Profiling Patterns of Volatile Organic Compounds in Intact, Senescent, and Litter Red Pine (Pinus densiflora Sieb. et Zucc.) Needles in Winter
17
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
수치
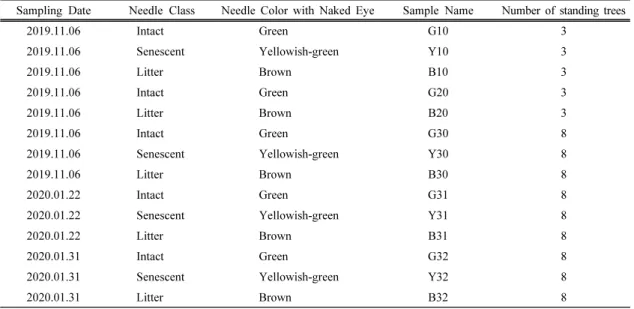

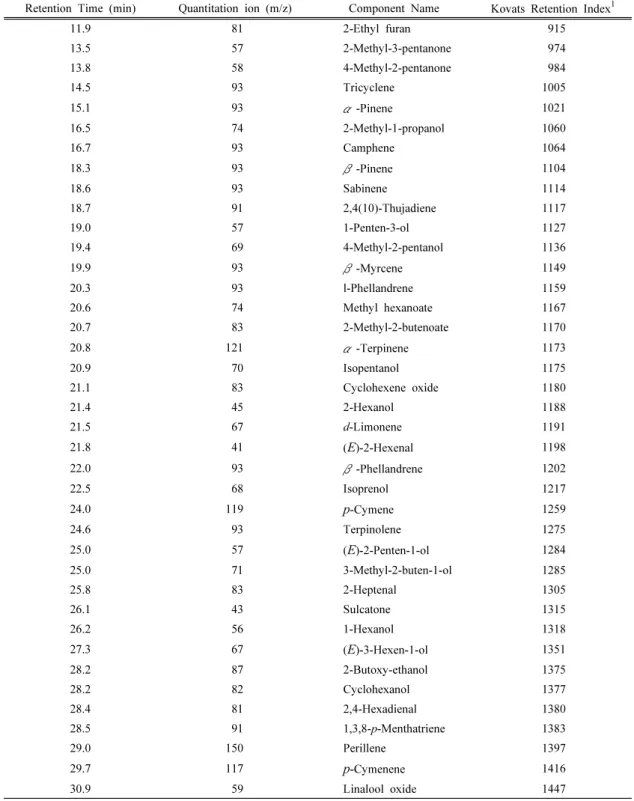
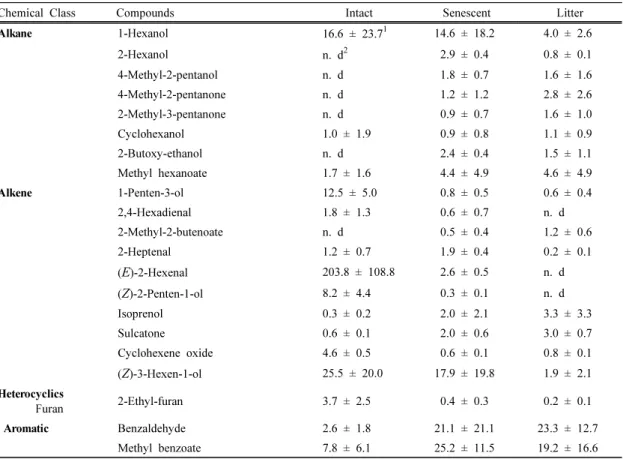
+5
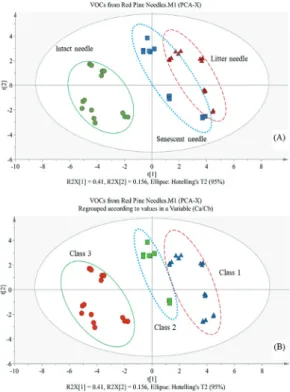
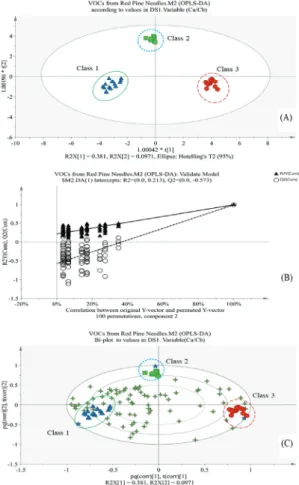
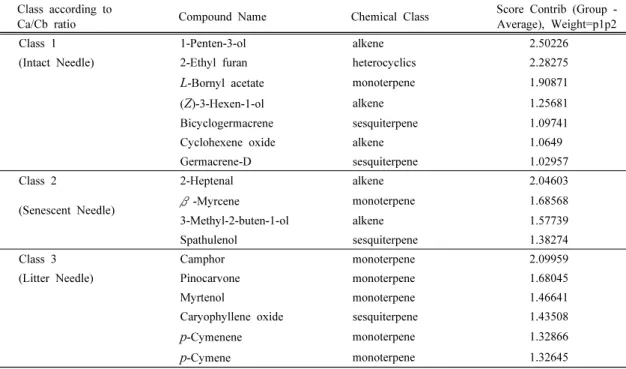
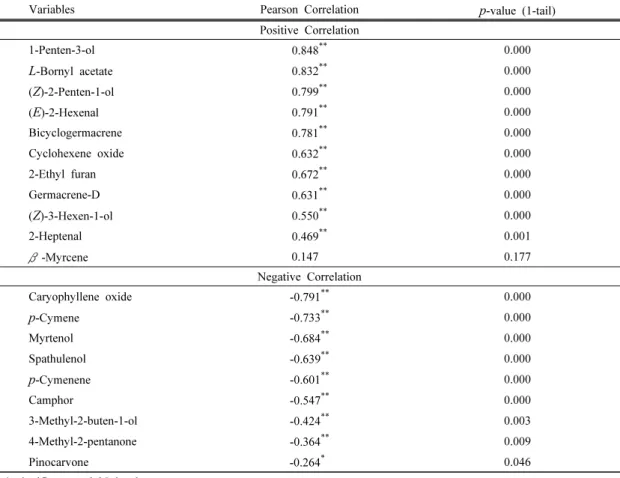
관련 문서
The purpose of this study is to investigate changes in listening, reading achievement, and learning attitudes of Korean elementary students who enrolled in
In this study, we have fabricated the 660 nm red LED illumination controller and LED array module and evaluated the proliferation and differentiation
This study was conducted to investigate the effects of 12 weeks after school physical activity program on body composition and PAPS in obese elementary
In SDE extraction, a total of 117, 115 and 120 volatile compounds were detected in Aster yomena, Aster tataricus and Chrysanthemum zawadskii, individually, whereas their total
The purpose of this study was to investigate how the BOSU Ball exercise, which can develop core muscles in middle-aged women, affects body composition and
Purpose: This study was aimed to compare the functional outcomes of the continuous and discontinuous fractures, retrospectively, in multiple thoracolumbar
To investigate the PL quenching sensitivity behaviors of the organic vapor, an organic vapor sensor based on rugate PSi filters was fabricated and tested
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
