk1 억제제인 UCN-01, Chk1/2 억제제인 Caffeine 을 처리하였다. 그 다음 각 시간 별로 얻은 세포로 mitotic index 와 western blot 을 수행하였다 (Figure 7C). Western blot 결과 각 inhibitor 들을 처리하였을 때 Chk1 의 인산화가 줄어드는 것을 볼 수 있었다 (Figure 7D). 그 다음 mitotic index 를 확인한 결과 Chk1 이 억제되었음에도 불구하고 mitosis 로 들어가지 못하는 것을 확인하였다 (Figure 7E). 따라서 ATP depletion 에 의한 Chk1 인산화의 증가는 세포주기가 멈추는 것과 관련 없다는 것을 알 수 있었다.
26
C
B
A
Seeding Thymidine treatment Thymidine release 2-DG + NaN3 20h 6h 2h 2h 2h Inhibitor HeLa P-Chk2(T68) P-Chk1(S345) GAPDH0h 6h Con2+NCon2+NCon2+N
8h 10 12h
0h 9h Con2+NCon2+NCon2+N
11h 13h 15h HeLa U2OS H2O2 100uM H2O2 100uM γ – H2AX
0h 6h Con2+NCon2+NCon2+N
8h 10h 12h
0h 9h Con2+NCon2+NCon2+N
11h 13h 15h H2O2 100uM H2O2 100uM HeLa U2OS
27
Figure 8. DNA damage did not affect G2/M transition during ATP depletion
(A~B) Each cell was collected at the indicated times after thymidine block and then cell lysates were immunoblotted with antibodies to the indicated proteins. (C) Scheme showing the experimental design (D) Each cell was collected at the indicated times after thymidine block and then cell lysates were immunoblotted with antibodies to the indicated proteins. (E) Cells were stained with aceto-orcein solution to count mitotic cells.
D
E
0 2 4 6 8 10 12 14 0h 6h 8h 10h 12h M it o ti c in d ex ( %) Control 2-DG + NaN3 UCN-01 2-DG + NaN3 + UCN-01 0 2 4 6 8 10 12 14 0h 6h 8h 10h 12h M it o ti c in d ex ( %) Control 2-DG + NaN3 Caffeine 2-DG + NaN3 + CaffeineATP depletion ATP depletion
Caffeine UCN-01
0h 6h Con2+NCon 2+NCon 2+NCon 2+NCon2+N
8h Con UCN Con UCN
10h 12h
0h 6h Con 2+NCon 2+NCon 2+NCon 2+NCon2+N
8h Con Caffeine Con Caffeine
10h 12h
P-Chk1 GAPDH
28 F. ATP 결핍이 Wee1 과 Cdc25C 에 미치는 영향 G2 phase 에서 mitosis 로 세포주기가 진행되기 위해서는 Cdk1 의 활성화가 필수적이다. 따라서 ATP 가 결핍되었을 때 Cdk1 의 활성에 영향을 미치는지 확인하고자 Cdk1 을 억제하는 tyrosine 15 번의 인산화를 확인해보았다. 그 결과 ATP 가 결핍되었을 때 Cdk1 의 인산화가 줄어들지 않고 그대로 유지되는 것을 볼 수 있었다. Cdk1 의 인산화는 Wee1 kinase 에 의해 일어나고 탈 인산화는 Cdc25C 에 의해 일어난다. 따라서 western blot 을 통해 Weee1 과 Cdc25C 의 protein level 을 확인해 본 결과 Wee1 과 Cdc25C 의 protein level 이 유지되는 것을 볼 수 있었다. 따라서 ATP 결핍이 Wee1 의 활성에 영향을 미치는지 알아보고자 하였다.
ATP 가 결핍되었을 때 Wee1 의 kinase activity 를 in vitro kinase assay 를 통해 확인 하고자 하였다. HeLa 세포에 thymidine 을 처리 하여 G1/S phase 에 세포 주기를 정지 시킨 다음 PBS 세척을 통하여 thymidine 을 release 하였다. 이후 6 시간 후에 ATP 를 결핍 시키고 2 시간 후에 세포를 얻어 IP 를 수행하였다. IP 후 invitro kinase assay 를하기위해 GST-Cdk1 을 넣어주고 invitro kinase assay 를 진행하였다. 그 결과 ATP 가 결핍 되었을 때 Wee1 의 activity 가 높은 것을 확인하였다.
29 Thymidine treatment Thymidine release 2-DG + NaN3 20h 6h 2h 2h 2h HeLa Seeding Thymidine treatment Thymidine release 2-DG + NaN3 22h 9h 2h 2h 2h U2OS Seeding
A
B
Wee1
Cdc25C
GAPDH
0h 6h Con 2+N Con 2+NCon 2+N
8h 10h 12h
0h 9h Con 2+N Con 2+NCon 2+N
11h 13h 15h
HeLa
U2OS
Seeding Thymidine treatment Thymidine release 20h 6h 2h IP 2-DG + NaN3 HeLaC
30
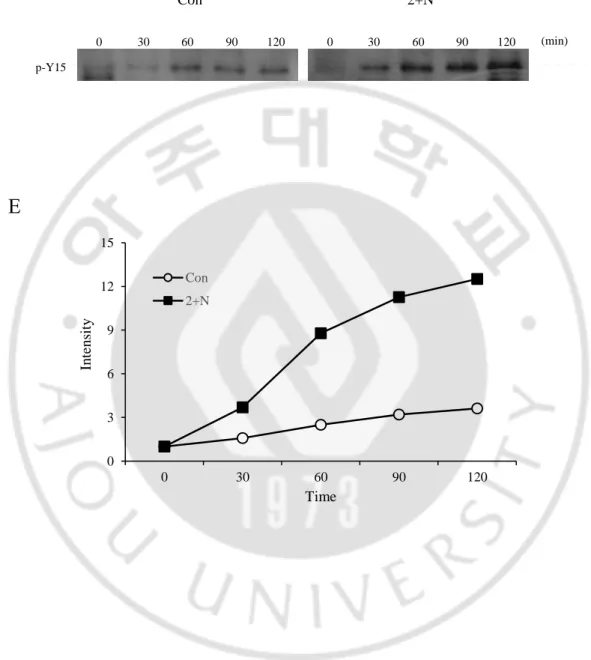
Figure 9. ATP depletion affect Cdk1 regulator
(A), (C) Scheme showing the experimental design (B) Each cell was collected at the indicated times after thymidine block and then cell lysates were immunoblotted with antibodies to the indicated proteins (D) Wee1 kinase assay (E) Band intensity of kinase assay
D
E
0 30 60 90 120 0 30 60 90 120 p-Y15 Con 2+N (min) 0 3 6 9 12 15 0 30 60 90 120 In te n sity Time Con 2+N31
G. Wee1 의 억제를 통한 G2-M transition 확인
In vitro kinase assay 결과 ATP 가 결핍되었을 때 Wee1 의 활성이 높았기 때문에 이를 억제할 수 있는 PD166285 를 처리하였을 때 G2/M transition 에 어떤 영향을 미치는지 알아보고자 실험을 진행하였다. HeLa 와 U2OS 에 thymidine 을 각각 1 mM, 2.5 mM 로 20 시간, 22 시간 처리 하여 G1/S phase 에 세포 주기를 정지 시킨 다음 PBS 세척을 통하여 thymidine 을 release 하였다. 이후 HeLa 는 6 시간, U2OS 는 9 시간 후에 ATP 를 결핍 시켰다. 그 다음 ATP 를 결핍 시키고 2 시간 후에 PD166285 를 0.5 μM 농도로 처리하였다. HeLa 는 6 시간, U2OS 는 9 시간째부터 2 시간 마다 세포를 얻어 mitotic index, western blotting 과 ICC 를 수행하였다 (Figure 8A). Western blotting 결과 P-Y15(Cdk1)도 줄어든 것을 확인하였다 (Figure 8B). Mitotic index 결과 두 세포 모두 PD166285 를 처리하였을 때 ATP 가 결핍되었음에도 불구하고 세포들이 mitosis 로 들어가는 것을 확인하였다 (Figure 8C). 그 다음 ICC 를 하여 cyclin B1 의 nuclear translocation 을 확인한 결과 PD166285 를 처리하면 ATP 를 결핍 시켰음에도 불구하고 핵으로 이동하는 것을 볼 수 있었다 (Figure 8D and 8E). 결과적으로 ATP 결핍으로 인한 G2 phase arrest 는 Wee1 에 의해 Cdk1 을 억제하는 인산화(P-Y15)가 유지 됨으로써 일어나는 현상임을 알 수 있었다.
32 Seeding Thymidine treatment Thymidine release 2-DG + NaN3 20h 6h 2h 2h 2h
HeLa
Seeding Thymidine treatment Thymidine release 2-DG + NaN3 22h 9h 2h 2h 2hU2OS
PD166285 PD166285A
B
HeLa
U2OS
0 5 10 15 20 25 30 35 0h 6h 8h 10h 12h M it o ti c in d ex (% ) Control 2-DG + NaN3 PD166285 2-DG + NaN3 + PD166285 2-DG + NaN3 PD166285 0 10 20 30 40 50 0h 9h 11h 13h 15h M it o ti c in d ex (% ) Control 2-DG + NaN3 PD166285 2-DG + NaN3 + PD166285 2-DG + NaN3 PD166285C
33
D
0 5 10 15 20 25 30 35 40 0h 6h 8h 10h 12h Nu cl ea r tr an sl oc ati on of C y cl in B ( % ) Control ATP depletion PD166285 ATP depletion + PD166285 0 10 20 30 40 50 0h 9h 11h 13h 15h Nu cl ea r tr an sl oc ati on of C y cl in B ( % ) Control ATP depletion PD166285 ATP depletion + PD166285 HeLa U2OSE
Cyclin BLamin B DAPI Merge
Control 2-DG + NaN 3 0h 6h 8h HeLa PD166285 PD + 2-DG + NaN3
Cyclin BLamin B DAPI Merge
Control 2-DG + NaN3 PD166285 PD + 2-DG + NaN3 Control 2-DG + NaN3 10h 12h U2OS Control 2-DG + NaN3 PD166285 PD + 2-DG + NaN3 Control 2-DG + NaN3 PD166285 PD + 2-DG + NaN3 Control 2-DG + NaN3 0h 9h 11h 13h 15h
34
Figure 10. PD166285 dramatically overcome G2 arrest by ATP depletion
(A) Scheme showing the experimental design. (B) Each cell was collected at the indicated times after thymidine block and then cell lysates were immunoblotted with antibodies to the indicated proteins. Cells treated with 6 mM 2-DG, 10 mM sodium azide and 0.5 μM PD166285. (C) Immunocytochemistry after release from thymidine block and ATP depletion. Cyclin B(red), Lamin B(green) and DAPI. (D) The nuclear translocation of cyclin B was analyzed by Immunocytochemistry in which we counted the percentage of cells showing an overlapped cyclin B among DAPI-positive cells.
35
H. Wee1 의 억제를 통해 mitosis 로 들어간 세포의 운명 확인
ATP 결핍된 상태에서 Wee1 을 억제하면 ATP 가 부족해도 mitosis 로 들어가는 것을 확인하였다. 따라서 ATP 를 결핍 시키고 Wee1 을 억제하여
mitosis 로 들어간 세포는 어떻게 되는지 알아보기 위해서 HeLa 세포를
이용하여 time-lapse 촬영을 하였다. 먼저 HeLa 에 thymidine 을 1 mM 로 20 시간 처리 하여 G1/S phase 에 세포 주기를 정지 시킨 다음 PBS 세척을 통하여
thymidine 을 release 하였다. 그 다음 6 시간 후에 2-DG 와 NaN3를 처리하였고, 그
다음 2 시간 후에 PD166285 를 처리하였다 (Figure 9A). Time-lapse 촬영 결과
ATP 를 결핍 시켰을 때 세포주기가 멈추는 것을 확인하였다. 그 다음
ATP 결핍으로 인해 세포주기가 멈춘 상태에서 PD166285 를 처리하게 되면 멈췄던 세포주기가 다시 진행되어 세포들이 mitosis 로 들어가는 것을 확인할 수 있었다. 그런데 mitosis 에 들어간 세포들을 관찰한 결과 G1 phase 로 빠지지 않고 mitosis 에 계속해서 머무는 것을 확인하였다 (Figure 9 B, C). Mitosis 에서 세포주기가 더 이상 진행되지 않을 경우 cell death 가 일어나기 때문에 trypan blue staining 을 이용하여 cell death 를 측정하였다. HeLa 와 U2OS 에 thymidine 을 각각 1 mM, 2.5 mM 로 20 시간, 22 시간 처리 하여 G1/S phase 에 세포 주기를 정지 시킨 다음 PBS 세척을 통하여 thymidine 을 release 하였다. 이후 HeLa 는 6 시간, U2OS 는 9 시간 후에 ATP 를 결핍 시켰다. 그 다음 ATP 를 결핍 시키고 2 시간 후에 PD166285 를 0.5 μM 로 처리하였다. 그 다음 HeLa 는 thymidine release 후 16 시간부터 24 시간 까지 4 시간 간격으로, U2OS 는 19 시간부터 27 시간 까지 4 시간 간격으로 cell death 를 측정하였다. 그 결과 Wee1 억제로 인해 mitosis 에 들어간 세포들이 ATP 결핍에 의해 죽는 세포 보다 더 많이 죽는 것을 관찰 할 수 있었다 (Figure 9D). 따라서 이러한 결과를 통해 G2 energy checkpoint 를 이용하여 암세포를 죽일 수 있는 새로운 항암화학요법을 제안한다.
36 Seeding Thymidine treatment Thymidine release 20h 6h 2h Wee1 inhibition 2-DG + NaN3
HeLa
A
B
Control ATP depletion PD166285 ATP depletion + PD1662857h
8h
9h
10h
11h
12h
37
D
C
2-DG + NaN 3 + PD166285 Control 2-DG + NaN 3 PD166285 8 9 10 11 12 (Hour) 8 9 10 11 12 (Hour) Mitosis Interphase * : p < 0.05 ** : p < 0.005 U2OS HeLa 0 5 10 15 20 25 30 35 40 Con 2-D G + N a N 3 P D 16628 5 2-D G + N a N 3+ P D Con 2-D G + N a N 3 P D 16628 5 2-D G + N a N 3+ P D Con 2-D G + N a N 3 P D 16628 5 2-D G + N a N 3+ P D 16h 20h 24h C el l de at h ( % ) ** ** * 0 5 10 15 20 25 Con 2-D G + N a N 3 P D 16628 5 2-D G + N a N 3+ P D Con 2-D G + N a N 3 P D 16628 5 2-D G + N a N 3+ P D Con 2-D G + N a N 3 P D 16628 5 2-D G + N a N 3+ P D 19h 23h 27h C el l de at h ( % ) ** * * * ** **38
Figure 11. To follow the cell fate after mitotic entry by PD166285
(A) Scheme showing the experimental design. (B) Time-lapse imaging of HeLa cell treated with 6 mM 2-DG, 10 mM sodium azide and 0.5 μM PD166285. The cell fate after mitotic entry by PD166285 (C) The cell fate after mitotic entry by PD166285 (D) Cells were stained with trypan blue to count dead cells.
39
IV. 고찰
본 연구에서는 ATP 결핍에 의한 세포주기 정지에 대한 현상확인과 기전규명에 초점을 맞추고 있다. ATP 의 양이 줄어들었음에도 불구하고 Cdk1 을 억제하는 인산화는 줄어들지 않았다. 이 인산화는 Wee1 과 Cdc25C 에 의해 조절된다. 본 연구에서는 Wee1 의 역할에 초점을 맞추고 실험을 진행하였다. Western blot 을 통해 Cdc25C 의 protein level 을 확인하였을 때 ATP 결핍시에 protein level 이 감소되는 것처럼 보였다. 이에 대해 protein degradation 의발생인지 확인하고자 protease inhibitor 인 MG132 를 처리하여 실험을
진행하였지만 세포주기가 진행되지 않았다. 따라서 ATP 결핍시에 Cdc25C 의 역할에 대한 추가적인 실험이 필요하다.
본 연구는 암세포의 G2 phase 에서 energy stress 가 세포주기를 멈춘다는 것을 보여준다. 세포주기가 멈추는 원인에는 Cdk1 의 활성이 감소되는 것과 cyclin B1 의 핵으로의 이동이 억제된 것으로 생각된다. 이러한 현상은 세포의 종류와 상관없이 다양한 암세포에서 보여진다.
NaN3 는 독성이 강한 약물로써 실험적으로 ATP 결핍을 유도하기 위해
사용하였다. 따라서 임상에서 사용할 수 있으며 비슷하게 mitochondria 의 활성을 억제할 수 있는 phenformin 을 사용하였다 (Wheaton et al., 2014). 그리고 실제 암이 발달하는 과정에서 필연적으로 energy stress 를 받을 수 밖에 없다. 따라서 이를 모방하여 glucose starvation 조건을 추가적으로 실험하였다. 2-DG 와 phenformin 을 처리했을 때 ATP level 이 asynchronous 수준으로 감소하고 세포주기가 멈추는 것을 확인하였다. 반면에 Glucose starvation 조건에서는 drugs 를 처리했을 때 보다는 ATP level 감소가 약한 정도였고 세포분열기로의 이행이 지연되는 것을 볼 수 있었다. 암세포에는 특이적으로 알코올 분해효소(ALDH)가 많이 존재한다고 밝혀졌는데, 암세포는 이를 이용하여
40
NADH 를 생산하여 ATP 를 생성할 수 있다고 알려져 있다 (Kang et al., 2016). 따라서 glucose starvation 상황에서 ATP 생성에 대한 연구가 필요하다. 결과적으로 세포분열기로의 이행에 있어서 일정 수준의 ATP 가 필요하다는 것을 나타낸다.
따라서 energy stress 에 의해 세포주기가 멈추는 원인을 찾고자 하였다. 세포내 ATP 가 줄어들게 되면 AMPK 가 활성화 되기 때문에 AMPK 의 억제제를 처리하고, siRNA 를 이용하여 knock down 을 하였지만 세포주기가 멈춘 것을 회복시킬 수 없었다. 따라서 G1 phase energy stress 에서는 AMPK 의 역할이 중요하지만 G2 energy checkpoint 에서는 큰 역할을 하지 않는 것으로 생각된다. ATP 의 결핍을 유도하는데 사용하는 약물로 인한 DNA 손상이 없었음에도 불구하고 DNA damage signaling pathway 인 Chk1 의 인산화가 증가하는 것을 확인하였다. 따라서 UCN-01 과 caffeine 을 처리하여 Chk1 의 인산화를 억제하였지만 세포주기가 멈춘 것을 회복시킬 수 없었다. 그리고 0h 에서 Chk1 의 인산화가 증가하였는데 이는 thymidine 에 의한 replication stress 에 의한 현상이라고 생각된다 (Feijoo et al., 2001). 따라서 ATP 가 결핍되었을 때 Chk1 의 인산화에 대한 추가적인 연구가 필요하다.
ATP 가 결핍되었음에도 불구하고 Cdk1 을 억제하는 인산화는 줄어들지 않았다. 따라서 Wee1 을 억제하는 PD166285 를 처리하였을 때 멈췄던 세포주기가 진행하는 것을 볼 수 있었다. 하지만 mitosis 에서 더 이상 세포주기가 진행되지 않고 세포주기가 멈추는 것을 확인하였다. 이는 mitosis 때 Cdk1-cyclin B 의 활성이 줄어들어 mitotic exit 이 일어나야 하지만 Wee1 의 억제에 의해 Cdk1 의 활성이 줄어들지 않아 일어나는 현상으로 생각된다. 그 근거로 Wee1 inhibitor 단독으로 처리된 조건에서 cytokinesis 가 일어나지 않는 것을 ICC 를 통해 관찰하였다.
41
PD166285 의 경우 Wee1 특이적 억제제는 아니다. 현재 임상에서 쓰이고 있는 MK-1775 라는 Wee1 특이적인 약물로도 같은 효과를 보이는지 추가적인 연구가 필요하다.
Energy stress 에 의해 세포주기가 멈춘 상태에서 Wee1 억제에 의한
세포주기의 진행은 분열 효모에서는 lifespan 을 증가시켰지만 (Masuda et al., 2016), 암세포에서는 오히려 정 반대의 결과를 초래하였다. 따라서 이를 이용한 항암화학요법을 기대할 수 있다.
방사선 요법이나 여러 항암제들은 세포내에서 DNA 손상을 일으켜 세포를 죽이는 방법으로 다양한 종양을 치료하는데 사용되어져 왔기 때문에 이를 이용한 치료 효율의 향상이 요구 되어왔다 (Indovina and Giordano, 2010). Wee1 은 DNA 손상에 대한 반응으로 G2 checkpoint 를 조절하기 때문에 Wee1 의 억제는 DNA 복구를 하지 못하게 함으로써 치료 효율을 향상 시킬 수 있다. 방사선 치료 외에도 gemcitabine 이나 5-FU 와 Wee1 억제제인 MK-1775 와 함께 사용하면 항암효과를 증대 시킬 수 있다 (Hirai et al., 2010; Bridges et al., 2011; Kreahling et al., 2013). 이러한 결과로 인해 MK-1775 에 대한 임상실험도 진행 중이다 (Do et al., 2015).
42
V. 결론
본 연구에서는 G2 phase 에서 ATP 결핍이 G2-M transition 에 어떤 영향을
미치는지 알아보고자 하였다. 따라서 G2 phase 에서 2-DG 와 NaN3 를 처리하여
ATP 생성을 억제하고 mitotic index 를 확인하였다. 그 결과 ATP 생성을 억제하자 G2 phase 에서 세포주기가 멈추는 것을 확인하였다. 이러한 현상이 정말로
ATP 의 감소에 의한 것인지 알아보고자 ATP 결핍 이후에 2-DG 와 NaN3 를
wash out 하였다. 그 결과 멈췄던 세포주기가 다시 정상적으로 회복하는 것을 확인하였다. Cdk1-cyclin B 복합체는 G2 phase 에서 핵으로 들어가 G2-M transition 을 촉진한다. 따라서 ATP 결핍이 Cdk1-cyclin B 복합체의 핵으로의 이동에 영향을 미치는지 알아보고자 ICC 를 통해 확인하였다. 그 결과 ATP 를 결핍 시키면 cyclin B 가 핵으로 들어가지 못하고 세포질에 머무는 것을 확인하였다.
이러한 현상이 정말로 ATP level 의 감소에 의한 것인지 알아보고자 또 다른 energy stress 를 가해주었다. 먼저 glucose starvation 상태에서도 G2 arrest 가
일어나는지 알아보기 위하여 Thymidine release 후 2-DG 와 NaN3 대신에 glucose
free media 로 media change 를 해주었다. 그 결과 세포주기가 완전히 멈추지는
않고 지연되는 것을 확인하였다. 그리고 NaN3 대신 phenformin 을 사용했을
때에도 G2 arrest 가 일어나는지 확인하고자 Thymidine release 후 2-DG 와 phenformin 을 처리하였다. 그 결과 세포주기가 멈추는 것을 확인하였다. Energy stress 를 가해주고 ATP level 을 측정한 결과 glucose starvation 상황에서는 조금 감소하였고, 2-DG 와 phenformin 을 처리하였을 때는 asynchronous 수준으로 감소하는 것을 볼 수 있었다.
결과적으로 ATP 결핍은 G2-M transition 을 억제한다는 것을 확인하였다. 그렇다면 이러한 현상이 AMPK 에 의해 일어나는 것인지 알아보고자 AMPK siRNA 와 AMPK inhibitor 인 Compound C 를 처리하여 mitotic index 를 확인하였다.
43
그 결과 AMPK 를 억제했음에도 불구하고 ATP 를 결핍 시켰을 때 세포주기가 정지되어 있는 것을 확인하였다. 따라서 ATP 결핍으로 인한 G2 phase arrest 는
AMPK 에 의한 것이 아니라는 것을 확인하였다. 그렇다면 2-DG 와 NaN3 의
처리로 인해 DNA damage 가 발생하여 세포주기가 멈추는 것인지 알아보고자 Western blot 을 통해 확인하였다. 그 결과 γ-H2AX 는 발현하지 않았지만 DNA damage signaling pathway 인 Chk1 이 인산화 된 것을 확인하였다. 따라서 inhibitor 를 사용하여 Chk1 의 인산화를 억제하였지만 ATP 결핍으로 인한 G2 arrest 를 회복시킬 수 없었다. 따라서 ATP 결핍으로 인한 Chk1 의 인산화는 G2 arrest 와 관련이 없다는 것을 알 수 있었다. Western blot 을 통하여 Cdk1 의 억제 인산화를 확인한 결과 ATP 가 결핍되었을 때 Cdk1 의 인산화가 줄어들지 않고 유지되는 것을 확인하였다. 이 인산화는 Wee1 에 의해 인산화 되는 것으로 알려졌기 때문에 Wee1 inhibitor 인 PD166285 를 처리하고 western blot 과 mitotic index 를 측정하였다. 그 결과 Cdk1 의 인산화(Y-15)가 감소하였고 ATP 가 결핍되었음에도 불구하고 G2 phase 에서 mitosis 로 진행되는 것을 확인하였다. 같은 방법으로 ICC 를 수행한 결과 ATP 가 결핍 되었음에도 Wee1 inhibitor 를 처리하게 되면 Cyclin B 가 핵으로 이동할 수 있는 것을 확인하였다. 이러한 현상을 HeLa 세포를 사용하여 Time lapse 를 통해 관찰하였다. 그 결과 PD166285 로 인해 대부분의 세포들이 mitosis 에 들어가는 것을 관찰하였다. 그런데 이렇게 mitosis 에 들어간 세포들은 G1 phase 로 넘어가지 않고 계속해서 mitosis 에 머물며 죽는 것처럼 보였다. 따라서 cell death 를 측정하였다. 그 결과 ATP 가 결핍된 상태에서 PD166285 로 인해 mitosis 에 들어가게 되면 많은 세포가 죽는 것을 확인하였다. 즉, G2 phase 에서 energy stress 가 발생하면 세포주기가 멈추고 이러한 현상을 표적으로 하는 항암치료를 기대할 수 있다.
44
참고문헌
1. Arellano M, Moreno S: Regulation of CDK/cyclin complexes during the cell cycle.
The international journal of biochemistry & cell biology 29: 559-573, 1997 2. Bartek J, Lukas J: Mammalian G1-and S-phase checkpoints in response to DNA
damage. Current opinion in cell biology 13: 738-747, 2001a
3. Bartek J, Lukas J: Pathways governing G1/S transition and their response to DNA damage. FEBS letters 490: 117-122, 2001b
4. Bhowmick NA, Ghiassi M, Aakre M, Brown K, Singh V, Moses HL: TGF-β-induced RhoA and p160ROCK activation is involved in the inhibition of Cdc25A with resultant cell-cycle arrest. Proceedings of the National Academy of Sciences 100: 15548-15553, 2003
5. Boxem M, van den Heuvel S: lin-35 Rb and cki-1 Cip/Kip cooperate in developmental regulation of G1 progression in C. elegans. Development 128: 4349-4359, 2001
6. Bridges KA, Hirai H, Buser CA, Brooks C, Liu H, Buchholz TA, Molkentine JM, Mason KA, Meyn RE: MK-1775, a novel Wee1 kinase inhibitor, radiosensitizes p53-defective human tumor cells. Clinical cancer research 17: 5638-5648, 2011 7. Cahill DP, Lengauer C, Yu J, Riggins GJ, Willson JK, Markowitz SD, Kinzler KW,
Vogelstein B: Mutations of mitotic checkpoint genes in human cancers. Nature
392: 300-303, 1998
8. Carling D, Sanders MJ, Woods A: The regulation of AMP-activated protein kinase by upstream kinases. Int J Obes (Lond) 32 Suppl 4: S55-59, 2008
9. Chignola R, Milotti E: A phenomenological approach to the simulation of metabolism and proliferation dynamics of large tumour cell populations. Phys Biol 2: 8-22, 2005
10. Do K, Wilsker D, Ji J, Zlott J, Freshwater T, Kinders RJ, Collins J, Chen AP, Doroshow JH, Kummar S: Phase I study of single-agent AZD1775 (MK-1775), a Wee1 kinase inhibitor, in patients with refractory solid tumors. Journal of Clinical Oncology 33: 3409-3415, 2015
45
DNA-PKcs to sites of DNA damage. Nature 434: 605-611, 2005
12. Feijoo C, Hall-Jackson C, Wu R, Jenkins D, Leitch J, Gilbert DM, Smythe C: Activation of mammalian Chk1 during DNA replication arrest. The Journal of cell biology 154: 913-924, 2001
13. Finkel T, Hwang PM: The Krebs cycle meets the cell cycle: Mitochondria and the G1–S transition. Proceedings of the National Academy of Sciences 106: 11825-11826, 2009
14. Hardie DG, Ross FA, Hawley SA: AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat Rev Mol Cell Biol 13: 251-262, 2012
15. Hermeking H, Lengauer C, Polyak K, He T-C, Zhang L, Thiagalingam S, Kinzler KW, Vogelstein B: 14-3-3σIs a p53-Regulated Inhibitor of G2/M Progression. Molecular cell 1: 3-11, 1997
16. Hirai H, Arai T, Okada M, Nishibata T, Kobayashi M, Sakai N, Imagaki K, Ohtani J, Sakai T, Yoshizumi T: MK-1775, a small molecule Wee1 inhibitor, enhances anti-tumor efficacy of various DNA-damaging agents, including 5-fluorouracil. Cancer biology & therapy 9: 514-522, 2010
17. Hurford R, Cobrinik D, Lee M-H, Dyson N: pRB and p107/p130 are required for the regulated expression of different sets of E2F responsive genes. Genes & development 11: 1447-1463, 1997
18. Indovina P, Giordano A: Targeting the checkpoint kinase WEE1: selective sensitization of cancer cells to DNA-damaging drugs. Cancer biology & therapy
9: 523-525, 2010
19. Kang JH, Lee S-H, Hong D, Lee J-S, Ahn H-S, Ahn J-H, Seong TW, Lee C-H, Jang H, Hong KM: Aldehyde dehydrogenase is used by cancer cells for energy metabolism. Experimental & molecular medicine 48: e272-e272, 2016
20. Kreahling JM, Foroutan P, Reed D, Martinez G, Razabdouski T, Bui MM, Raghavan M, Letson D, Gillies RJ, Altiok S: Wee1 inhibition by MK-1775 leads to tumor inhibition and enhances efficacy of gemcitabine in human sarcomas. PloS one 8: e57523, 2013
21. Lindqvist A, Rodríguez-Bravo V, Medema RH: The decision to enter mitosis: feedback and redundancy in the mitotic entry network. The Journal of cell biology
185: 193-202, 2009