736
FOREST SOCIETY
The Classification of Forest Communities by Cluster Analysis in Mt. Seokbyung Experimental Forest of Gangwon-Do
Sang Hoon Chung and Ji Hong Kim
*Department of Forest management, College of Forest & Environmental Sciences, Kangwon National University, Chuncheon, 200-701, Korea
Abstract :This study examined the ecological attributes of classified forest community by cluster analysis in the mixed forest of Mt. Seokbyung Experimental Forest of Gangwon-Do. The vegetation data were collected in randomly established 51 sample plots (2.04 ha) and analysis adopted the cluster analysis, importance value index, and Shannon's diversity index. Main results were as follows; 1) the study area was classified into 4 clusters (A, B, C and D). 2) The cluster A was dominated by Pinus densiflora with an importance value of 71.6%. The most dominant species in the cluster B and cluster C were Larix leptolepis (57.1%) and Quercus mongolica (40.2%), respectively. Finally, The cluster D was dominated by P.
densiflora (30.6%) and Q. mongolica (31.0%) with the mixed forest. 3) In the P. densiflora community (cluster A), distribution of DBH class showed a reverse J-shaped curve. In the L. leptolepis community (cluster B), individuals of dominant species had the bell-shaped distribution. Oak species indicated uniform distribution of DBH class (under 25 cm) in the mixed P. densiflora - Q. mongolica community (cluster D).
4) The species diversity index of the communities in descending order were: Pinus densiflora - Q.
mongolica community > Larix leptolepis community > Pinus densiflora community > Quercus mongolica community.
Key words :cluster analysis, forest community, importance value, distribution of dbh class, species diversity
Introduction
Statistical procedure for classification has been devel- oped to improve multivariate analysis of vegetation data in forest ecology, being accompanied by the advance- ment of computer application. They have played remark- able role in evolution of vegetation science from having mainly a qualitative assessment aspect to its present state as a highly quantitative discipline. Various references have reviewed and explained their technical development, stressed general discussion and comparison of the advan- tages for methods from one viewpoint to another. Some authors have introduced mathematically related method- ologies with explanation of details of individual tech- niques (Jongman et al., 1995; Legendre and Legendre, 1998; Young and Young, 1998; Krebs, 1999).
Given a sufficiently large group of forest vegetation, ecological clustering methods should be able to recog- nize clusters of similar vegetation while ignoring the few intermediates which often persist between clusters. Clus- tering is an operation of multidimensional analysis which
consists in partitioning the collection of vegetation data in the study. A partition is a division of a set into sub- sets. In another words, clustering is the process of data points by combining similar points to form small classes, then combining small classes into larger classes, and so on (Pielou, 1984). Likewise, cluster analysis is a type of analysis that classifies species, site, variables, or com- munities as done as in this study. The data tables of spe- cies by sample plots are made rearrangement, followed by the division of community types of which detailed characteristic information could be described and explained in terms of ecological phenomena for each type.
Cluster analysis is an explicit way of identifying groups in raw vegetation data and helps us to find struc- ture in the data. However, even if there is a continuous structure in the data, cluster analysis may impose a group of structure. Through the classification of forest communi- ties by cluster analysis, we try to secure information on the occurrence of species, that is, internal data structure, to establish forest community types for descriptive stud- ies, and to detect relations between communities and environmental variables (Pielou, 1984).
The primary objectives of this study was to apply the method of cluster analysis in a highly diversified mixed
*Corresponding author E-mail: [email protected]
forest of Mt. Seokbyung Experimental Forest of Gang- won-Do. The mixed forest is composed of a variety of plant species, which have evolved based on its high capability of adaptation to the climatic and environmen- tal conditions of the region. The aim of this study was to obtain ecological information for sustainable forest management of various forest communities. Based on floristic composition, cluster analysis was utilized to form and identify similar groups and outliers among sampling units, so as to classify the sampling units into several groups according to similar vegetation composi- tion, and to describe ecological attributes such as species composition, DBH & height classes, and species diver- sity in each type.
Materials and Methods
1. Site description
The study was conducted in the Mt. Seokbyung (37°
32'~34'N, 128° 52'~53') located in Jeongseon-gun, Gang- won-do in the east-central portion of the Korea (Figure 1). Forests in the study area were called “Economic For- ests for the Future”, which were managed by Forest Research Institute of Gangwon-do.
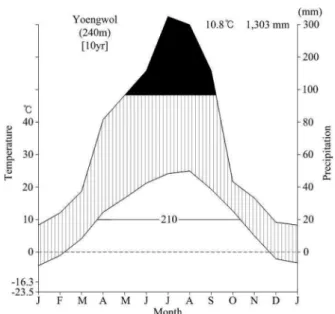
The mean annual temperature of the study area for the last 10 years was 10.8oC, and annual precipitation was 1,303 mm. Temperature of the warmest and coldest month were 24.8oC in August and -10.7oC in January, respectively (Korea Meterological Administration, 2008).
Mean annual precipitation was approximately 75% of that occurred during the summer season, June to Sep- tember (Figure 2). The elevation is from 500 m to 1,000 m above sea level. The vegetation of this area is mixed forest dominated mainly by Pinus densiflora and Quer-
cus mongolica. In addition, Larix leptolepis and Pinus koraiensis plantations have not received any silvicultural treatments except release cutting, creating a mosaic land- scape.
2. Data collection
In 2006, the vegetation data were collected in ran- domly established fifty one 20 m×20 m square sample plots of the study area (marked with black dots in Figure 1). To be able to describe the vertical structure of the stand, we divided into 3 layers (overstory, midstory, and understory). For each plot, we recorded plot coordinates and the diameter at breast height (DBH), total height, and species name for all woody plants.
3. Data analysis
In order to assort similar sampling units and to make group for the study forest according to the species com- position, the cluster analysis was carried out. The cluster analysis was based on for fifty one sampling units using the Ward's method of hierarchical clustering. Ward's method, also known as Minimum variance clustering, has been popular with ecologists (Everitt, 1974; Hartigan 1975;
Orloci 1967). This method has great intuitive appeal because it is based on the simple underlying principle that at each stage of clustering the variance within clusters is minimized with respect to the variance between cluster (Ludwig and Reynolds, 1988). The basal area data for all woody plants in the overstory of all sampling units were subjected to cluster analysis. The SPSS Statistics 17.0 (SPSS, Inc.) software was employed to implement the analysis.
The importance value (IV) gives an overall estimate of the influence or importance of a plant species in the Figure 1. Location of the study area within Jeongseon-
gun. The insert indicates the location of Jeongseon-gun, Gangwon-do in the Korea.
Figure 2. Climate diagram of the study area.
community (Brower and Zar, 1977). The sum of the three relative measures for species i is considered as the importance value (IVi) of the species by following equa- tion;
IV= (RD+RF+RC) / 3
where RD is the relative density, RF is the relative fre- quency, and RC is the relative basal area.
The species diversity in each cluster were quantified using (1) the Shannon's diversity index (H'), (2) the Maximum diversity (Hmax), (3) the Evenness (J'), and (4) the Dominance (1-J'). The species diversity were calcu- lated as;
H'= (N log N − ni log ni) / N, Hmax= logs, J'=H'/Hmax.
where N is the total number of individuals, and ni is the number of species i in each cluster. For this calculation, we used logarithmic base e.
Results and Discussion 1. Classification of forest communities
Cluster analysis is a classification technique for plac- ing similar entities or objects into groups or clusters. The cluster analysis models are used to place similar samples into clusters, which are arranged in a hierarchical tree-
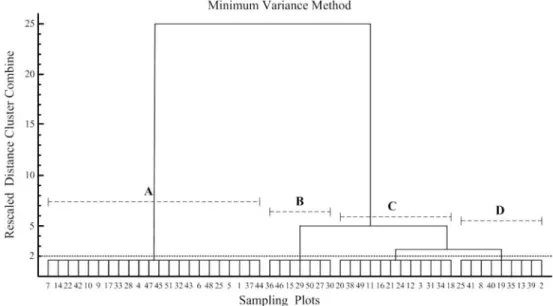
Figure 3. The dendrogram was yielded by minimum variance method (Ward's method) for fifty sampling plots in the study area.
Figure 4. Importance values (%) of woody species for P. densiflora community.
like structure called a dendrogram.
No. 23 sampling unit was excluded using K-means method of non-hierarchical clustering because it was an outlier in the sampling units. Using Ward's method for fifty sampling units and taking 'rescaled distance cluster combine' 2 as the standard, the study area was classified into 4 clusters (A, B, C and D), diagrammed in Figure 3.
2. Species composition
The species composition of the each plant community was given in Figure 4~7 with importance values of woody species. The total number of species at the four plant communities was 75.
Dominant species include Larix leptolepis, Pinus den-
siflora, Quercus mongolica, and Quercus serrata in the overstory of cluster A (Pinus densiflora community). Of those species, P. densiflora (71.6%) was the most dom- inant overstory constituent. The midstory and understory were mostly dominated by Q. mongolica (30.9%) and Rhododendron schlippenbachi (24.1%), respectively (Figure 4). Since P. densiflora is a pioneer tree species with shade intolerant characteristics, it appeared in the over- story and midstory, but did not present in the understory.
The most dominant species of the overstory in the cluster B (Larix leptolepis community) was L. leptolepis (57.1%). Dominant species in the midstory were L. lep- tolepis (22.0%) and Q. mongolica (16.8%). Tripterygium regelii (9.3%) and Q. mongolica (10.7%) were dominant
Figure 5. Importance values (%) of woody species for L. leptolepis community.
Figure 6. Importance values (%) of woody species for Q. mongolica community.
species in the understory (Figure 5). Like P. densiflora, L. leptolepis in P. densiflora community did not appear in the understory.
In the overstory and midstory of the cluster C (Quer- cus mongolica community), the most dominant species was Q. mongolica which was recorded in 40.2% of IV (Figure 6). In the understory, Pinus koraiensis was the most dominant tree species with an importance value of 17.6%. The appearance of P. koraiensis in the commu- nity was because seeds of that species were dispersed by rodents under natural condition. This phenomenon can also be found in the natural P. densiflora forest, L. lep- tolepis forest, P. rigida plantation, natural broadleaved forest, etc (Lee, 2002). Though P. koraiensis was shade tolerant species in the early life stage, when gap forms in the overstory, the species would experience high growth rate and may become a dominant tree species in the forest.
The overstory was dominated by Pinus densiflora (30.6%) and Quercus mongolica (31.0%) in cluster D (P.
densiflora - Q. mongolica community). The most dom- inant species of the mid-story was Q. mongolica (43.8%). In the understory of this community, the dominant species were Q. mongolica (19.5%) and Lespedeza maximowic- zii (17.4%). If the density of overstory Q. mongolica is moderately regulated by thinning, more ecologically healthy conifers (P. densiflora and P. koraiensis) and broadle- aved (Quercus species) mixed forest can be maintained (Figure 7).
In every community, the smaller number of species were composed of in overstory, mainly because only a few dominant species rose up to the canopy by superi- ority from adaptation of surrounding environment and
competition with other species during the growth in understory and midstory. Quercus species had highest value of IV in all community types and distributed in all three layers. It is indicated that the distribution range and abundance of Quercus species will be increased in suc- cessive generation of the forest by high potentiality of adaptation for the every given growing condition.
3. Distribution of DBH class
In the Pinus densiflora community, density distribu- tion of the trees by DBH class showed a reverse J- shaped distribution, the community was dominated by broadleaved species such as Q. mongolica, L. obtusi- loba, and R. schlippenbachii with diameter smaller than 15 cm. Shade tolerant species have an all age population structure with the characteristic reverse J-shaped curve or negative exponential stem distribution. As the DBH class increased, the percentage occupation by P. densi- flora became higher (Figure 8A). In addition, since the midstory was largely occupied by Q. mongolica and Q.
serrata with DBH 5-15cm, if living condition of mid- story can be improved due to gap formation in the over- story, Pinus densiflora stand in the study area would be replaced by mixed Q. mongolica, Q. serrata-Pinus den- siflora forest. Shade tolerant species, oak species in this case, have an all age population structure with the char- acteristic reverse J-shaped curve. These species are sta- ble species as far as the equilibrium stand distribution is concerned (Rollet, 1974; Veblen et al., 1980).
In the Larix leptolepis community, density distribution of L. leptolepis by DBH class was bell-shaped distribu- tion. While other broadleaved species, A. pseudo-siebold- ianum, F. rhynchophylla, Q. serrata and Q. mongolica, Figure 7. Importance values (%) of woody species for P. densiflora - Q. mongolica community.
showed a reverse J-shaped distribution (Figure 8B).
Bell-shaped distribution species are generally considered to be light demanding species. They depend on gaps in the canopy for their regeneration. Mortality is higher in the earlier stage under a closed forest canopy (Sokpon and Biaou, 2002). To relieve the competition stress and promote the diameter growth of L. leptolepis, thinning should be operated on the 15-25 cm DBH class which had the highest density of Larix leptolepis. Using this method, large diameter wood production of L. leptolepis can be realized.
In the Quercus mongolica community, density distri- bution of Q. mongolica by DBH class also showed a normal distribution. The overstory was largely occupied by broadleaved trees, midstory and understory were largely occupied by Pinus koraiensis and Acer mono
with DBH<10cm (Figure 8C). The range of DBH class distribution of Q. mongolica population was narrow and more or less bell shaped. As time passes, these trees grow older and larger, the tree DBH class distribution moving to the right on the graph like P. densiflora in Figue 8A.
In the Pinus densiflora - Quercus mongolica commu- nity, like the Q. mongolica community, density distribu- tion of Q. mongolica by DBH class showed a normal distribution, but overstory of the P. densiflora - Q. mon- golica community was largely occupied by P. densiforla and Q. mongolica (Figure 8D). As time passes by, the overstory P. densiflora in the P. densiflora community would gradually lost its dominance, and the community would be replaced by P. densiflora - Q. mongolica com- munity during the process of succession.
Figure 8. Distribution of major woody species by DBH class for each cluster.
Table 1. Species diversity indices of four plant communities.
Plant community No. of species Shannon's diversity index (H') Maximum
diversity (Hmax) Evenness
(J') Dominance (1-J')
Pinus densiflora (A) 60 2.839 4.094 0.693 0.307
Larix leptolepis (B) 38 2.723 3.638 0.749 0.251
Quercus mongolica (C) 39 2.431 3.664 0.663 0.337
Pinus densiflora - Quercus mongolica (D) 39 2.348 3.045 0.771 0.229
4. Species diversity
Species diversity index was the highest in the P. den- siflora community (2.839), and lowest in the P. densi- flora - Q. mongolica forest (2.348) (Table 1). Comparison of species diversity among the communities can not be made by using the species diversity index only. To have high species diversity index, both of species richness and evenness should be high (Kim, 1999). So species diver- sity should be judged by making comparison of species diversity index with maximum species diversity index and evenness index.
So the species diversity index of the communities in descending order were: P. densiflora - Q. mongolica com- munity, L. leptolepis community, P. densiflora community and Q. mongolica community. The highest species diversity in the P. densiflora - Q. mongolica community may due to the relatively high species evenness in that community, though the species richness was small.
Comparative studies of species diversity between plan- tations and other forest types have shown those planta- tions have either higher (Fahy and Gormally, 1998; Bhuju and Ohsaea, 1999; Nagaike, 2000) or lower (Peterken and Game, 1984; Michelsen et al., 1996; Amezaga and Onaindia, 1997; Qian et al., 1997; Stapanian and Cas- sell, 1999) diversity. In the former case, as in this study, the higher diversity in plantations than P. densiflora and Q. mongolica community. That was because a large vari- ety of outer species invaded and occupied the midstory and understory of the community with high species even- ness, due to lacking of sustainable management. Plant spe- cies diversity in the plantation understory has become a topic of increased interest worldwide (Ferris et al, 2000;
Lindgren and Sullivan, 2001).
Literature Cited
1. Amezaga, I. and Onaindia, M. 1997. The effect of ever- green and deciduous coniferous plantations on the field layer and seed bank of native woodlands. Ecography 20: 308-318.
2. Bhuju, D.R. and Ohsawa, M. 1999. Species dynamics and colonization patterns in an abandoned forest in an urban landscape. Ecological Research 14: 139-153.
3. Brower, J.E. and Zar, J.H. 1977. Field and laboratory methods for general ecology. Wm. C. Brown Company Publishers, USA, pp. 73.
4. Everitt, B. 1973. Cluster Analysis. John Wiley & Sons, Inc., New York.
5. Fahy, O., Gormally, M., 1998. A comparison of plant and carabid beetle communities in an Irish oak wood- land with a nearby conifer plantation and clearfelled site. Forest Ecology and Management 110: 263-273.
6. Ferris, R., Peace, A.J., Humphrey, J.W. and Broone,
A.C. 2000. Relationships between vegetation, site type and stand structure in coniferous plantations in Britain.
Forest Ecology and Management 136: 35-51.
7. Hansen, A.J., Spies, T.A., Swanson, F.J. and Ohmann, J.L., 1991. Conserving biodiversity in managed forests.
BioScience 41: 382-392.
8. Hartigan, J.A. 1975. Clustering Algorithms. John Wiley
& Sons, Inc., New York.
9. Jongman, R.H.G., Ter Braak, C.J.F. and Van Tongeren, O.F.R., 1995. Data analysis in community and land- scape ecology. Cambridge University Press. United King- 10. Kim, J.H. 1999. Forest Ecology; Attributions and anal-dom.
ysis of Forest Community. HyangMoonSa. pp. 208.
11. Korea Meterological Administration. 2008. Statistical Database.
12. Krebs, C.J. 1999. Ecological methodology. 2nd Ed. An Important of Addison Wesley Longman. USA.
13. Lee, W.S. 2002. Abundance and growth of naturally regenerated Pinus koraiensis wildlings in four foest types. Ph.D. Thesis. Kangwon National University. Korea.
14. Legendre, P. and Legendre, L. 1998. Numerical ecology.
Second english edition. Elsevier Science B.V. Netherlands.
15. Lindgren, P.M.F. and Sullivan, T.P. 2001. Influence of alternative vegetation management treatments on coni- fer plantation attributes: abundance, species diversity, and structural diversity. Forest Ecology and Manage- ment 142: 163-182.
16. Ludwig, J.A. and Reynolds J.F. 1988. Statistical Ecol- ogy. John Wiley & Sons, Inc., New York. pp. 125-144.
17. Michelsen, A., Lisanework, N., Friis, I. and Holst, N.
1996. Comparisons of understorey vegetation and soil fertility in plantations and adjacent natural forests in the Ethiopian highlands. Journal of Applied Ecology 33: 627-642.
18. Moore, S.E. and Allen, E.l. 1999. Plantation forestry.
In: Hunter Jr., M.L (Ed.), Maintaining Biodiversity in Forest Ecosystems. Cambridge University Press, Cam- bridge, pp. 400-433.
19. Nagaike, T. 2000. A review of ecological studies on plant species diversity in plantation ecosystems. Journal of Japanese Forest Society 82: 407-416. (in Japanese with English summary)
20. Nagaike, T. 2000. The effects of human disturbance on forested landscape structure and plant species diversity in Fagus crenata foreset regions. Bulletin of Yama- nashi Forest Research Institute 21: 29-85. (in Japanese with English summary).
21. Orloci, L. 1967. An agglomerative method for classi- fication of plant communities. Journal of Ecology 55:
193-206.
22. Peterken, G.F. and Game, M. 1984. historical factors affecting the number and distribution of vascular plant species in the woodlands of centeral Lincolnshire. Jour-
nal of Ecology 72: 155-182.
23. Pielou, E.C. 1984. The interpretation of ecological data - A primer on classification and ordination. John Wiley
& Sons. USA.
24. Qian, H., Klinka, K. and Sivak, B. 1997. Diversity of the understory vascular vegetation in 40-year-old and old-growth forest stands on Vancouver Island, British Columbia. Journal of Vegetation Science 8: 773-780.
25. Rollet, B. 1974. L'architecture des forts denses humides sempervirentes de plaines. C.T.F.T., Paris. pp. 298.
26. Sokpon, N. and Biaou, S.H. 2002. the use of diameter distributions in sustained-use management of remnant forests in Benin: case of Bassila forest reserve in North Benin. Forest Ecology and Management 161: 13-25.
27. Stapanian, M.A. and Cassell, D.L. 1999. Regional fre- quencies of tree species associated with anthoropogenic disturbances in three forest types. Forest Ecology and Management 117: 241-252.
28. Veblen, T.T., Schlegel, F.M. and Escobar, R.B. 1980.
Structure and dynamics of old-growth Nothofagus for- ests in the Valdivian Andes. Journal of Ecology 68: 1- 29. Young, L.J. and Yong, J.H. 1998. Statistical ecology; A31.
population perspective. Kluwer Academic Publishers.
London, United Kingdom.
(Received September 3, 2010; Accepted October 6, 2010)