저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게 l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다: l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다. l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다. 저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다. Disclaimer 저작자표시. 귀하는 원저작자를 표시하여야 합니다. 비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다. 변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
Doct
or
alThesi
si
n
t
heDepar
t
mentofBi
omedi
calSci
ences
St
udy on t
hePhysi
ol
ogi
cal
Funct
i
on ofTRADD
i
n TRAI
L-Si
gnal
i
ng and
DNA DamageResponse
Aj
ou Uni
ver
si
t
y Gr
aduat
eSchool
Maj
ori
n Mol
ecul
arMedi
ci
ne
St
udy on t
hePhysi
ol
ogi
cal
Funct
i
on ofTRADD
i
n TRAI
L-Si
gnal
i
ng and
DNA DamageResponse
You-Sun Ki
m,Advi
sor
Isubmi
tt
hi
st
hesi
sast
heDoct
or
alt
hesi
s
i
n t
heDepar
t
mentofBi
omedi
calSci
ences.
12 2016
Aj
ou Uni
ver
si
t
y Gr
aduat
eSchool
Maj
ori
n Mol
ecul
arMedi
ci
ne
Gi
-Bang Koo
- ABSTRACT-
Study on the Physiological Function of TRADD
in TRAIL-Signaling and DNA Damage Response
TNF Receptor-Associated Death Domain (TRADD) is an essential mediator of TNF receptor1 (TNFR1)-mediated TNF signaling. TRADD has a critical function in TNFR1 signaling pathway by orchestrating the formation of the TNFR1 signaling complex as well as Toll-like receptor3 (TLR3) and TLR4 signaling. The cytoplasmic functions of TRADD in death receptor signaling have been intensively studied. However, the involvement of TRADD in TNF-related apoptosis-inducing ligand (TRAIL) - and DNA damage-induced cell death is unclear. In this study, I investigated the role of TRADD in cell death induced under these two conditions. TRAIL is known to possess selective cytotoxicity towards cancer cells and therefore is considered as a promising anti-cancer therapeutic agent. One major obstacle in the clinical application of TRAIL as a cancer therapeutic agent is the acquisition of TRAIL resistance. Therefore, it is important to search for novel mechanisms underlying such resistance. In this study, I found that TRADD deficiency sensitizes cells to TRAIL-induced apoptosis. Enhanced cell death in TRADD-/- MEFs is associated with defective NF- B activation,
indicating that the pro-survival function of TRADD in TRAIL signaling is mediated at least in part via NF- B activation. Consistently, siRNA knock -down
of TRADD in cancer cells sensitizes them to TRAIL-induced apoptosis. Thus, our data clearly demonstrate the pro-survival role of TRADD in TRAIL signaling and targeting TRADD appears to be a logical approach in enhancing the killing effect of TRAIL and in overcoming TRAIL resistance in cancer therapy. On the other hand, it has been found that TRADD shuttles dynamically from the cytoplasm into the nucleus through its nuclear localization sequence (NLS) and nuclear export sequence (NES). However, the function of nuclear localization of TRADD remains elusive. In this study, I aimed to examine the physiological functions of TRADD in nucleus upon treatment with DNA damage agents. First, I found that TRADD deficiency increases the accumulation of double strand break (γH2AX) foci compared to wild type in response to DNA damage agent treatment. Second, I identified that DNA damage promotes translocation of TRADD from cytoplasm to the nucleus. Third, I found that nuclear localization of TRADD facilitates DNA repair through promotion of non homologous end-joining (NHEJ) repair rather than homologous recombination (HR) repair. Finally, I demonstrated that TRADD deficiency is able to sensitize MEFs to DNA damage-induced cell death through the persistent activation of JNK and accumulation of ROS. Taken together, data from this study collectively demonstrate that TRADD has a pro-survival role in TRAIL- and DNA damage- induced cell death. Thus, TRADD may be one of the key potential target for overcoming TRAIL resistance and DNA damage resistance in cancer therapy.
. Keywords: TRADD, TNFR1 signaling, TRAIL, DNA Damage Response, Cancer Cell Death
TABLE OF CONTENTS
ABSTARCT ···
iTABLE OF CONTENTS ···
ivLIST OF FIGURES ···
viiI. INTRODUCTION ···
1II. MATERIALS AND METHODS ···
6A. Cell culture ··· 6
B. Reagents and Antibodies ··· 6
C. Western blot ··· 7 D. Cytotoxicity Assay ··· 7 E. Transfection ··· 7 F. Flow cytometry ··· 8 G. RNA Interference ··· 8 H. Immunofluorescence ··· 9 I. Immunoprecipitation ··· 9
J. Pull Down Assay ··· 9
K. HR- or NHEJ-mediated DSB repair GFP reporter systems ··· 10
L. Nucleus and cytoplasm fractionation ··· 10
M. Laser microirradiation··· 10
N. FokI assay ··· 11
III. RESULTS ···
12A. TRADD is critical for resistance to TRAIL-induced cell death through NF-κB activation ··· 12
1. Deficiency of TRADD sensitizes cells to TRAIL-induced cell death in MEFs 12 2. TRADD reconstitution suppressed TRAIL-induced apoptosis ··· 16
3. TRAIL-induced NF-κB activation has pro-survival function in MEF cells ··· 18
4. Impaired JNK and ERK activation by TRAIL in TRADD-/-MEFs ··· 21
5. Knockdown of TRADD sensitizes cancer cells to TRAIL-induced cell death · 24 6. TRAIL-induced DISC complex formation ··· 26
B. Nuclear TRADD has a Protective Function During DNA Damage Responses ··· 28
1.TRADD is involved in the Hydrogen Peroxide-Induced DNA Damage Response ··· 28
2. Depletion of TRADD sensitizes to DNA-Damage··· 33
3. TRADD translocates to the nucleus following DNA damage..··· 36
4. Nuclear localization of TRADD required for DNA repair ··· 39
5. TRADD is required for non-homologous end joining pathway. ··· 42
6. Deletion of TRADD sensitizes cells to DNA-damaging agent ··· 47
7. Deficiency of TRADD Leads to ROS accumulation and prolonged JNK activation ··· 51
8. Proposed model ··· 55
V. CONCLUSION ··· 60
Ⅵ. REFERENCES ···
61LIST OF FIGURES
Figure. 1. Deletion of TRADD sensitizes cells to TRAIL-induced cell death ··· 13
Figure. 2. TRADD deficiency sensitizes cells to apoptosis induced by TRAIL but not TNFa ··· 14
Figure. 3. Reconstitution of TRADD in TRADD-/-MEFs blocks TRAIL-induced Cell Death ··· 17
Figure. 4. TRAIL-Induced NF-kB activation is impaired in TRADD-/-MEFs ··· 19
Figure. 5. TRAIL-induced JNK and ERK activation are impaired in TRADD-/-MEFs, but do not contribute to cell death sensitivity ··· 22
Figure. 6. Depletion of TRADD can sensitize cancer cell to TRAIL-induced cell death · 25 Figure. 7. DISC formation was detected by GST-pull down assay ··· 27
Figure. 1. Deficiency of TRADD induced impaired DNA Damage Response ··· 30
Figure. 2. TRADD regulates DNA repair following DNA-damage agent treatment ··· 34
Figure. 3. DNA-Damage induces nuclear translocation of TRADD. ··· 37
Figure. 4. Nuclear localization of TRADD required for DNA repair ··· 40
Figure. 5. TRADD is required for non-homologous end joining repair ··· 44
Figure. 6. Deletion of TRADD sensitizes to DNA-Damage agent induced cell death ··· 49
Figure. 7. TRADD Induces ROS accumulation and prolonged JNK activation ··· 53
I. INTRODUCTION
TNF receptor associated death domain (TRADD) was identified as the first component in the TNF receptor 1(TNFR1) signaling pathway and it is believed to be a critical mediator of TNFR1-mediated TNF signaling. Engagement of TNFR1 by the TNF homotrimer initiates the binding of the adaptor protein, which then recruits other effector proteins, such as Receptor interacting protein 1 (RIP1) and TNFR-associated factor 2 (TRAF2) to form a TNFR1 signaling complex leading to the activation of several pathways, including NF-kB and MAP kinase (Hsu H et al., 1995; Chen et al., 2002; Wajant et al., 2003). Under certain conditions, it is thought that the TRADD, TRAF2, and RIP1 proteins dissociates from the receptor and form a secondary protein complex containing other proteins such as FADD and caspase 8 leading to apoptosis (Micheau et al., 2003). In addition to TNFR1, TRADD may mediate signaling downstream of the death receptors DR3 and DR6. However, because of the lack of the TRADD-deficient mice model, the physiological function of TRADD has remained unclear and controversial. Kim YS and Ermolaeva generates TRADD knockout mice and found that TRADD has a critical function in TNFR1 signaling pathway by orchestrating the formation of the TNFR1 signaling complex as well as TLR3 and TLR4 signaling (Kim YS et al., 2008; Ermolaeva MA et al., 2008). Deficiency of TRADD abrogated TNF-induced apoptosis and necrosis, abolished both NF-κB and MAP kinase activation. TRADD
was also required for TRIF-dependent TLR signaling in mouse embryonic fibroblasts but not macrophages suggesting TRADD is an essential component of TNFR1 signaling pathway and has a critical but apparently cell type-specific function in TRIF-dependent TLR signaling (Kim YS et al., 2008).
Despite its role in mediating TNFR1 signaling, many studies suggested that TRADD might also play a role in the response to viral infection. Using the TRADD knockout mice, Michallet and colleagues reported that the TRADD functions as an important component of the antiviral signaling pathway in response to infection by RNA viruses showing deficiency of TRADD prevented Cardif-dependent activation of IFN, reduced the production of IFN in response to RNA viruses (Michallet MC et al., 2008).
The cytoplasmic function of TRADD in death receptor signaling have been intensively studied (Yelena L et al., 2012) (Table. TRADD is necessary; -, TRDD is not involved; NA, not applicable.).
TRADD in Death Receptor signaling
Although TRADD is usually studied on function of a cytoplasmic protein, it has been known that TRADD is shuttling protein from the cytoplasm into the
nucleus due to the their Nuclear Export Signal (NES) at amino acid 147-163 and Nuclear Localization Signal (NLS) at amino acid 229-242, which allow shuttling between the nucleus and the cytoplasm (Michael Morgan et al., 2002; Bender LM
et al., 2005).
The domain structure of TRADD
Recently, it was reported that nuclear localization of TRADD contributes to tumor suppression by regulating ULF-dependent p19Arf ubiquitylation in skin cancer mice model (Chio II et al., 2012). In this report, TRADD shuttles dynamically from the cytosol into the nucleus to promote p19Arf protein stability and tumor suppression. TRADD-deficient mice exhibit enhanced tumor formation in DMBA/TPA-induced skin carcinogenesis. TRADD also has reported that it is commonly expressed at high levels in Glioblastoma Multiforme (GBM) and is detected in both cytoplasm and nuclear distribution (Chakraborty S et al., 2013). Silencing of TRADD in glioma cells resulted in increased sensitivity to temozolomide (TMZ) by regulating NF-kB suggesting that cytoplasmic TRADD is
a key driver of NF-kB activation in GBM. Despite the fact that TRADD was detected in nucleus, the function of nuclear localization of TRADD still remain unclear.
In this study, we show that TRADD deficiency renders cells more susceptible to TRAIL-induced cell death. This is likely due to impairment of the pro-survival NF-kB activation since TRAIL does not induce NF-kB activation in TRADD
-/-MEFs, while ectopic expression of p65 or IKKβ prevents TRAIL-induced cell death in these cells. Despite the fact that TRADD-/- MEFs lack TRAIL-induced ERK and JNK activation, the loss of these pathways does not contribute to TRAIL-induced cell death in the knockout MEFs.
Although TRADD has been investigated intensively in cytoplasmic function, much less is known about its function in nucleus. Therefore, we investigated that the physiological function of TRADD in nucleus. Here we show that TRADD translocates into the nucleus to regulate the DNA damage repair. Upon DNA damage, TRADD moves to the nucleus and modulates Non-Homologous End-Joining (NHEJ) pathway. Under the DNA damage, deficiency of TRADD causes increased ROS and persistent activation of JNK, which leading to cell death.
Taken together, our data suggest that TRADD is potential target for cancer cell death in TRAIL- and DNA damage-induced cell death. Thus, The regulation of TRADD expression in cancer serve as a target for sensitizing cancer cells to TRAIL- and DNA damage-induced cytotoxicity.
II. MATERIALS AND METHODS
A. Cell culture
TRADD-/- mouse embryonic fibroblast (MEF) cells and their culture conditions
have been previously described (Kim YS et al., 2008). Murine stable cell lines were established by transfected with WT TRADD, NES mutant TRADD, and Src-TRADD and then selected in media containing G418/Neomycin. MEF, HeLa, HepG2, U2O2 2-6-3, and U2OS cell were cultured in DMEM with 10% FBS, 2 mM glutamine, 100 U/ml penicillin, and 100 mg/ml streptomycin. Wild type and
TRADD-/- MEFs (Kims YS et al., 2008; Chen, G et al., 2002; Hsu H et al., 1996;
Hsu H et al., 1995) and HepG2 cells TRADD-/- (Flag-mTRADD) stable cell lines
were selected in media containing 2 μg/ml puromycin.
B. Reagents and Antibodies
Hydrogen Peroxide (H2O2), Doxorubicin (Doxo), Etoposide (Etopo), Camptothecin (Cpt), Cisplatin (CDDP), Hydroxyurea (Hu), NAC, and H2DCF-DA were purchased from Sigma Aldrich. Recombinant murine TRAIL and TNFα were from R&D. Cycloheximide (CHX), SP 600125 (SP), PD 98059 (PD), SB 202190 (SB), and IKK-2 inhibitor were from Calbiochem.
γH2AX(S139), phospho-ERK, phospho-p38, phospho-IκBα, ERK, p38, Caspase 3, Caspase 8, 53BP1 and PARP1 antibodies were purchased from Cell
signaling. Actin, mouse TRADD, Lamin B, FADD, IκBα, RAD51, Myc and Sp1 antibodies were purchased from Santa cruz. . Hsp90 antibody were purchased from Abcam. Human TRADD antibody were purchased from millipore. Phospho-JNK, and JNK were purchased from invitrogen. TRAIL-R, RIP1 and RIP1 antibodies were purchased from BD transduction. Glutathione S-transferase (GST)-TRAIL was described previously (Lin Y et al., 2000)
C. Western blot
Cells were lysed in M2 buffer (20 mM Tris at pH 7, 0.5% NP-40, 250 mM NaCl, 3 mM EDTA, 3 mM EGTA, 2 mM DTT, 0.5 mM PMSF, 20 mM β-glycerol phosphate, 1 mM sodium vanadate, and 1 μg ml−1 leupeptin). Equal amounts of cell extracts were resolved by SDS-PAGE and analyzed by immunoblotting.
D. Cytotoxicity Assay
Cell viability was determined using tetrazolium dye colorimetric test (MTT assays with absorbance read at 570 nm ). Cell survival assays were performed using the Cell Titer-Glo Luminescent Cell Viability Assay kit (Promega, G7570) according to manufacturer’s instruction.
E. Transfection
Wild type-TRADD, NES mutant TRADD, and Src-TRADD and GFP-p65 or Mock vector with Lipofectamine PLUS reagent according to manufacturer's protocol (Invitrogen, Inc.). For generation of mutant TRADD expressing cell line in
TRADD-/- MEFs, trasnfected cell selected with G418 at 7days.
F. Flow cytometry
To detect intracellular ROS, we incubated cells treated with 10 μM H2 DCFDA for 30 minutes before the end of the indicated drug treatments and the increase in fluorescence was measured by FACS Aria III (BD Biosciences, San Jose, CA, USA). Cell death also measured by using a FITC-Annexin V Apoptosis. Detection kit (BD Phamingen) according to the manufacturer’s protocol.
G. RNA Interference
Human cervical cancer HeLa cells were plated and grown in 60 mm culture dishes. After overnight, they were transiently transfected with TRADD small interfering RNA or a nonsilencing control siRNA according to manufacturer’s instructions (Invitrogen). HeLa and U2OS cells plated in 60mm dish plates were transfected with 100pmol of TRADD (Bioneer) or non-targeting control RNAi oligo (Bioneer) using Lipofectamine 2000 (Invitrogen, Inc.). After 24 hours, cell were seeded in 6 well or 12 well plates and then treated with DNA damage agent. Knockdown efficiency of TRADD confirmed by western blotting. HepG2 cells
plated in 12-well plates were transfected with 50 pmol of TRADD or non-targeting control (NC) RNAi oligo (Dharmacon) using Lipofectamine 2000 reagent (Invitrogen, Inc).
H. Immunofluorescence
Cells (1.5 x104) were fixed in 4 % paraformaldehyde for 10 min at room temperature and permeated in 0.2 % Triton X-100. After blocking with 1 % BSA for 1 hour, cells were incubated with anti-γH2AX (Cell signaling) antibody for overnight at 4 °C. All fluorescence photographs were taken using a confocal microscope (Carl Zeiss, Germany).
I. Immunoprecipitation
For immunoprecipitation, lysates were mixed and precipitated with antibody and protein A-sepharose or protein G-agarose beads overnight at 4oC. Bound proteins were removed by boiling in SDS and resolved by SDS-PAGE for immunoblotting. Immunoblotting visualized by enhanced chemiluminescence (ECL, Amersham).
J. Pull Down Assay
Wild type and TRADD-/- MEFs (1 × 108) cells were treated with 10 μg/ml GST-TRAIL for 15 min or left untreated. Cells were then washed with PBS and lysed
with M2 buffer. The soluble fraction was incubated with 30μl of 50% glutathione-Sepharose beads (GE healthcare) for 3 h on a rotator at 4 °C. Bound proteins were eluted, resolved in 12% SDS–PAGE, and analyzed by immunoblotting.
K. HR- or NHEJ-mediated DSB repair GFP reporter systems
U2OS GFP reporter cell lines, U2OS-EJ5-GFP (NHEJ) and U2OS-DR-GFP (HR), were transfected with the indicated siRNAs by Lipofactamine RNAiMAX (Invitrogen). On the following day, I-SceI endonuclease was delivered to the cells, and 72 hr later, they were assayed for GFP-positive cells by flow cytometry (BD Biosciences) (Gunn et al., 2011).
L
. Nucleus and cytoplasm fractionation
For nuclear and cytoplasmic fractionation, cells were washed twice in ice-cold PBS. Nuclear and cytoplasmic extraction were performed using NE-PER nuclear and cytoplasmic extraction kit(Thermo, #7833) according to manufacturer`s instruction. equal amount of proteins were loaded in western blot.
M. Laser microirradiation
For the recruitment of NHEJ or HR repair factors at DNA break sites, U2OS cells were processed as described previously (Min, S et al., 2014).
N. FokI assay
For the recruitment of Myc-TRADD at single DNA double-strand break site (sDSB), mCherry-LacI-FokI and Myc-TRADD plasmids were cotransfected into U2OS 2-6-3 cell lines. After 48 hrs, fixed cells were stained with Myc antibody for detection of Myc-TRADD. Colocalization of mCherry-FokI and Myc-TRADD at sDSB was analyzed with a Nikon confocal microscope A1 (Min, S et al., 2014)
O. Statistical analysis
All results are presented as means ± SD. All experiments were performed at least three times. Excel (Microsoft, Redmond, WA, USA) was used for statistical analysis, and statistical significance was determined using Student's t-test. P<0.05 was considered significant.
III. RESULTS
A. TRADD is critical for resistance to TRAIL-Induced Cell Death
through NF-κB activation
1. Deficiency of TRADD sensitizes cells to TRAIL-induced cell death in MEFs
Treatment of TRADD-/- MEFs with recombinant murine TRAIL showed the
TRADD-/- MEFs were sensitive to TRAIL-induced cell death, while wild type (WT)
MEFs were resistant, suggesting TRADD prevented TRAIL-mediated cell death under normal circumstances (Figure 1A). Cycloheximide sensitized WT MEFs to TRAIL, however TRADD-/- MEFs continued to be more sensitive than the WT
MEFs to TRAIL as shown by contrast microscopy or MTT assay (Figure 1B) or by PI/Annexin V staining (Figure 1C). Though TRADD-/- MEFs were more sensitive
to CHX plus TRAIL than WT cells, they were completely refractory to CHX plus TNFα (Fig. 2 A). This suggests that TRADD-/- MEFs are not more generally
susceptible to cell death, and that TRAIL-induced sensitivity is stimulus specific. Cell death induced by TRAIL was completely blocked by zVAD, indicating that only apoptotic cell death was occurring in the TRADD-/- MEFs (Figure 2B). Consistent with this, we detected fully processed caspase 8, caspase 3, and PARP-1 cleavage in the TRAIL-treated TRADD-/- MEFs, but not in WT MEFs (Figure 2C), and zVAD efficiently blocked these events (Figure 2D).
Figure 1. Deletion of TRADD sensitizes cells to TRAIL-induced cell death.
(A, B) Phase contrast images (left) and MTT viability assay (right, Error bars: +/− S.E.M.) of wild type and TRADD-/- MEFs treated with mouse recombinant TRAIL
(40 ng/ml, 16 h). In panel B, cells were pretreated with CHX (0.1 μg/ml or 1 μg/ml). (C) Wild type and TRADD-/- MEFs were similarly treated and apoptotic cell
Figure 2. TRADD deficiency sensitizes cells to apoptosis induced by TRAIL but not TNFα.
(A, B) MTT assays of WT and TRADD-/- MEFs treated with CHX (1 μg/ml) and mTRAIL (40 ng/ml) or CHX (10 μg/ml) and TNFα (30 ng/ml) for 24 h (Error bars: +/− S.E.M.). In panel B, some cells were pretreated with zVAD (20 μM). (C, D). Immunoblot analysis in TRAIL-treated wild type and TRADD-/- MEFs.
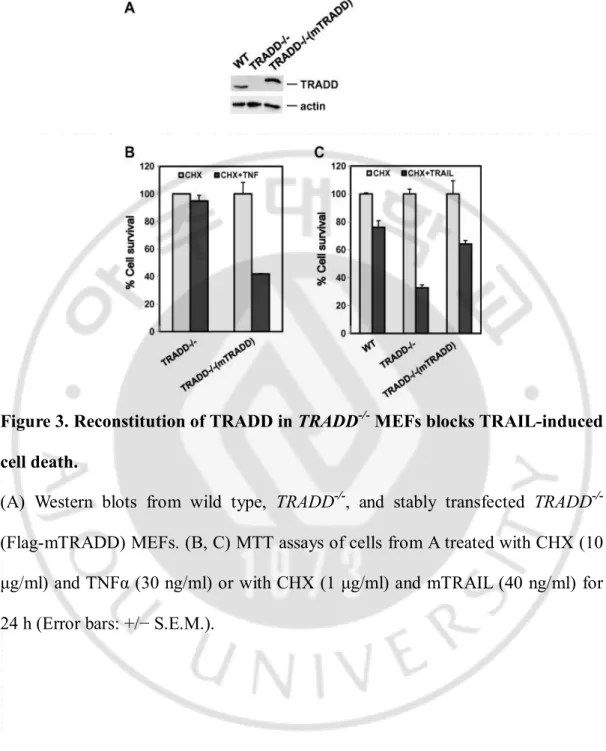
2. TRADD reconstitution suppressed TRAIL-induced apoptosis
We verified the involvement of TRADD in these events via stable TRADD reconstitution in TRADD-/- MEFs (Figure 3A). Reconstitution of TRADD expression restored TNFα-induced cell death in TRADD-/- MEFs (Figure 3B), but
substantially blocked TRAIL-induced cell death (Figure 3C), confirming that increased sensitivity of TRADD-/- MEFs to TRAIL was due to loss of TRADD.
Figure 3. Reconstitution of TRADD in TRADD-/- MEFs blocks TRAIL-induced cell death.
(A) Western blots from wild type, TRADD-/-, and stably transfected TRADD -/-(Flag-mTRADD) MEFs. (B, C) MTT assays of cells from A treated with CHX (10 μg/ml) and TNFα (30 ng/ml) or with CHX (1 μg/ml) and mTRAIL (40 ng/ml) for 24 h (Error bars: +/− S.E.M.).
3. TRAIL-induced NF-κB activation has pro-survival function in MEF cells
TRAIL can activate the transcription factor NF-κB, which can protect from TRAIL-induced apoptosis (Jeremias, I et al., 1998; Harper, N et al., 2001). To investigate whether TRAIL-induced NF-κB activation requires TRADD, we treated WT and TRADD-/- MEFs with GST-TRAIL because we were able to increase the activity in MEFs compared to mTRAIL (Figure 4A)(Lin, Y et al., 2000). While TRAIL activated IκBα phosphorylation and degradation in WT MEFs, GST-TRAIL-induced NF-κB activation was non-existent in TRADD-/- MEFs (Figure 4B).
Reconstitution of TRADD expression restored NF-κB activation in response to TRAIL as well as to TNFα in TRADD-/- MEFs (Figure 4C). These results strongly
suggest that TRADD is crucial for TRAIL-induced NF-κB activation and impaired NF-κB activation could explain the observed sensitization to TRAIL-induced apoptosis in TRADD-deficient cells. Inhibition of NF-κB activation by IκBα super suppressor in WT MEFs bestowed sensitivity to TRAIL (Figure 4D), while ectopic expression of IKKβ or p65 rescued TRADD-/- MEFs (Figure 4E) or HepG2 cells (Figure 4F) in response to TRAIL. Taken together, our data suggest that TRADD protects cells from TRAIL-induced cell death through NF-κB activation.
Figure 4. TRAIL-induced NF-κB activation is impaired in TRADD-/- MEFs.
(A) Wild type and TRADD-/- MEFs were treated with CHX (0.1 μg/ml) or CHX /GST-TRAIL (40 ng/ml) for 16 h and viability was analyzed by MTT assay. (B and C) Immunoblots from cell lysates of wild type, TRADD-/- (B) or stably
transfected TRADD-/- (Flag-mTRADD) MEFs (C) treated with GST-TRAIL (40 ng /ml) or TNFα (30 ng/ml) for indicated time points.
(D-F) MTT assay of cells transfected with IκBα super suppressor, IKKβ, p65 (RelA), or empty vector, then treated with GST-TRAIL (D, E: 40 ng/ml for WT and
TRADD-/- MEFs and F: 50 ng/ml for HepG2 cells) for 24 h (A, D, E, F Error bars:
4. Impaired JNK and ERK activation by TRAIL in TRADD-/- MEFs
MAPKs are readily activated in response to various stresses, and play key roles in deciding cell fate (Johnson G.L et al., 2002; Chang L et al., 2001). v MEFs lacked TRAIL-induced activation of ERK and JNK, while the activation of p38 was not affected (Figure 5A). However, the MAPK inhibitors, SP 600125 for JNK, PD 98059 for ERK, and SB 202190 for p38, respectively, did not influence TRAIL-induced cell death in either WT and TRADD-/- MEFs (Figure 5B), indicating that TRAIL-induced cell death in MEFs is independent of MAPK activity. In contrast, when NF-κB activation was blocked in WT MEFs or HepG2 cells through IKK inhibition, TRAIL-induced cell death was dramatically increased (Figure 5C and 5D) suggesting the NF-κB defect is responsible for the their susceptibility to TRAIL-induced cell death.
Figure 5. TRAIL-induced JNK and ERK activation are impaired in TRADD -/-MEFs, but do not contribute to cell death sensitivity.
(A) Immunoblot analysis of wild type and TRADD-/- MEFs treated with GST-TRAIL (40 ng/ml) for indicated time points.
(B) MTT assay of wild type and TRADD-/- MEFs pretreated with MAPK inhibitors, SP, PD, and SB for 30 min before CHX and GST-TRAIL (40 ng/ml) treatment for overnight.
(C and D) MTT assay of wild type MEFs (C) and HepG2 cells (D) pretreated with IKK-2 inhibitor V for 30 min before GST-TRAIL treatment (24 h). (B–D Error bars: +/− S.E.M.).
5. Knockdown of TRADD sensitizes cancer cells to TRAIL-induced cell death
TRAIL a highly promising cancer therapeutic agent, however, cancer cells can acquire resistance to TRAIL-induced cytotoxicity. We sought to determine whether TRADD knockdown can also sensitize cancer cells to TRAIL. Knockdown of TRADD expression in HepG2 cells (Figure 6A) led to an increased sensitivity of HepG2 cells to TRAIL (Figure 6B and 6C), suggesting that targeting TRADD is able to sensitize human cancer cells to TRAIL-induced cytotoxicity.
Figure 6. Depletion of TRADD can sensitize cancer cell to TRAIL-induced cell death.
(A–C) Western blots of lysates from HepG2 cells transfected with non-targeting control siRNA (NC) or TRADD siRNA (48 h). Cells were treated with GST-TRAIL (50 ng/ml, 12 h) and cell viability was analyzed by LDH leakage assay (B) (Error bars: +/− S.E.M.) or PI/Annexin V staining (C).
6. TRAIL-induced DISC complex formation
Finally, we investigated recruitment of signaling adaptors to the TRAIL receptor. We were able to detect RIP1 in the complex in both MEF cell types (Figure 7), though RIP1 recruitment to the receptor complex lasted longer in
TRADD-/- MEFs. Although less clear, FADD recruitment to the DISC also appeared
to slightly increase in TRADD-/- MEFs (Figure 7). We therefore hypothesize that
TRADD may compete with FADD and/or RIP1 for DISC recruitment and the additional FADD recruitment in TRADD-/- MEFs may contribute to enhanced
Figure 7. GST-pull down assay. DISC formation was detected by GST-pull down assay.
WT and TRADD-/- MEFs were treated with GST-TRAIL (40 ng/ml) for indicated
time points and GST-TRAIL-bound protein complexes were pulled down by glutathione-agarose beads and analyzed for presence of RIP1 and FADD by western blot.
B. Nuclear TRADD has a Protective Function During DNA
Damage Responses
1. TRADD is involved in the Hydrogen Peroxide-Induced DNA Damage Response.
Although the cytoplasmic functions of TRADD have been investigated intensively, much less is known about its function in the nucleus. To investigate this role, we first tested whether TRADD status affects the cellular response to DNA damage induced by hydrogen peroxide (H2O2), which generates hydroxyl radicals in the presence of transition metal ions, and can reach the nucleus to cause DNA strand breaks. TRADD wild type (TRADD+/+) and TRADD knockout (TRADD-/-) MEFs were treated with H2O2 and the phosphorylation (at Ser 139) of histone H2AX (γH2AX), one of the major markers for DNA double strand breaks (DSBs) (Clingen et al., 2008), was followed. Deficiency of TRADD potentiated the appearance of γH2AX in response to H2O2 in both western blotting and immunofluorescence staining (Figure 1A and 1B), indicating that TRADD may be involved in the DNA damage response (DDR). In further experiments, cell growth media was replaced after 2 hours of H2O2 treatment of the TRADD+/+ and TRADD
-/- MEFs, to prevent further DNA damage. In the presence of TRADD, γH2AX was
significantly reduced or gone 4 hours later; however, γH2AX detection largely persisted in TRADD-/- MEFs (Figure 1C and 1D). Furthermore, ectopic expression
of TRADD in TRADD-/- MEFs inhibited the appearance of H2AX in response to H2O2 (Figure 1E), confirming that this response was specifically due to TRADD deficiency in these MEFs, and not an artifact of MEF cell line generation. To determine whether TRADD-deficiency sensitized to DSBs in other cell types, we reduced TRADD expression in HeLa cells by siRNA knock-down. In these TRADD knock-down (KD) cells, TNFR1-mediated NF- B activation and ce ll death were inhibited (data not shown), but hydrogen peroxide induced-γH2AX was substantially increased in response to H2O2 (Figure 1F). Taken together, these data suggest that TRADD is involved in the cellular response and DSB repair.
Figure 1. Deficiency of TRADD induced impaired DNA Damage Response A. Western blotting analysis shows blotting for γH2AX, TRADD and Actin in
TRADD+/+ and TRADD-/- MEF cells treated with H2O2 (0.5 mM)
B. γH2AX foci were analyzed in TRADD+/+ and TRADD-/- MEF cells treated with
H2O2 (0.5 mM) at indicated time periods by Immunofluorescense.
C-D. Immunofluorescence analysis of γH2AX (green) in H2O2 treated TRADD+/+ and TRADD-/- MEF (upper panels). Cells were stained with anti-γH2AX (Green) and DAPI (Blue) after treated with H2O2 (0.5 mM) for 1 hour. Western blotting analysis (lower panels) shows results consistent with immunofluorescence.
E. Reconstitution of TRADD in TRADD-/- MEFs. Western blotting analysis shows
γH2AX expression in response to H2O2 (0.5 mM).
F. Transient knockdown of TRADD induces unrepaired DNA damage in HeLa cells. Western blot analysis shows γH2AX status in response to H2O2 in TRADD KD HeLa cells. Cells were transfected with siRNA TRADD or siRNA CONTROL, respectively. After 48 hours, the cells were treated with H2O2 (0.5 mM).
2. Depletion of TRADD sensitizes to DNA-Damage.
Hydrogen peroxide induces a large amount of different cellular damage to DNA and proteins. To verify the involvement of TRADD in the DNA damage response, we treated cells with specific DNA damage-inducing agents. As shown in Figure 2A, deficiency of TRADD potentiates appearance of the dose-specific gH2AX response to the alkylating agent N-methyl-N′-nitro-N′-nitrosoguanidine (MNNG); doxorubicin also has a similar pronounced effect in TRADD-deficient cells. As expected, various DNA damage-inducing agents including doxorubicin (Doxo), etoposide (Etopo), camptothecin (Cpt), and hydroxyurea (Hu) resulted in a differential increase of gH2AX in TRADD-/- MEFs (Figure 2B), but cisplatin
(CDDP) had a minimal effect on gH2AX. Etoposide, in particular, led to a more potent activation of gH2AX in TRADD-/- MEFs compare to TRADD+/+ MEFs.
TRADD+/+ and TRADD-/- MEFs were treated with etoposide and after 1 hour later,
the media was replaced with fresh media to remove DNA damage agents. In the presence of TRADD, γH2AX foci were significantly reduced compared with
TRADD-/- MEFs, as were γH2AX detection by immunostaining and western
blotting (Figure 2C). Increased gH2AX was further observed in TRADD KD U2OS cells treated with phleomycin, DSB-mimetic drug (Figure 2D). Thus, these results indicate that TRADD is required for the increased DNA repair and/or reduced DNA damage.
Figure 2. TRADD regulates DNA repair following DNA-damage agent treatment A. Western blotting analysis show γH2AX and Actin in TRADD+/+ and TRADD
-/-MEF cells treated with MNNG (0.25 μM) and Doxorubicin (2.5 μM).
B. Impaired DDR (DNA damage response) in TRADD-/- MEFs is a common
phenotype. Deficiency of TRADD induces unrepaired DNA damage compared with wild type cells in response to various DNA damaging agents (Doxo 2.5 μM, Etoposide 25 μM, Cpt 2.5 μg/mL, CDDP 25 μM, Hu 2.5 mM)
C. Immunofluorescence analysis of γH2AX (Red) in etoposide-treated TRADD+/+ and TRADD-/- MEF (upper panels). After cells were treated with etoposide (25 μM) for 1 hour, cells were stained with Anti-γH2AX (Red) and DAPI (Blue). Western blotting analysis (lower panels) shows the consistent results with Immunofluorescence.
D. Quantitative analysis of γH2AX foci was conducted in TRADD knock-downed U2OS cells. After TRADD knockdown, cells were treated with phleomycin and then stained with γH2AX antibody.
3. TRADD translocates to the nucleus following DNA damage.
To monitor the translocation of TRADD into nucleus in response to DNA damage, we transfected a GFP-TRADD plasmid into HeLa cells. As expected, when cells were treated with hydrogen peroxide to induced DNA damage, GFP-TRADD was translocated into nucleus (Figure 3A). This translocation began as early as 10-15 minutes post-treatment, and further increased in a time-dependent manner, as measured by time-lapse confocal microscopy (Figure 3B and 3C). To determine whether TRADD was recruited specifically DNA break sites, we used the mCherry-LacI-FokI endonuclease system, which targets mCherry-tagged FokI nuclease activity specifically to lac operator repeats (Min, S et al., 2014). The majority of Myc-TRADD was localized in the cytosol rather than in nucleus without DNA damage, whereas some translocated into nucleus upon DNA damage (Figure 3D). Furthermore, the nuclear translocated Myc-TRADD was colocalized with mCherry-FokI signal, suggesting that cytosolic TRADD is recruited to the nucleus at DNA breakage sites upon DNA damage.
Figure 3. DNA-Damage induces nuclear translocation of TRADD
A. HeLa cells were transiently transfected with GFP-TRADD and treated with H2O2 (0.5 mM) for indicated times. Cells were analyzed by confocal fluorescence microscopy.
B. HeLa cells were transfected with GFP-TRADD and treated with H2O2 (0.5 mM) for indicated time (minute). Cells were analyzed by JuLI stage (live cell imaging system)
C. HeLa cells were transiently transfected with GFP-TRADD and treated with H2O2 (0.5 mM). After treatment, live cell Images were analyzed by confocal fluorescence microscopy for 1 hour (left panel). Quantitative analysis of nuclear translocation of TRADD was measured by GFP intensity in the nucleus (right Panel).
D. Colocalization of Myc-TRADD and mCherry-FokI at single DNA double-strand break site. Constructs of Myc-TRADD and mCherry-FokI were cotransfected into U2OS 2-6-3 cell lines. After 48 hr, cells were fixed and stained with anti-53BP1 and anti-Myc antibodies. DAPI was used for nuclear staining. Images were analyzed confocal microscope (Nikon A1). 2D, two-dimensional analysis for images; 3D, three-dimensional analysis for images. Scale bar, 10 uM.
4. Nuclear TRADD is necessary for DNA-damage response.
TRADD has an NLS and an NES, which causes shuttling between nucleus and cytosol (Morgan et al., 2002). To clarify the role of TRADD translocation into the nucleus, we expressed previously-generated TRADD constructs (Morgan
et al., 2002) after testing their expression and localization. The NES
mutant-TRADD localizes both in the nucleus and cytosol, but once imported into the nucleus, which cannot translocate to the cytosol, since it lacks an export sequence. Cytoplasmic-TRADD is a C-terminal deletion lacking the NLS that localizes exclusively in the cytosol, while Src-myr-TRADD is largely confined to the plasma membrane by ectopic myristolyation. HeLa cells were transfected with different TRADD constructs and their localization was verified by fluorescence microscopy (Figure 4A). We found that NES mutant-TRADD and endogenous TRADD were accumulated in the nucleus after H2O2 or MNNG treatment, as measured by western blotting of nuclear and cytoplasmic fractions (Figure 4B).
TRADD-/- MEFs were transfected with different TRADD plasmids (Figure
4C). Reconstitution of NES mutant TRADD in TRADD-/- MEFs repressed the
increase of gH2AX and gH2AX foci in response to H2O2 treatment (Figure 4D) but expression of Src-myr-TRADD, which is confined to the plasma membrane had no effect on gH2AX status upon H2O2 treatment (Figure 4D, right panel), indicating that that nuclear localization of TRADD is required for the increased DNA repair or reduced DNA damage.
Figure 4. Nuclear localization of TRADD required for DNA repair
A. Localization of TRADD constructs transfected into HeLa cells. HeLa cells were transfected with wild type-TRADD, cytoplasmic-TRADD, Nuclear-TRADD, NES (Nuclear Export Sequence) mutant-TRADD and Src-myristoylated-TRADD. B. HeLa cell were transiently transfected with NES mutant TRADD and treated with H2O2 (0.5 mM) and MNNG (0.25 μM) for indicated times. Cells were fractionated into cytoplasmic and nuclear fractions using an NE-PER fractionation kit. Anti-Hsp90 and anti-Sp1 used as a control for normalization of cytoplasm and nuclear lysates, respectively.
C. Western blotting shows that expression of TRADD constructs in TRADD-/-
MEFs
D. Nuclear localization of TRADD regulates DNA-damage response. Western blotting analysis was conducted with lysates from TRADD-/-, TRADD-/- (NES-mutant TRADD) and TRADD-/- (Src-myristoylation -TRADD) MEFs treated with
H2O2 (0.5 mM) for indicated time periods. Expression of γH2AX was analyzed in
TRADD-/- and TRADD-/- (NES-mutant TRADD) MEFs treated with H
2O2 (0.5 mM) for 1hour using immunofluorescence.
5. TRADD is required for non-homologous end joining pathway.
There are two major DNA double strand break repair pathways: non-homologous end-joining (NHEJ) and non-homologous recombination (HR) repair (Ciccia et al., 2010 and Huertas, P et al., 2010). Since deficiency of TRADD revealed the persistence of γH2AX propagation with DNA damaging agents, we sought to determine which of these pathways that TRADD participates in. Depletion of TRADD with two different siRNAs (TRADD#1 and TRADD#2) was done in EJ-5 and DR GFP DNA repair reporter cell lines (Figure 5A). In EJ-5 cells, a promoter is separated from a GFP-coding sequence by a puro gene that has two I-SceI sites that are in the same orientation at opposite ends, such that EJ repair restores GFP expression. In DR-GFP cells, the genome contains a full-length GFP mutated to contain an I-SceI site, as well as a 5′ and a 3′-truncated GFP, which can then reconstitute GFP by HR machinery. After I-Sce expression, it was apparent that TRADD was required for efficient NHEJ repair, whereas this phenomenon was dispensable for HR repair (Figure 5B). TRADD silencing gave a similar effect as the knockdown of RAP80, a protein important in NHEJ repair. To evaluate whether TRADD is critical for recruitment of repair factors at DNA break sites, we laser microirradiated control and TRADD-KD U2OS cells, and stained with 53BP1 and Ku70/80 antibodies for NHEJ or with RPA32 and RAD51 antibodies for HR, respectively in control and TRADD-KD U2OS cells. NHEJ repair factors were dramatically diminished at laser strips after microirradiation, whereas factors for
HR pathway did not vary their localizations at DNA breaks (Figure 5C-5F). Interestingly, γH2AX recruitment patterns did not show any alterations in TRADD knockdown cells in these experiments, though overall fluorescence was decreased when quantitated (Figure 5C and 5E). However, the Ku70/80 complex, a major factor for NHEJ, binding to DNA double-strand break ends, was not accumulated at DNA break sites in TRADD KD cells, and there was a significant decrease in 53BP1 recruitment at these sites (Figure 5D). RPA32 and RAD51 were not decreased at the microirradiation strips in the TRADD-deficient cells (Figure 5E and 5F). 53BP1, but not RPA32, recruitment intensity, was also somewhat reduced at γH2AX foci in TRADD KD cells treated with phleomycin (Figure 5G). These findings suggest that TRADD mainly facilitates NHEJ repair rather than the HR repair of DNA breaks.
Figure 5. TRADD is required for non-homologous end joining repair.
A. Knockdown efficacy for TRADD in DNA repair reporter cell lines EJ5 and DR. B. After 48 hours transfection with TRADD, RPA80, or BRCA1 targeting siRNAs into reporter cell lines, each siRNA was again cotransfected with an I-SceI endonuclease construct. After 72 hours, GFP positive cells were analyzed with a flow cytometer (FACScan).
C and D. After 1 hour with laser microirradiation, endogenous NHEJ repair factors were stained with each antibody at DNA break sites: 53BP1 (C); Ku70/80 and 53BP1 (D).
E and F. Endogenous HR repair factors were stained as described in C. RAD51 (E); RPA32 and RAD51 (F). γH2AX was used as a DNA damage marker at DNA break sites in (C) and (E).
G. analysis of γH2AX foci, 53BP1 and RPA32 was conducted in TRADD knock-downed U2OS cells. After TRADD knockdown, cells were treated with phleomycin and then stained with γH2AX, 53BP1 and RPA32 antibody.
6. Deletion of TRADD sensitizes cells to DNA-damaging agent
It would be expected that a defect in a gene responsible for DNA damage repair frequently causes cell cycle arrest and cell death (Wang Y.N. et al., 2016). We, therefore, investigated whether depletion of TRADD would sensitize cell to DNA damage agents. Indeed, TRADD-/- MEFs were more sensitive to H2O2 treatment, but were completely resistant to TNF (Figure 6A). Reconstitution of TRADD in TRADD-/- MEF reduced H2O2-induced cell death compared to TRADD -/- MEFs (Figure 6B). Consistent with this data, TRADD KD HeLa cells were more
sensitive to H2O2 treatment (Figure 6C). As shown in Figure 6D, doxorubicin- and etoposide-induced cell death were also increased in TRADD-/- MEFs, indicating that defects in in DNA damage repair by depletion of TRADD also causes sensitization of cell death induced by DNA-damaging agents.
To verify the role of nuclear localization TRADD on cell death, MEFs expressing various TRADD constructs were treated with H2O2 to induce cell death. As expected, TRADD-/- MEFs were completely resistant to TNF-induced cell death, but highly sensitive to H2O2 to induced-cell death (Figure 6E). Expression of NES mutant-TRADD made cells sensitive to TNF-induced cell death, but protected cells from H2O2; while the plasma membrane-anchored Src-myr TRADD was able to reconstitute TNF toxicity in TRADD-/- MEFs, it was unable to protect cells from H2O2 (Figure 6E). Taken together with our previous data, including data that show impaired DNA damage response in TRADD deficient cells, with nuclear TRADD
necessary to inhibit persistent γH2AX accumulation (Figure 4D). Our data indicate that nuclear TRADD is important NHEJ repair pathway and has an important nuclear role in survival in response to DNA damage. The accumulation of aberrant unrepaired DNA damage such as DSBs in TRADD deficient cells is likely to cell death, consistent with what is previously known about the toxicity of DSBs (Wang, Y.N et al., 2016).
Figure 6. Deletion of TRADD sensitizes to DNA-Damage agent induced cell death
A. TRADD+/+ and TRADD-/- MEFs were treated with H2O2 for 24 hours and then cell viability was analyzed by MTT assay.
B. TRADD+/+, TRADD-/- and TRADD-/- (mTRADD) cells were treated with H2O2 for 24 hours. Cell viability was analyzed by MTT assay.
C. HeLa cells were transfected with siRNA TRADD or siRNA CONTROL, respectively. After 48 hours, the cells were treated with H2O2 for 24 hours. Cell viability was analyzed by LDH assay.
D. TRADD+/+ and TRADD-/- MEFs were treated with doxorubicin and etoposide for 24 hours and then cell viability was analyzed by MTT assay.
E. MTT assay of TRADD+/+, TRADD-/-, TRADD-/- (NES mutant TRADD) and
TRADD-/- (Src-TRADD) MEFs treated with H2O2 and TNF (30 ng/ml) plus CHX
7. Deficiency of TRADD leads to ROS accumulation and prolonged JNK activation
DNA damaging agents can increase cellular ROS level; newly formed ROS can further contribute to DNA damage leading to cell death (Kang, M.A et al., 2012 and Rowe, L.A et al., 2008). To further verify the mechanism of sensitization in impaired DNA repair induced by deficiency of TRADD, we investigated the amount of intracellular ROS. ROS accumulated more readily in TRADD-/- MEFs compared to TRADD+/+ MEFs (Figure 7A) and unsurprisingly, the ROS scavenger,
N-Acetyl-L-cysteine (NAC), dramatically decreased cell death in TRADD-/- MEFs, indicating increased ROS in TRADD-/- MEFs contributes to hydrogen peroxide-
induced cell death (Figure 7B). It is well known that ROS induces prolonged JNK activation via inhibition of JNK phosphatase and that prolonged JNK activation contributes to cell death (Kamata H et al., 2005). Thus, we checked whether ROS induces this signals. Indeed, JNK has the prolonged activation in
TRADD-/- MEFs treated with H2O2 compared with TRADD+/+ MEFs (Figure 7C).
Moreover, the NES-mutant TRADD suppressed JNK activation compared with
TRADD-/- MEFs (Figure 7D). The JNK inhibitor (SP600145) inhibited H2O2
-induced cell death in TRADD+/+ and TRADD-/- MEFs (Figure 7E), while ROS
scavenger treatment inhibited H2O2-induced prolonged JNK activation (Figure 7F). As with other DNA damaging agents, TRADD-/- MEFs are more sensitive to
TRADD-/- MEFs (Figure 7G). However, MNNG-induced cell death is inhibited by NAC (Figure 7G). These data suggest that impaired DNA damage induced by deficiency of TRADD affects ROS generation and sustained JNK activation, further potentiating cell death. DNA repair pathways enable cancer cells to survive DNA damage induced by chemotherapeutic treatments, while unrepaired DSBs are highly toxic and can lead to cell death (Figure 8) (Helleday, T et al., 2008). Taken together, our data suggest that TRADD could be a potential target for DNA-damage based chemotherapeutic agent-induced cancer cell death.
Figure 7. TRADD Induces ROS accumulation and prolonged JNK activation A. TRADD+/+ and TRADD-/- MEFs were pretreated with H
2O2 (0.5 mM) for 4 hours followed by treatment with H2DCFDA (10 μM) for 30 minutes. ROS level was measured by flow cytometry.
B. MTT assays of TRADD+/+ and TRADD-/- MEFs treated with NAC (1 mM) and H2O2 (0.5 mM) for 24 hours (Error bars:+/- S.E.M).
C. Western blotting of lysates from TRADD+/+ and TRADD-/- MEFs treated with H2O2 (0.5 mM) for indicated time periods.
D. Western blotting of lysates from TRADD+/+, TRADD-/- and TRADD-/- (NES-mutant TRADD) MEFs treated with H2O2 (0.5 mM) for indicated time periods. E. TRADD+/+ and TRADD-/- MEFs were pretreated with JNK inhibitor (SP, 20 μM) for 30 minutes followed by treatment with H2O2 (0.5 mM) for 24 hours. Cell viability was analyzed by MTT assay.
F. Western blotting of lysates from TRADD+/+ and TRADD-/- MEFs pretreated with NAC (1 mM) for 30 minutes followed by treatment with H2O2 (0.5 mM) for indicated time periods and then analyzed by western blotting.
G. Western blotting of lysates from TRADD+/+ and TRADD-/- MEFs treated with MNNG (0.25 μM) for indicated time periods. MTT assays of TRADD+/+ and
8. proposed model
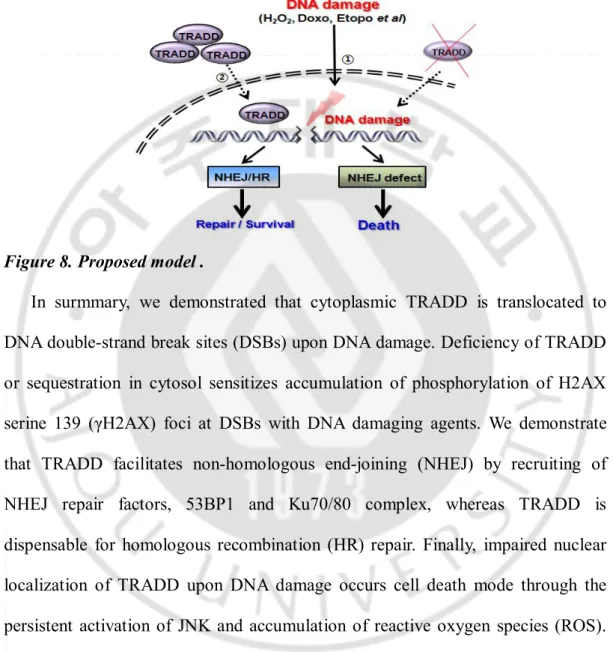
Figure 8. Proposed model .
In surmmary, we demonstrated that cytoplasmic TRADD is translocated to DNA double-strand break sites (DSBs) upon DNA damage. Deficiency of TRADD or sequestration in cytosol sensitizes accumulation of phosphorylation of H2AX serine 139 (γH2AX) foci at DSBs with DNA damaging agents. We demonstrate that TRADD facilitates non-homologous end-joining (NHEJ) by recruiting of NHEJ repair factors, 53BP1 and Ku70/80 complex, whereas TRADD is dispensable for homologous recombination (HR) repair. Finally, impaired nuclear localization of TRADD upon DNA damage occurs cell death mode through the persistent activation of JNK and accumulation of reactive oxygen species (ROS). Thus, our findings suggest that cytoplasmic TRADD needs to translocate at DSBs to modulate DNA repair.
IV. DISCUSSION
TRAIL is selective cytotoxic in cancer cells and therefore a promising anti-cancer therapeutic agent. However, anti-cancer cells often acquire TRAIL resistance and underlying molecular mechanisms are still elusive. To gain better understanding of TRAIL-induced signaling, we investigated that function of TRADD in this pathway. Deficiency of TRADD sensitizes cells to TRAIL's cytotoxicity, and cytotoxicity is blocked when TRADD expression is reconstituted. Thus, TRADD plays a protective role against TRAIL-induced cell death.
Our data indicate that TRAIL-induced NF-κB activation has pro-survival role in MEFs and TRADD is required for activation of this signaling pathway. This is consistent with several earlier reports showing the anti-apoptotic function of NF-κB in TRAIL-induced cell death (Lin Y et al., 2000). For instance, Lluis et al showed that NF-κB negatively regulated TRAIL induced cell death in TAK1 knockout MEFs by increasing c-FLIPL levels (Lluis J.M. et al., 2010) and a specific IKK inhibitor BMS-345541 or expression of IκBαM, which inhibits constitutively active NF-κB, confers sensitivity to TRAIL-induced cell death (Jani T.S. et al., 2010). In addition, IKKγ−/− MEFs are sensitive to TRAIL-induced cytotoxicity (Lluis J.M. et al., 2010).
Although our data suggest that lack of NF-κB activation is the primary reason for TRAIL-induced death of TRADD-/- cells, we do not exclude the possibility that TRADD also limits FADD recruitment to the DISC and that TRADD depletion
results in greater FADD recruitment, as has been recently suggested (Cao X. et al., 2011). We saw some evidence that this could be the case. Most likely both mechanisms play a role in the sensitivity of TRADD-/- cells to TRAIL. In contrast to this previous study (Cao X. et al., 2011), we observed that RIP1 was recruited to the TRAIL receptor complex in the absence of TRADD, and additional RIP1 recruited, suggesting that NF-κB deficiency occurs from loss of another TRADD-recruited component, such as cIAPs, that are necessary for NF-κB activation, and not from loss of RIP1 recruitment. Additional RIP1 recruitment in the absence of NF-κB activation could also play a role in greater caspase-8 activity by TRAIL as has been suggested for Fas (Morgan M.J et al., 2009).
There is mounting evidence that inhibition of NF-κB signaling leads to TRAIL-resistance by upregulating anti-apoptotic genes in multiple human cancer cells (Braeuer S.J et al., 2006; Ahn K.S. et al., 2007; Lanuti P. et al., 2009). Since TRADD depletion in cancer cells increases TRAIL-induced cell death, TRADD could therefore serve as a target for sensitizing cancer cells to TRAIL cytotoxicity.
It is well known that the TRADD serves as adaptor protein in cytoplasm and death receptor signaling. However, the function of TRADD in the nucleus poorly understood, though TRADD contains an NLS and an NES and shuttles through the nucleus. Nucleocytoplasmic shuttling of TRADD clearly affects distinct apoptosis mechanisms than are initiated by death receptor apparatus (Morgan, M et al., 2008). Chio and colleagues suggested that TRADD shuttles
dynamically from the cytoplasm into the nucleus to modulate the interaction between p19Arf and its E3 ubiquitin ligase ULF, thereby promoting p19Arf protein stability and thus tumor suppression (Chio, I.I.C et al., 2012). We show here that upon DNA damage, TRADD accumulates in the nucleus and participates in the NHEJ DNA repair process. Deficiency of TRADD leads to abnormal translocation of Rad51 and Ku70/80, indicating TRADD is required for the translocation of NHEJ related repair proteins to sites of DNA damage.
Using various TRADD constructs, we show that nuclear localization of TRADD is important for its DNA damage response functions. However, the mechanism of DNA damage-induced TRADD nuclear localization is still unclear. Unfortunately, post-translational modification of TRADD is not well-characterized. Using a phosphorylation prediction database, we examined the possibility that TRADD could be phosphorylated by various kinases involved in DNA-damage sensing. Interestingly, TRADD is predicted to be phosphorylated by ATM kinase (data not shown). Further study is required to determine if this putative phosphorylation event may play a role in TRADD’s nuclear functions.
Although DNA repair mechanisms seem to be well understood, the cellular response to genotoxic injury induced by conventional chemotherapies and radiotherapy is very complex. In mammals, NHEJ and HR are the two major pathways that repair DNA double-strand breaks (DSBs). The consequences of impairing either NHEJ or HR have been investigated in mice and similar
phenotypes are caused by defects in either pathway: hypersensitivity to ionizing radiation and TSA (Sharan, S.K. et al., 1997 and Lim, D.S. et al 1996 and Yaneva, M et al., 2005). The major proteins involved in NHEJ include the DNA-PKcs and the Ku70/80 heterodimer. These proteins have been reported to be up-regulated in tumors or radiation-resistant cell lines, indicating that NHEJ is likely to have a role in survival and resistance to DNA-damaging chemotherapy (Beskow, C. et al., 2009). In support of this, Ku70- or 80-depletion sensitizes pancreatic cells to IR, suggesting that it may be a potential target for inhibition in cancer therapy (Jekimovs, C. et al., 2014). Thus, the disruption of the DSB repair mechanisms by NHEJ may be of promise in the clinical target for the treatment of various cancers. Given our data, TRADD might be a potential drug target to promote cancer cell death in response to chemotherapeutic DNA-damaging agents.
In summary, our data show that deficiency of TRADD sensitizes to TRAIL and DNA damage-induced cell death through NF-kB and NHEJ pathway, respectively. Therefore, we suggest that TRADD is potential target for TRAIL- and chemotherapeutic agent-induced cancer cell death.
V. CONCLUSION
In conclusion, the present study showed that deficiency of TRADD sensitizes cells to TRAIL and DNA damage-induced cytotoxicity, but cytotoxicity is blocked when TRADD expression is reconstituted. Thus, TRADD plays a protective role against TRAIL- and DNA damage induced cell death. In TRAIL signaling, enhanced cell death in TRADD-/- MEFs is associated with defective NF-kB
activation. In DNA damage signaling, DNA damage promotes translocation of TRADD from cytoplasm to the nucleus. Nuclear localization of TRADD facilitates DNA damage repair. Since TRADD depletion in cancer cells increases TRAIL-and DNA damage-induced cell death, TRADD may be a good candidate as a target for sensitizing cancer cells to TRAIL- and DNA damage-induced cytotoxicity.
Ⅵ
. REFERENCES
1. Bender LM, Morgan MJ, Thomas LR, Liu ZG, Thorburn A. The adaptor protein TRADD activates distinct mechanisms of apoptosis from the nucleus and the cytoplasm. Cell Death Differ. 12(5):473-81, 2005
2. Beskow C, Skikuniene J, Holgersson A, Nilsson B, Lewensohn R, Kanter L. Radioresistant cervical cancer shows upregulation of the NHEJ proteins DNA-PKcs, Ku70 and Ku86. Br J Cancer. 101:816–2110.1038/sj.bjc.6605201, 2009
3. Chakraborty S, Li L, Tang H, Xie Y, Puliyappadamba VT, Raisanen J, Burma S, Boothman DA, Cochran B, Wu J, Habib AA. Cytoplasmic TRADD confers a worse prognosis in glioblastoma. Neoplasia , 15(8):888-97, 2013
4. Chio II, Sasaki M, Ghazarian D, Moreno J, Done S, Ueda T, Inoue S, Chang YL, Chen NJ, Mak TW. TRADD contributes to tumour suppression by regulating ULF-dependent p19Arf ubiquitylation. Nat Cell Biol, 14(6):625-33, 2012
5. Chen G, Goeddel DV. TNF-R1 signaling: a beautiful pathway. Science. 31;296(5573):1634-5, 2012
6. Ciccia, A., and Elledge, S.J. The DNA damage response: making it safe to play with knives. Mol Cell, 40, 179-204, 2010
7. Ermolaeva MA1, Michallet MC, Papadopoulou N, Utermöhlen O, Kranidioti K, kollias G, Tschopp J, Pasparakis M. Function of TRADD in tumor necrosis factor receptor 1 signaling and in TRIF-dependent inflammatory responses. Nat Immunol. 9(9):1037-46, 2008
8. G.L. Johnson , R. Lapadat. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science, 298: 1911– 1912, 2002
9. Gunn, A., Bennardo, N., Cheng, A., and Stark, J.M. Correct end use during end joining of multiple chromosomal double strand breaks is influenced by repair protein RAD50, DNA-dependent protein kinase DNA-PKcs, and transcription context. J Biol Chem, 286, 42470-42482, 2011
10. Gudjonsson T, Altmeyer M, Savic V, Toledo L, Dinant C, Grl te M, Bartkova J, Poulsen M, Oka Y, Bekker-Jensen S, Mailand N, Neumann B, Heriche JK, Shearer R, Saunders D, Bartek J, Lukas J, Lukas C. TRIP12 and UBR5 suppress spreading of chromatin ubiquitylation at damaged chromosomes. Cell. Aug 17;150(4):697-709, 2012
11. Huertas, P. DNA resection in eukaryotes: deciding how to fix the break. Nature
structural & molecular biology, 17, 11-16, 2010
12. Hsu H, Xiong J, Goeddel DV. The TNF receptor 1-associated protein TRADD signals cell death and NF-kappa B activation, Cell. 81(4):495-504, 1995
13. H. Hsu , H. Shu , M. Pan , D. Goeddel. TRAF2 and TRADD-FADD interactions define two distinct TNF receptor 1 signal transduction pathways.
Cell, 84: 299– 308, 1996
14. I. Jeremias , K.M. Debatin. TRAIL induces apoptosis and activation of NFkappaB. Eur. Cytokine Netw, 9: 687– 688, 1998
15. I. Jeremias , C. Kupatt , B. Baumann , I. Herr , T. Wirth , K.M. Debatin , Inhibition of nuclear factor kappaB activation attenuates apoptosis resistance in lymphoid cells. Blood, 91: 4624– 4631, 1998
16. J.M. Lluis. TAK1 is required for survival of mouse fibroblasts treated with TRAIL, and does so by NF-kappaB dependent induction of cFLIPL. PLoS One, 5: e8620–, 2010
17. Jekimovs C, Bolderson E, Suraweera A, Adams M, O'Byrne KJ, Richard DJ. Chemotherapeutic compounds targeting the DNA double-strand break repair pathways: the good, the bad, and the promising. Front Oncol. Apr 22;4:86, 2014
18. K.S. Ahn, G. Sethi, B.B. Aggarwal, Embelin. an inhibitor of X chromosome-linked inhibitor-of-apoptosis protein, blocks nuclear factor-kappaB (NF-kappaB) signaling pathway leading to suppression of NF-kappaB-regulated antiapoptotic and metastatic gene products. Mol. Pharmacol, 71: 209– 219, 2007
19. Kim JY, Lee JY, Kim DG, Koo GB, Yu JW, Kim YS.TRADD is critical for resistance to TRAIL-induced cell death through NF-κB activation. FEBS Lett. 585(14):2144-50, 2011
20. Kamata H, Honda S, Maeda S, Chang L, Hirata H, Karin M.Reactive oxygen species promote TNF alpha-induced death and sustained JNK activation by inhibiting MAP kinase phosphatases. Cell. 120(5):649-61, 2005
21. L. Chang , M. Karin , Mammalian MAP kinase signaling cascades. Nature, 410: 37– 40, 2001
embryonic lethal that is suppressed by a mutation in p53. Mol. Cell. Biol. 167133– 7143, 1996
23. Michael Morgan, Jacqueline Thorburn, Pier Paolo Pandolfi, and Andrew Thorburn. Nuclear and cytoplasmic shuttling of TRADD induces apoptosis via different mechanisms. J Cell Biol. 157(6): 975–984, 2002
24. Micheau O, Tschopp J. Induction of TNF receptor I-mediated apoptosis via two sequential signaling complexes. Cell. 114(2):181-90, 2003
25. M.J. Morgan, Y.S. Kim, Z.G. Liu, Membrane-bound Fas ligand requires RIP1 for efficient activation of caspase-8 within the death-inducing signaling complex. J.
Immunol., 183: 3278– 3284, 2009
26. Martin Loignon, Lilian Amrein, Michael Dunn and Raquel Aloyz. XRCC3 Depletion Induces Spontaneous DNA Breaks and p53-Dependent Cell Death. Cell Cycle. Mar 1;6(5):606-11, 2007
27. Mariana Yaneva, Han Li,Teresa Marple, and Paul Hasty. Non-homologous end joining, but not homologous recombination, enables survival for cells exposed to a histone deacetylase inhibitor. Nucleic Acids Res. 2005 Sep 21;33(16):5320-30.
28. Michallet MC, Meylan E, Ermolaeva MA, Vazquez J, Rebsamen M, Curran J, Poeck H, Bscheider M, Hartmann G, König M, Kalinke U, Pasparakis M, Tschopp J. TRADD protein is an essential component of the RIG-like helicase antiviral pathway. Immunity. 28(5):651-61, 2008
29. N. Harper, S.N. Farrow, A. Kaptein, G.M. Cohen, M. MacFarlane, Modulation of tumor necrosis factor apoptosis-inducing ligand- induced NF-kappa B activation by inhibition of apical caspases. J. Biol. Chem., 276: 34743– 34752, 2001
30. P. Lanuti , Enhancement of TRAIL cytotoxicity by AG-490 in human ALL cells is characterized by downregulation of cIAP-1 and cIAP-2 through inhibition of Jak2/Stat3. Cell Res, 19: 1079– 1089, 2009
31. Pobezinskaya YL, Kim YS, Choksi S, Morgan MJ, Li T, Liu C, Liu Z. The function of TRADD in signaling through tumor necrosis factor receptor 1 and TRIF-dependent Toll-like receptors. Nat Immunol. 9(9):1047-54. 2008
32. Patricia S. Hähnel, Birgit Enders, Daniel Sasca, Wynand P. Roos, Bernd Kaina, Lars Bullinger, Matthias Theobald and Thomas Kindler. Targeting components of the alternative NHEJ pathway sensitizes KRAS mutant leukemic cells to