■
Hea-Woon Park, MD, PhD; Jo-Young Son, MS; Yun-Woo Cho, MD, PhD
1; Se-Jin Hwang, MD, PhD
2; Su-Jeong Kim, PhD
3; Sang-Ho Ahn, MD, PhD
1; Sung-Ho Jang, MD, PhD
1; Yong-Jae Jung, MD
1■
Department of Rehabilitation Medicine, School of Medicine, Catholic University of Deagu;
1Department of Rehabilita- tion Medicine, College of Medicine, Yeungnam University;
2Department of Anatomy and Cell Biology, College of Medicine, Hanyang University;
3Institute of Medical Science, Yeungnam University
Purpose: The purpose of this study was to evaluate the development of pain behavior and the expression of CCL2/CCR2 and CX3CL1/CX3CR1 at above and below the level of hemisection of the spinal cord in a rat model.
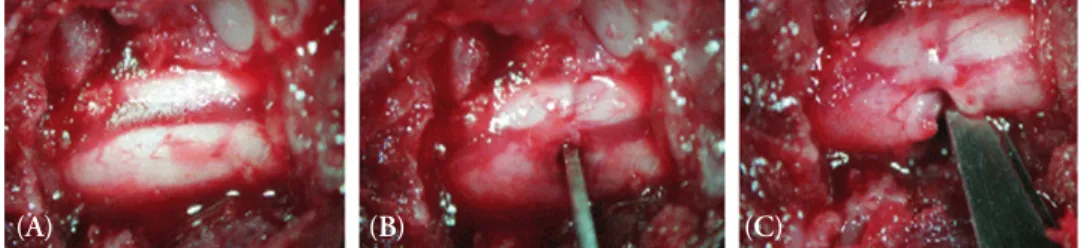
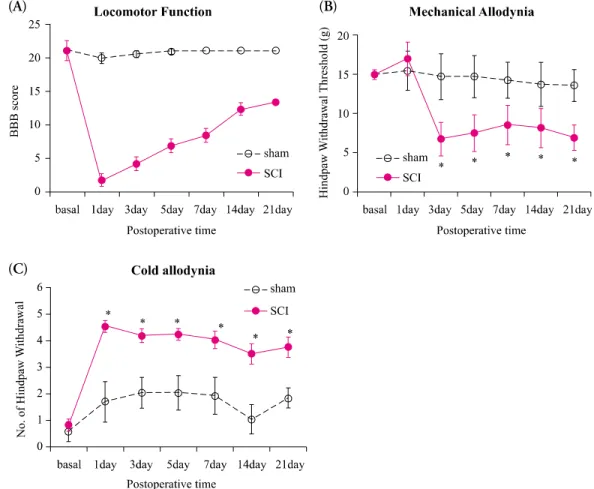
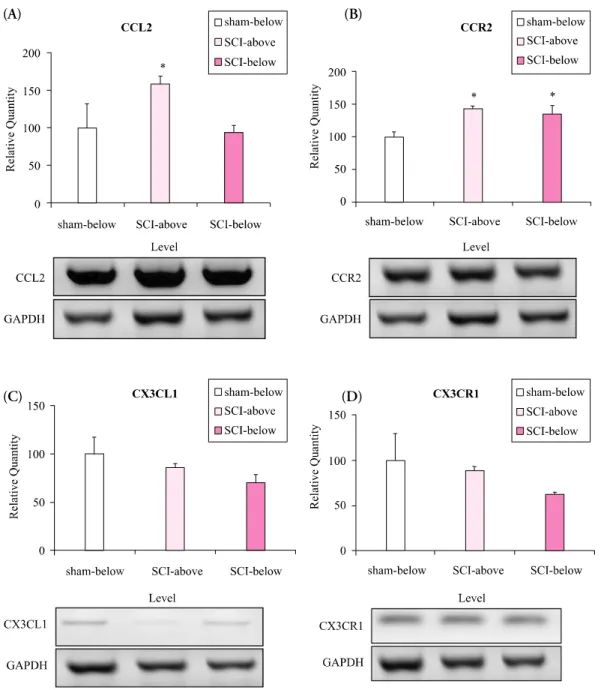
Methods: Spinal cords of adult female Sprague-Dawley rats (n= 16, 200~250 g, 6~8 weeks old) were hemisected at T13 on the right side to develop the spinal hemisection injury model. We compared behavioral responses of the hemisection and of a sham surgery group. Behavioral tests for motor function (by the BBB locomotor scale), and for pain response for mechanical and cold allodynia were assessed postoperatively (PO) for 21 days. Expression of mRNA for chemokines and their receptors (CCL2/CCR2 and CX3CL1/CX3CR1) below and above the level of the spinal cord dissection were examined by RT-PCR.
Results: We observed gradual motor improvement and the development of mechanical and cold allodynia on the ipsilateral hindpaw after spinal hemisection injury. We also found upregulation of mRNA expression of CCL2/CCR2 both above and below the level of spinal cord dissection but CX3CL1/CX3CR1 mRNA expression.
Conclusion: Upregulation of CCL2/CCR2 is associated with neuropathic pain after spinal hemisection injury. CCL2/CCR2 may play an important role in the development of neuropathic pain after SCI as well as of peripheral neuropathic pain. These findings may improve understanding of the pathophysiological mechanism of neuropathic pain after SCI.
Keywords: Spinal cord injury, Neuropathic pain, Chemokine, CCL2/CCR2, CX3CL1/CX3CR1 Received: May 19, 2010
Revised: June 9, 2010 Accepted: June 15, 2010
Corresponding author: Se-Jin Hwang, [email protected]; Sang-Ho Ahn, [email protected]
CCL2/CCR2 and CX3CL1/CX3CR1 after Spinal Cord Hemisection
The Journal Korean Society of Physical Therapy