THE KOREAN JOURNAL OF HEMATOLOGY R E V I E W A R T I C L E
Adenovirus as a new agent for multiple myeloma therapies:
Opportunities and restrictions
Svjetlana Raus
1, Silvia Coin
1, Vladia Monsurrò
1,21Department of Pathology and Diagnostics, University of Verona Medical School, 2Azienda Ospedaliera Universitaria Integrata, Verona, Italy
p-ISSN 1738-7949 / e-ISSN 2092-9129 http://dx.doi.org/10.5045/kjh.2011.46.4.229 Korean J Hematol 2011;46:229-38.
Received on December 3, 2011 Accepted on December 19, 2011
Multiple myeloma is a malignancy of B-cells that is characterized by the clonal expansion and accumulation of malignant plasma cells in the bone marrow. This disease remains incurable, and a median survival of 3-5 years has been reported with the use of current treatments. Viral-based therapies offer promising alternatives or possible integration with current therapeutic regimens. Among several gene therapy vectors and oncolytic agents, adenovirus has emerged as a promising agent, and it is already being used for the treatment of solid tumors in humans. The main concern with the clinical use of this vector has been its high immunogenicity; adenovirus is often able to induce a strong immune response in the host. Furthermore, new limitations in the efficacy of this therapy, intrinsic to the nature of tumor cells, have been recently observed. For example, our group showed a strong antiviral phenotype in vitro and in vivo in a subset of tumors, shedding new insights that may explain the partial failure of clinical trials based on this promising new therapy.
In this review, we describe novel therapeutic approaches that implement viral-based treat- ments in hematological malignancies and address the novelty as well as the possible limi- tations of these new therapies, especially in the context of the use of adenoviral vectors for treating multiple myeloma.
Key Words Adenovirus, Oncolytic therapy, Multiple myeloma, Antiviral phenotype
*This work was supported by the Italian Ministery of Health, Fondazione Cariverona and AIRC.
Correspondence to Vladia Monsurrò, M.D., Ph.D.
Department of Pathology and Diagnostics, Immunology Section, University of Verona Medical School, c/o Policlinico G.B. Rossi, P.le L.A. Scuro 10, 37134 Verona, Italy Tel: +39-45-812-6452
Fax: +39-45-812-6455
E-mail: vladia.monsurro@univr.it
Ⓒ 2011 Korean Society of Hematology
INTRODUCTION
Multiple myeloma (MM) is the most common primary bone cancer, representing 10% of all hematological malig- nancies and 1-2% of all cancer-related deaths [1], and occurs when malignant of B-cells progressively infiltrate the bone marrow and produce immunoglobulin after clonal expansion [2]. Conventional therapeutic protocols include chemo- therapy with bone marrow transplantation and drug treat- ment involving combinations of melphalan, vincristine, car- mustine (bischloroethylnitrosourea), cyclophosphamide, doxorubicin (Adriamycin), thalidomide, and prednisone and dexamethasone [3, 4]. These agents, used as monotherapies or in combination, have significantly improved MM out- comes, but long-term tolerance, graft-versus-host disease, and toxicities associated with some of these drugs represent great limitations [5]. The median survival is still only 3-5 years, and cases of relapse are frequent [6]. New drugs, such as bortezomib (Velcade) and lenalidomide (Revlimid), have recently been introduced as novel and more-curative
therapies. However, like with conventional treatments, long-term tolerance and toxicities associated with these drugs are major limitations [5]. Therefore, new therapies are defi- nitely needed.
Historically, remission of hematological malignancies, such as Burkitt’s lymphoma and Hodgkin’s disease, has been shown to be induced by clinical infection with the measles virus (MV) [7]. This finding paved the way for 2 major therapeutic strategies: The first is based on the use of viruses as oncolytic agents (http://www.hindawi.com/journals/av/
2012/186512/), since oncolytic viruses preferentially repli- cate in tumor cells by taking advantage of cancer-specific cellular changes [8]. This specificity is usually improved by deleting the E1A viral gene that is required for replication [9]. The second strategy, instead, uses different categories of viruses as possible vectors to deliver genes inside human tumor cells. Among the viruses used for viral therapy of tumors, 4 RNA viruses (MV, vesicular stomatitis virus, re- ovirus, and CVA21 [coxsackievirus A21]) and 2 DNA viruses (adenovirus and VV [vaccinia virus]) have been studied, with the goal of finding a translational application for the treat-
ment of multiple myeloma [10].
The adenovirus vectors have been the most commonly used vectors in human cancer treatment, especially for cancer gene therapy strategies based on intratumoral injection (Supplementary Table 1) [11]. Therefore, the first part of this review focuses on adenovirus and adenovirus-derived vectors and describes the state-of-the-art application of these viruses as oncolytic or gene therapy vectors and the potential therapeutic use of adenovirus vectors for the cure of MM.
In the second part of this review, we discuss a significant new cancer antiviral phenotype described by our group that is shared by solid and hematologic malignancies and that may prove to be one of the causes of the partial failure of viral-based therapeutic trials.
ADENOVIRUS AS A VECTOR FOR ONCOLYTIC THERAPY IN MULTIPLE MYELOMA
Adenoviruses are nonenveloped, dsDNA viruses that, in nature, infect cells by binding the fibrous knob of the cox- sackie and adenovirus receptor (CAR) expressed on the sur- face of target cells [11]. As vectors for oncolytic therapies, these viruses have many advantages over other vectors, in- cluding the capability of transducing and replicating in divid- ing as well as non-dividing cells, the ease of manipulation, and a naturally lytic replication cycle, highlighting the use- fulness of these viruses for in vitro production and in vivo curative effects (viruses naturally increase the “dosage” while replicating) [12]. As vectors, adenoviruses have a relatively good safety profile and result in a greater survival rate than other therapeutic vectors. Successful treatment of a variety of tumors using adenoviruses has been demonstrated, and they are highly efficient at in vivo gene delivery [11]. Since 1993, more than 300 clinical trials based on adenoviral vectors have been performed [11], with promising outcomes (Supplementary Table 1 and http://www.clinicaltrials.gov/).
Although the first clinical results of trials based on adenovirus as an oncolytic therapeutic agent have been promising, show- ing clinical safety and the feasibility of the approach, studies have also revealed that tumors can acquire a resistance against this type of therapy, and the efficacy of adenoviral treatment still needs to be improved [13]. Since in vitro adenovirus serotype 5 (Ad5) infects cells via the CAR receptor, and CAR [14] and most of the hematologic cells do not express high levels of these receptors, Ad5 was not initially consid- ered as a possible candidate for MM treatment. However, with recombinant technology, adenovirus can be redirected by modification of the viral attachment fiber knob [15], making its use in the treatment of hematological malig- nancies possible. MM seems to be a better potential target for this therapy than solid tumors since it is easily accessible, with malignant cells found predominantly in the bone mar- row and blood [10]. Several serotypes of adenovirus may be suitable as vectors for tumor therapies. Among the differ- ent serotypes, Ad5 was the first to be used as a therapeutic alternative for MM in 2007 [16]. In order to investigate
the ability of Ad5 to infect myeloma cells, Senac et al. studied the in vitro permissivity of 2 myeloma cell lines, ALMC-1 and ALMC-2, derived from patient samples and representing 2 different stages of disease [17]. Furthermore, by distinguish- ing tumor cells positive for the expression of the CD138 surface marker from normal bone marrow cells that lack CD138 expression, they confirmed that Ad5 infects a higher ratio of tumor cells in MM. Senac et al. proved that not only Ad5, but also Ad6, Ad26, and Ad48 are capable of infecting and killing the majority of MM cell lines as well as ex vivo primary cells from patients, confirming the feasi- bility of translating adenovirus-based therapies to MM patients. Their data suggested that adenovirus may retain the ability to selectively kill tumor cells while sparing normal bone marrow cells in the context of MM treatment [18].
ADENOVIRUS AS A VECTOR FOR GENE THERAPY IN MULTIPLE MYELOMA
The second described strategy, taking advantage of the characteristics of adenovirus, is based on the insertion of genes of interest into the genome of a modified adenovirus.
In this way, it is possible to use the modified viral particles for delivering genes that are, for example, defective or mu- tated in the tumor [15], codifying for enzymes that can be then used to activate specific drugs [19], or codifying for proteins able to inhibit the tumor growth directly or indirectly by inducing an immune response [20], specifically to MM tumor cells.
An example of the first strategy (i.e., delivery of defective of mutated genes to the tumor) is found in the work of Torturro, who described adenovirus-mediated cytotoxic gene therapy, showing the efficiency of recombinant adenovirus- p53-mediated cytotoxicity in vitro in Burkitt's lymphoma and MM [15]. In their work, they emphasized the importance of CAR expression and cellular signaling pathways in ad- enovirus-mediated cytotoxic therapies for MM and other lymphoproliferative malignancies [15]. Using a similar strat- egy, Ni et al. were able to block proliferation in the MM cell line U266 by expressing the dominant negative inhibitor of κBα (Ad5IκB), blocking in this way the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) acti- vation [21].
As an example of coding for enzymes that activate specific drugs, Teoh et al. studied the ability of adenoviral vectors to deliver the thymidine kinase (tk) gene into MM cells.
This group demonstrated that MM cell lines and MM patient cells express both adenoviral receptors and DF3/MUC1 protein. They hypothesized that the DF3 promoter could be used as a selective promoter to control the expression of therapeutic recombinant genes only in tumor cells [19].
In that study, expression of the tumor-selective promoter DF3/MUC1 was found only in MM-derived cells (MUC-pos- itive) and was absent in hematopoietic progenitor cells (MUC1-negative) [19]. In a combinatory study, the ex- pression of tk in MM potentiated in vivo killing by a combina-
tion of viral therapy with ganciclovir [22].
An example of the third strategy is the work of Fernandes et al., who used a conditionally replicating adenovirus con- taining the CD40 ligand transgene (AdEHCD40L) to demon- strate growth inhibition in MM cells [20]. This strategy was based on previous findings that showed CD40L can directly modulate MM cell growth. Their work has effectively dem- onstrated that the presence of CD40L is associated with viral oncolysis and results in MM growth inhibition by activating cellular apoptosis [23]. Considering these findings, the clin- ical application of AdEHCD40L has been proposed in ex- perimental MM treatments [20]. Furthermore, wild type genes for mutated oncogenes can be introduced. For example, Ren et al. designed a vector combining p53 and im- munomodulatory molecules, including the GM-CSF (cyto- kine granulocyte macrophage colony-stimulating factor) and the costimulatory molecule B7-1 (Ad-p53/GM-CSF/B7-1).
In 2005, they used this strategy to cotransfer those 3 mole- cules into MM cell lines and primary myelomas, demonstrat- ing the feasibility and increased immunogenicity of those treated MM cells [24].
OTHER POSSIBLE VIRUSES EXPLORED AS ALTERNATIVE CURES FOR MM
Since the majority of the population has encountered ad- enovirus at some point in life, and therefore, a rapid humoral immune response versus the virus and the modified agent is generated, the field of viral therapy is also considering other viruses as alternatives for the treatment of MM. Among those, as comprehensively reported by Thirukkumaran and Morris, the VV was first used in 1980 as a virotherapeutic agent in a 67-year-old Japanese patient with IgA MM [25, 26]. Consequently, intravenous injection of the vaccinia strain was found to result in a significant reduction in IgA levels. To date, other clinical trials testing different VV mu- tants, such as JX-594, have been conducted in patients with metastatic liver cancer. This agent has been considered a possible candidate for clinical trials in hematological malig- nancies, including MM [27]. Other clinical trials in MM have been performed using vesicular stomatitis virus (VSV) as oncolytic agent. Data from those trials suggests that genet- ically engineered VSV strains such as VSVΔ51, which has been used in vivo, can be considered as components in poten- tial combination therapies (e.g., with radiation treatment) for MM [28]. Combination therapy with radiation and viral therapy was reported to have good efficacy in a phase I/II clinical trial using reovirus (reolysin) [29-32]. Reovirus was found to be an attractive anticancer therapeutic in further clinical tests for hematological malignancies. Thirukkumaran and Morris anticipated that after encouraging preclinical data is obtained, reovirus will be used in clinical settings in the very near future [10]. Myers et al. reported new ongoing phase I clinical trials with MV for MM. This trial includes a combination of viral and conventional therapies, consisting of intravenous administration of MV-NIS (MV
expressing the sodium-iodide symporter) used with or with- out cyclophosphamide [33]. In this trial, patients are pre- treated with cyclophosphamide 2 days prior to MV-NIS injection. Pretherapy and post-therapy hematological and biochemical parameters are yet to be determined, and an- ti-measles immunity is being monitored. This study also includes serial imagining of virus biodistribution post-123I administration [34]. In unpublished data, Msaouel et al. [35]
described encouraging results of this therapy in MM for future phase II/III clinical trials. The MV-Edm (Edmonston-B vaccine MV) strain has also been shown to have oncolytic activity against MM. In experiments employing a transplant model in immune-deficient mice, using different clinical MM samples, this agent successfully killed MM cells [36].
CVA21 is an another potential purging agent, with an ability to selectively target hematological malignant cells [37]. Its specificity is most likely related to the expression of both intracellular adhesion molecule-1 (ICAM-1) and decay accel- erating factor (DAF) on the surface of target cells [38, 39].
This virus has already been administered to end-stage MM patients without adverse effects [40], but ongoing human trials are still necessary to evaluate the safety of this therapy.
POSSIBLE OBSTACLES TO VIRAL THERAPIES
Immunogenicity of adenovirus
The injection of adenovirus can lead to the activation of innate and adaptive immune responses against the virus itself. In fact, the strong immunogenicity of this virus is considered one of the major limitations for the in vivo use of this agent. Nayak and Herzog provided a comprehensive overview on the interactions between the immune system and adenoviral vectors [41], suggesting that immune re- sponses against adenovirus can be directed either against the viral protein of the capsid, the vector backbone, or its genomic double-stranded DNA, as demonstrated in gene therapy trials testing the inserted gene.
Systemic delivery of adenovirus vectors results in rapid physiological responses that include activation of innate im- munity, induction of cytokines, inflammation, transient liver toxicity, and thrombocytopenia [42]. The innate immune response, through activation of Toll-like receptor (TLR)-2 and TLR-9, stimulates the production of type I interferons (IFNs), resulting in the production of inflammatory cytokines that promote Th1-type immunity with cellular and humoral immune responses [43, 44]. Natural killer (NK) cells are strongly activated by type I IFNs [45] and are known to be mediators of CD4 and CD8 responses. Adenovirus can also induce the innate immune response through MyD88/
TLR-dependent and/or MyD88/TLR-independent pathways in different cell types [46, 47]. Part of viral clearance is due to complement opsonization [44] and the generation of inflammation, especially in patients with pre-existing anti- bodies against adenovirus. Rapid innate activation, as well as the subsequent cytokine storm (IL-6, type I IFNs, RANTES, IL-12 (p40), IL-5, G-CSF, and GM-CSF), stimulate and acti-
vate the adaptive immune system [42]. Type I IFN signaling is important for the production of antibodies against ad- enovirus, and neutralizing antibodies have been found to be effective in blocking innate and adaptive immune re- sponses to the adenovirus.
The generation of humoral immune responses is crucial, since it precludes re-administration of the same serotype.
Moreover, more than 97% of humans have pre-existing anti- bodies against group C adenoviruses as a result of natural infection.
T cells directed against different serotypes have been found in humans. Adenovirus-specific CD4+ T cells recognize con- served epitopes among different serotypes, and it is possible to find these T cells as well pre-activated CD8 cells able to recognize adenoviral epitopes in the circulation of healthy donors. For these reasons, bypassing the immune response to adenovirus seems to be one of the major challenges in the optimization of this novel therapy.
In order to overcome this limitation, several strategies have been utilized, from targeting specific organs, to en- gineering viral envelopes, switching serotypes, or modifying the transgene cassette. Even immune modulation regimens associated with viral therapy can result in immune avoidance of the viral vector and transgene product, and in some cases, tolerance to the therapeutic gene product can be induced.
For example, Mastrangeli et al. showed that the use of sub- group D partially avoided the generation of neutralizing antibodies in a cystic fibrosis trial [48]. Despite the high immunogenicity of adenovirus vectors, which is generally considered a downside in the context of gene therapy, this could possibly prove to be advantageous when developing cancer vaccines since the adenovirus vector may serve as an optimum adjuvant [49].
IFN-STIMULATED GENES, VIRAL STRESS-INDUCED GENES AND THE PHYSIOLOGICAL RESPONSE
TO VIRUSES
Viruses physiologically trigger an immediate antiviral in- nate response that fights viral infection, replication, and spread. In fact, viral pathogens associated molecular patterns (PAMPs) are recognized by TLRs and are activated through IFN regulatory factor (IRF)-3, IRF-5, IRF-7, or NF-κB a transcription factor responsible for the regulation of hun- dreds of viral stress-inducible genes (VSIGs) that code for proteins with antiviral functions. The TLRs specifically in- volved in viral recognition are TLR2, TLR3, TLR4, TLR8, and TLR9 [50].
A similar antiviral status can also be induced in uninfected cells, through viral stress-related products originating from neighboring infected cells [51]. In fact, when a virus infects a cell, IFNs are synthesized and secreted as a first line of defense [52]. Transcriptional activation by IFN proteins bind- ing to their specific cell surface receptors leads to the tran- scription of IFN-stimulated genes (ISGs), whose products inhibit different stages of viral replication [52]. IFN genes
encode for a large family of multifunctional, secreted, small regulatory glycoproteins that have important signaling roles in the innate immune response.
There are 3 main types of IFNs: Type I or ‘viral’ IFNs include IFN-α, IFN-β, IFN-ω, and IFN-τ; type II IFNs include IFN-γ; and type III IFNs, including IFN-λ, are still not well described and have been suggested to be ancestral type I IFNs that also regulate the viral response [53]. Considerable progress has been made in describing the physiological role of IFN signaling components and subsequent antiviral activ- ities [47, 54].
Gene targeting studies have distinguished the 4 main effec- tor pathways of the IFN-mediated antiviral response:
1) myxovirus (Mx) GTPase pathway;
2) 2´,5´-oligoadenylate-synthetase-directed ribonuclease L pathway;
3) protein kinase R pathway;
4) IFN stimulated gene 15 (ISG15) ubiquitin-like pathway.
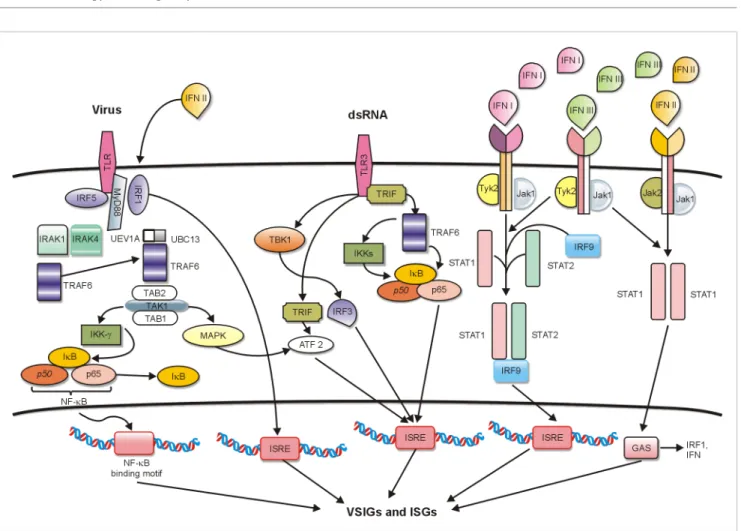
These pathways block viral transcription, degrade viral RNA, inhibit translation, and modify protein function to control each replication step of most viruses [53]. The sets of VSIGs and ISGs that are usually upregulated by viral infection and type I or type II stimulation (Fig. 1) clearly overlap partially [55]. The activation of ISGs promotes the expression of proteins with direct antiviral functions, such as the Mx-resistance-A (MxA) protein that protects infected as well as noninfected bystander cells. MxA proteins are rapidly induced to high levels following IFN or viral exposure and have direct antiviral activity against a wide variety of viruses, including adenovirus [56, 57]. Fig. 1 summarizes the well-studied pathways known to induce upregulation of VSIGs and ISGs.
With the molecular characterization of the transcriptional profiles of many tumors, our group and others have reported the existence of 2 subgroups of cancer cells, distinguishable by a spontaneous activation of the ISG molecular profile independent from viral infection or the presence of IFNs in the microenvironment [58-62]. Analysis of this new ge- nomic data has shown that histologically different cancer types, including pancreatic [58], breast, head and neck, pros- tate, and lung cancer, as well as melanomas and gliomas, generate microarray profiles that identify 2 subgroups dis- tinguishable by specific gene expression of IFNs and in- flammatory chemokines [60-63]. In epithelial ovarian cancer, deregulation of JAK/STAT signaling was identified as a cause of discrimination at the molecular level the 2 different sub- types of tumors characterized by the differential expression of ISGs [64]. Several reports have described some ISGs as markers in solid tumors, both in prognostic and diagnostic contexts. For example, in 2006, Andreu et al. showed that IFTM1, one of the most upregulated ISGs following viral infection, was found to be upregulated downstream of β-cat- enin signal in colorectal tumors [65]. The same marker was found by Gyorffy et al. in ovarian carcinoma, where IFITM1 was actually shown to be associated with therapeutic re- sponses in all treatments analyzed [66]. Weichselbaum et al. proved that, in breast cancer, the VSIG signature is very
Fig. 1. Different signaling pathways leading to the induction of virus stress-inducible genes (VSIGs) and interferon-stimulated genes (ISGs). Left:
virus binding to TLR, stimulating the TLR/MyD88 pathway and leading to the activation and release of NF-κB into nucleus. Center: the dsRNA signaling pathway requires TLR3; there are at least 3 different mechanisms of inducing VSIGs. Right: the JAK/STAT pathway is activated by IFNα, -υ, -β, or -λ; the ISGF3 complex, consisting of STAT1, STAT2, and IRF9, is formed and translocates into nucleus, binding to the ISRE promoter sequence of different ISGs and components of the IFNγ pathway for the activation of GAS promoter sequences. IFNλ mediates the transcription of VSIGs and ISGs through initiation of ISREs or GASs. Abbreviations: TLR, Toll-like receptor; MyD88, myeloid differentiation primary response gene 88; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; IFN, interferon; ISGF, interferon stimulated gamma factor; IRF, interferon regulatory factor; ISRE, interferon stimulated response element; STAT, signal transducer and activator of transcription; GAS, interferon-activated site.
important for DNA damage resistance and therefore can be used as a predictive marker for chemotherapy and radia- tion therapy [61].
While ISG overexpression in solid tumors has previously been described, and several reports have shown that the phenotype of the tumor is dependent on this profile, we were the first to associate this phenotype to an in vitro resistance to oncolytic and gene therapy approaches for pan- creatic cancer [58] and recently for other types of cancers as well, including MM, ovarian cancer, melanoma, RCC, and colon carcinoma (Raus et al., manuscript in preparation).
In fact, we showed that ISG-positive tumor cell lines do not allow either oncolysis or the expression of proteins deliv- ered by adenovirus or adeno-associated virus [58].
In our specific study, we reported for the first time an intrinsic antiviral phenotype in tumor cells that appeared to be independent of the tumor microenvironment, and we performed transcriptional profiling of 3 chronic pancreatitis, 3 primary pancreatic ductal adenocarcinoma (PDAC), 3 paired noncancerous surrounding pancreatic tissues, and 8
PDAC xenografts. We clearly identified 2 distinct PDAC cell phenotypes according to the expression of ISGs, and we found that among several ISGs, 2 phenotypes could be accurately identified by the downstream expression of MxA, which was strongly correlated with the activated antiviral phenotype. MxA, the mediator of one of the first antiviral mechanisms elucidated, is located at a critical intersection of the previously analyzed pathways and is shared by all of these pathways. Therefore, its expression is rapidly in- duced to high levels when IFN or TLR signals occur [56, 67]. We then expanded our analysis to test the level of MxA expression in 23 human PDACs and 10 human PDAC xenografts by tissue array immunohistochemistry, and we observed constitutive expression of MxA in about 50% of samples. This antiviral state is independent from the tumor microenvironment since it could be confirmed by an in vitro model of human tumors that demonstrated resistance to adenoviral replication and lysis [58]. Furthermore, we found that this status is independent from the presence of IFNs and is caused by spontaneous activation of IFN-stimu-
lated response element (ISRE) sequences [58]. Recently, we published an in vivo study where we confirmed that the antiviral MxA-positive phenotype is acquired by ovarian cancer cells in vivo as an escape to oncolytic adenoviral pressure in a murine model [13]. We believe that the ob- servation of this cancer antiviral phenotype might be of practical significance in stratifying patients as likely or un- likely to respond to viral vector-delivered gene therapy;
moreover, it might provide insights into the biology of a subset of human tumors. A thorough understanding of how this antiviral phenotype is generated and if this phenotype may become a new in vivo escape mechanism to viral-based therapies is important in improving the present clinical out- comes of viral therapies.
CONCLUSIONS
Because of the limited efficacy of conventional therapies, new strategies in the treatment of MM are required.
Numerous approaches to novel biological therapies are cur- rently under investigation. Remarkable progress has been made in the field of gene therapy, and, thanks to its great potentiality, the techniques of gene transfer are continuously being improved, attracting increasing interest from clinicians. However, many obstacles still need to be over- come, including improvement of transfection efficiencies, targeting to malignant cells, immune system humoral re- sponse against the inoculated viral particles, possible tumor escape through upregulation of specific gene sets, and possible toxic viral side effects, which are under evaluation in several completed phase I trials. Human clinical trials using ad- enovirus in MM patients have not yet been performed; how- ever, determining the safety of these therapies in phase I and phase II trials will be the first step in translating oncolytic adenoviral therapy to MM patients. Although oncolytic vi- rus-based therapies in MM are only beginning to realize their potential, they appear to be promising future treatments for this disease.
REFERENCES
1. Carli PM, Coebergh JW, Verdecchia A. Variation in survival of adult patients with haematological malignancies in Europe since 1978. Eur J Cancer 1998;34:2253-63.
2. Barlogie B, Epstein J, Selvanayagam P, Alexanian R. Plasma cell myeloma-new biological insights and advances in therapy. Blood 1989;73:865-79.
3. Kyle RA, Rajkumar SV. Multiple myeloma. Blood 2008;111:
2962-72.
4. Bhardwaj A, Sethi G, Vadhan-Raj S, et al. Resveratrol inhibits proliferation, induces apoptosis, and overcomes chemoresistance through down-regulation of STAT3 and nuclear factor-kappaB- regulated antiapoptotic and cell survival gene products in human multiple myeloma cells. Blood 2007;109:2293-302.
5. Delforge M, Blade J, Dimopoulos MA, et al. Treatment-related
peripheral neuropathy in multiple myeloma: the challenge continues. Lancet Oncol 2010;11:1086-95.
6. Greipp P. Treatment paradigms for the newly diagnosed patient with multiple myeloma. Semin Hematol 2005;42(4 Suppl 4):
S16-21.
7. Bluming AZ, Ziegler JL. Regression of Burkitt's lymphoma in association with measles infection. Lancet 1971;2:105-6.
8. Naschberger E, Werner T, Vicente AB, et al. Nuclear factor-kappaB motif and interferon-alpha-stimulated response element co-operate in the activation of guanylate-binding protein-1 expression by inflammatory cytokines in endothelial cells. Biochem J 2004;379:409-20.
9. Der SD, Zhou A, Williams BR, Silverman RH. Identification of genes differentially regulated by interferon alpha, beta, or gamma using oligonucleotide arrays. Proc Natl Acad Sci U S A 1998;
95:15623-8.
10. Thirukkumaran CM, Nodwell MJ, Hirasawa K, et al. Oncolytic viral therapy for prostate cancer: efficacy of reovirus as a biological therapeutic. Cancer Res 2010;70:2435-44.
11. Shirakawa T. The current status of adenovirus-based cancer gene therapy. Mol Cells 2008;25:462-6.
12. Pesonen S, Kangasniemi L, Hemminki A. Oncolytic adenoviruses for the treatment of human cancer: focus on translational and clinical data. Mol Pharm 2011;8:12-28.
13. Liikanen I, Monsurro V, Ahtiainen L, et al. Induction of interferon pathways mediates in vivo resistance to oncolytic adenovirus. Mol Ther 2011;19:1858-66.
14. Bergelson JM, Cunningham JA, Droguett G, et al. Isolation of a common receptor for Coxsackie B viruses and adenoviruses 2 and 5. Science 1997;275:1320-3.
15. Turturro F. Recombinant adenovirus-mediated cytotoxic gene therapy of lymphoproliferative disorders: is CAR important for the vector to ride? Gene Ther 2003;10:100-4.
16. Liu J, Li J, Su C, Huang B, Luo S. Soluble Fms-like tyrosine kinase-1 expression inhibits the growth of multiple myeloma in nude mice.
Acta Biochim Biophys Sin (Shanghai) 2007;39:499-506.
17. Arendt BK, Ramirez-Alvarado M, Sikkink LA, et al. Biologic and genetic characterization of the novel amyloidogenic lambda light chain-secreting human cell lines, ALMC-1 and ALMC-2. Blood 2008;112:1931-41.
18. Senac JS, Doronin K, Russell SJ, Jelinek DF, Greipp PR, Barry MA.
Infection and killing of multiple myeloma by adenoviruses. Hum Gene Ther 2010;21:179-90.
19. Teoh G, Chen L, Urashima M, et al. Adenovirus vector-based purging of multiple myeloma cells. Blood 1998;92:4591-601.
20. Fernandes MS, Gomes EM, Butcher LD, et al. Growth inhibition of human multiple myeloma cells by an oncolytic adenovirus carrying the CD40 ligand transgene. Clin Cancer Res 2009;15:
4847-56.
21. Ni H, Ergin M, Huang Q, et al. Analysis of expression of nuclear factor kappa B (NF-kappa B) in multiple myeloma: downre- gulation of NF-kappa B induces apoptosis. Br J Haematol 2001;
115:279-86.
22. Meeker TC, Lay LT, Wroblewski JM, Turturro F, Li Z, Seth P.
Adenoviral vectors efficiently target cell lines derived from selected lymphocytic malignancies, including anaplastic large cell lymphoma and Hodgkin's disease. Clin Cancer Res
1997;3:357-64.
23. Tong AW, Seamour B, Chen J, et al. CD40 ligand-induced apoptosis is Fas-independent in human multiple myeloma cells.
Leuk Lymphoma 2000;36:543-58.
24. Ren SP, Wang LS, Guo Q, et al. Genetically modified myeloma cell vaccine inducing antitumor immune response in vivo. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2006;14:54-60.
25. Thirukkumaran CM, Russell JA, Stewart DA, Morris DG. Viral purging of haematological autografts: should we sneeze on the graft? Bone Marrow Transplant 2007;40:1-12.
26. Kawa A, Arakawa S. The effect of attenuated vaccinia virus AS strain on multiple myeloma; a case report. Jpn J Exp Med 1987;
57:79-81.
27. Park BH, Hwang T, Liu TC, et al. Use of a targeted oncolytic poxvirus, JX-594, in patients with refractory primary or metastatic liver cancer: a phase I trial. Lancet Oncol 2008;9:533-42.
28. Goel A, Carlson SK, Classic KL, et al. Radioiodide imaging and radiovirotherapy of multiple myeloma using VSV(Delta51)-NIS, an attenuated vesicular stomatitis virus encoding the sodium iodide symporter gene. Blood 2007;110:2342-50.
29. Forsyth P, Roldán G, George D, et al. A phase I trial of intratumoral administration of reovirus in patients with histologically con- firmed recurrent malignant gliomas. Mol Ther 2008;16:627-32.
30. Gollamudi R, Ghalib MH, Desai KK, et al. Intravenous admini- stration of Reolysin, a live replication competent RNA virus is safe in patients with advanced solid tumors. Invest New Drugs 2010;28:641-9.
31. Vidal L, Pandha HS, Yap TA, et al. A phase I study of intravenous oncolytic reovirus type 3 Dearing in patients with advanced cancer. Clin Cancer Res 2008;14:7127-37.
32. Harrington KJ, Karapanagiotou EM, Roulstone V, et al. Two-stage phase I dose-escalation study of intratumoral reovirus type 3 dearing and palliative radiotherapy in patients with advanced cancers. Clin Cancer Res 2010;16:3067-77.
33. Myers RM, Greiner SM, Harvey ME, et al. Preclinical pharmacology and toxicology of intravenous MV-NIS, an oncolytic measles virus administered with or without cyclophosphamide. Clin Pharmacol Ther 2007;82:700-10.
34. Dingli D, Peng KW, Harvey ME, et al. Image-guided radiovirotherapy for multiple myeloma using a recombinant measles virus expressing the thyroidal sodium iodide symporter.
Blood 2004;103:1641-6.
35. Msaouel P, Dispenzieri A, Galanis E. Clinical testing of engineered oncolytic measles virus strains in the treatment of cancer: an overview. Curr Opin Mol Ther 2009;11:43-53.
36. Peng KW, Donovan KA, Schneider U, Cattaneo R, Lust JA, Russell SJ. Oncolytic measles viruses displaying a single-chain antibody against CD38, a myeloma cell marker. Blood 2003;101:2557-62.
37. Au GG, Lincz LF, Enno A, Shafren DR. Oncolytic coxsackievirus A21 as a novel therapy for multiple myeloma. Br J Haematol 2007;137:133-41.
38. Rogers JM, Diana GD, McKinlay MA. Pleconaril. A broad spectrum antipicornaviral agent. Adv Exp Med Biol 1999;458:
69-76.
39. Rotbart HA. Antiviral therapy for enteroviral infections. Pediatr Infect Dis J 1999;18:632-3.
40. Surah S, O'Shea S, Dunn H, et al. Utilization of HIV point-of-care
testing clinics in general practice and genitourinary medicine services in south-east London. Int J STD AIDS 2009;20:168-9.
41. Nayak S, Herzog RW. Progress and prospects: immune responses to viral vectors. Gene Ther 2010;17:295-304.
42. Seiler MP, Cerullo V, Lee B. Immune response to helper dependent adenoviral mediated liver gene therapy: challenges and prospects. Curr Gene Ther 2007;7:297-305.
43. Yamaguchi T, Kawabata K, Koizumi N, et al. Role of MyD88 and TLR9 in the innate immune response elicited by serotype 5 adenoviral vectors. Hum Gene Ther 2007;18:753-62.
44. Appledorn DM, McBride A, Seregin S, et al. Complex interactions with several arms of the complement system dictate innate and humoral immunity to adenoviral vectors. Gene Ther 2008;15:
1606-17.
45. Liu JM, Liu ZB, Zhu GH, et al. Determination of trace glucose and forecast of human diseases by affinity adsorption solid substrate- room temperature phosphorimetry based on triticum vulgaris lectin labeled with dendrimers-porphyrin dual luminescence molecule. Talanta 2008;74:625-31.
46. Muruve DA, Petrilli V, Zaiss AK, et al. The inflammasome recognizes cytosolic microbial and host DNA and triggers an innate immune response. Nature 2008;452:103-7.
47. Nociari M, Ocheretina O, Schoggins JW, Falck-Pedersen E.
Sensing infection by adenovirus: Toll-like receptor-independent viral DNA recognition signals activation of the interferon regulatory factor 3 master regulator. J Virol 2007;81:4145-57.
48. Mastrangeli A, Harvey BG, Yao J, et al. "Sero-switch" adenovirus- mediated in vivo gene transfer: circumvention of anti-adenovirus humoral immune defenses against repeat adenovirus vector administration by changing the adenovirus serotype. Hum Gene Ther 1996;7:79-87.
49. Dharmapuri S, Peruzzi D, Aurisicchio L. Engineered adenovirus serotypes for overcoming anti-vector immunity. Expert Opin Biol Ther 2009;9:1279-87.
50. Boo KH, Yang JS. Intrinsic cellular defenses against virus infection by antiviral type I interferon. Yonsei Med J 2010;51:9-17.
51. Sarkar SN, Sen GC. Novel functions of proteins encoded by viral stress-inducible genes. Pharmacol Ther 2004;103:245-59.
52. zum Büschenfelde CM, Metzger J, Hermann C, Nicklisch N, Peschel C, Bernhard H. The generation of both T killer and Th cell clones specific for the tumor-associated antigen HER2 using retrovirally transduced dendritic cells. J Immunol 2001;167:
1712-9.
53. Sadler AJ, Williams BR. Interferon-inducible antiviral effectors.
Nat Rev Immunol 2008;8:559-68.
54. Sung RS, Qin L, Bromberg JS. TNFalpha and IFNgamma induced by innate anti-adenoviral immune responses inhibit adenovirus- mediated transgene expression. Mol Ther 2001;3:757-67.
55. Wacher C, Muller M, Hofer MJ, et al. Coordinated regulation and widespread cellular expression of interferon-stimulated genes (ISG) ISG-49, ISG-54, and ISG-56 in the central nervous system after infection with distinct viruses. J Virol 2007;81:860-71.
56. Staeheli P, Pavlovic J. Inhibition of vesicular stomatitis virus mRNA synthesis by human MxA protein. J Virol 1991;65:
4498-501.
57. Krug RM, Shaw M, Broni B, Shapiro G, Haller O. Inhibition of influenza viral mRNA synthesis in cells expressing the
interferon-induced Mx gene product. J Virol 1985;56:201-6.
58. Monsurró V, Beghelli S, Wang R, et al. Anti-viral state segregates two molecular phenotypes of pancreatic adenocarcinoma:
potential relevance for adenoviral gene therapy. J Transl Med 2010;8:10.
59. Marincola FM, Wang E, Herlyn M, Seliger B, Ferrone S. Tumors as elusive targets of T-cell-based active immunotherapy. Trends Immunol 2003;24:335-42.
60. Tsai MH, Cook JA, Chandramouli GV, et al. Gene expression profiling of breast, prostate, and glioma cells following single versus fractionated doses of radiation. Cancer Res 2007;67:
3845-52.
61. Weichselbaum RR, Ishwaran H, Yoon T, et al. An interferon- related gene signature for DNA damage resistance is a predictive marker for chemotherapy and radiation for breast cancer. Proc Natl Acad Sci U S A 2008;105:18490-5.
62. Martin DN, Boersma BJ, Yi M, et al. Differences in the tumor
microenvironment between African-American and European- American breast cancer patients. PLoS One 2009;4:e4531.
63. Khodarev NN, Minn AJ, Efimova EV, et al. Signal transducer and activator of transcription 1 regulates both cytotoxic and pro- survival functions in tumor cells. Cancer Res 2007;67:9214-20.
64. Konstantinopoulos PA, Spentzos D, Cannistra SA. Gene- expression profiling in epithelial ovarian cancer. Nat Clin Pract Oncol 2008;5:577-87.
65. Andreu P, Colnot S, Godard C, et al. Identification of the IFITM family as a new molecular marker in human colorectal tumors.
Cancer Res 2006;66:1949-55.
66. Györffy B, Dietel M, Fekete T, Lage H. A snapshot of microarray- generated gene expression signatures associated with ovarian carcinoma. Int J Gynecol Cancer 2008;18:1215-33.
67. Hale BG, Randall RE, Ortín J, Jackson D. The multifunctional NS1 protein of influenza A viruses. J Gen Virol 2008;89:2359-76.
Supplementary Table 1. Oncolytic adenoviral therapy trials in solid tumors.
Virus N Disease Phase Route Results References
Onyx-015 23 Pancreatic ca. I Direct inj. 6 TR, 11 SD Hammersmith Hospital, UK
Onyx-015 21 Pancreatic ca. II Direct inj. 2 PTR, 2 MR, 6 SD Jonsson Comprehensive
Cancer Center, LA, USA.
Onyx-015 21 Pancreatic ca. I/II Virus±
gemcitabine No objective response Sangro B, Mazzolini G, Ruiz J et al.
Onyx-015 30 H&N tumors II IT+CTx 8 CR but long-term survival
not improved Ganly et al., 2000
Onyx-015 22 SCCHN I Inj. 3 PR, 2 MR, 8 SD Ganly et al., 2000
Onyx-015 30 SCCHN II Inj. 1 CR, 3 PR, 12 SD Nemunaitis et al., 2001
Onyx-015 10 SCCHN II Inj. 1 CR, 4 SD Nemunaitis et al., 2001
Onyx-015 37 SCCHN II Inj. 8 CR, 11 PR, median survival
10.5 months Khuri et al., 2000
Onyx-015 16 Ovarian ca. I IT 0 P/CR Vasey et al., 2002
Onyx-015 5 Solid tumor I IV 0 P/CR Nemunaitis et al., 2003
Onyx-015 5 Colon ca. I IV 0 P/CR Nemunaitis et al., 2003
Onyx-015 5 Hepatocellular ca. I IV+IT 1 PR Habib et al., 2003
Onyx-015 9 Colon ca. I IHA 1 PR Reid et al., 2001
Onyx-015 10 Solid tumor I IV 0 P/CR, 8 SD Nemunaitis et al., 2001
Onyx-015 23 Pancreatic ca. I IT 0 P/CR, 6 MR, 10 SD Mulvihill et al., 2001
Onyx-015 27 GI ca. liver meta. II IHA 3 PR, 4 MR, 9 SD Reid et al., 2002
Onyx-015 21 Pancreatic ca. I Inj. 2 PR, 2 MR, 6 SD Hecht et al., 2003
Onyx-015 18 Colon ca. II IV+IT 7 SD Hmid et al., 2003
Onyx-015 20 Hepatabilliary II IT 1 PR Makower et al., 2003
Ad-OC-TK 6 Prostate ca. I/II inj. 3 GR Shirakawa et al., 2007
Ad.IL12 21 GI tumor I IT Mild tumor effects Sangro B, Mazzolini G, Ruiz J
Ad.IL12 7 Pancreatic ca. I IT Mild tumor effects Sangro B, Mazzolini G, Ruiz J
Ad. vector 15 Melanoma, meta. I ex vivo/ s.c. 3 PR Schreiber S et al., 1999
Ad. vector 23 Melanoma, meta. I IT 7 LR Stewart AK et al., 1999
Ad. vector 12 Prostate ca. local. I IT+surgery Significant decrease of PSA levels
in most patients Trudel S et al., 2003
Ad. vector 21 NSCLC I IT NR Griscelli F et al., 2003
Ad. vector 21 NSCLC I IT Amelioration of time to disease
progression Roth JA et al., 1998
Ad. vector 24 NSCLC I IT+CTx 2 PR, 17 SD, 4 PD, 1 NE Nemunaitis J et al., 2004
Ad. vector 15 NSCLC I IT 4 SD Schuler M et al., 1998
Ad. vector 25 NSCLC II IT+CTx No significant benefit over
CTx alone Schuler M et al., 2001
Ad. vector 12 Bladder ca. I IT or
intravesical+
Instillation+
cystectomy
Not evaluated Kuball J et al., 2001
Ad. vector 13 Bladder ca. I Intravesical
instillation 2 SD, 11 PD Pagliaro LC et al., 2003 Ad. vector 36 Ovarian ca. I/II IP+CTx >50% reduction of CA125 levels
in 8; 10 long-term survivors after treatment with multiple doses
Buller RE et al., 2002;
Pegram M et al., 2002
Ad. vector 27 NSCLC I IT 2 PR, SD in 16, 7 PD Swisher SG et al., 1999
Ad. vector 19 NSCLC II IT+RTx 1 CR, 11 PR, 3 SD, 2 PD, 2 NE Swisher SG et al., 2003 Ad. vector 15 Glioma I IT+surgery Mean progression-free survival
13 wk, median overall survival 43 wkLang FF et al., 2003
Ad. vector 15 Ovarian ca. I IP 5 SD, 8 PD Alvarez RD et al., 2000
Ad. vector 13 Glioblastoma I IT 3 SD Trask TW et al., 2000
Ad. vector 14 Glioblastoma I/II IT 1 CR, 5 PR, 4 SD, 4 PD Sandmair AM et al., 2000
Ad. vector 14 Glioblastoma I IT+surgery No clinical response Smitt PS et al., 2003
Abbreviations: CR, complete response; PR, partial response; MR, minor response; SD, stable disease; PD, progressive disease; H&N, head and neck; SCCHN, squamous cell carcinoma of the head and neck; NSCLC, non-small-cell lung cancer; HCC, hepatocellular cancer; Ad., Adenoviral; CTx, chemotherapy; RTx, radiotherapy; TR, tumor regression; PTR, partial tumor regression; GR, good response; LR, local response; PSA, prostate specific antigen; ATAP, Advanced Therapy Access Program; inj., injection; IT, intratumoral; IV, intravenous; IP, intraperitoneal; IHA, intrahepatic artery.
Supplementary Table 1. Continued.
Virus N Disease Phase Route Results References
Ad. vector 18 Prostate ca. I IT PSA response in 1 Herman JR et al., 1999
Ad. vector 30 Prostate ca. I/II IT+RTx Not reported Teh BS et al., 2001
Ad. vector 11 Prostate ca. I IT 1 SD Kubo H et al., 2003
Ad. vector 22 H&N ca. I IT 3 PR Ganly I et al., 2000
Ad. vector 3 HCC I IT Not reported Habib NA et al., 2001
Ad. vector 6 Colon ca.
liver meta. I IHA or IV Not reported Habib NA et al., 2001
Ad. vector 7 Liver meta. from colon ca. or unknown primary
II IHA+CTx 6 SD, 1 PD Habib NA et al., 2001
Ad. vector 5 HCC II IV+IT 1 PR, 4 PD Habib NA et al., 2002
Ad. vector 23 Pancreatic ca. I IT 6 MR, 10 SD Mulvihill S et al., 2001
Ad. vector 10 Meta. solid tumors I IT+CTx 1 MR, 8 SD Nemunaitis J et al., 2001
Ad. vector 16 Ovarian ca. I IP 4 SD Vasey PA et al., 2002
Ad. vector 21 Pancreatic ca. I/II IT+CTx 2 PR, 2 MR, 6 SD, 11 PD Hecht JR et al., 2003
Ad. vector 37 H&N ca. II IT+CTx 8 CR, 1 PR Khuri FR et al., 2000
Ad. vector 40 H&N ca. II IT 3 CR, 2 PR, 3 MR, 13 SD, 15 PD Nemunaitis J et al., 2001 Ad. vector 27 GI ca. liver meta. II IHA+CTx 3 PR, 4 moderate response
(including 3 CTx resistant tumors), 9 SD, 11PD
Reid T et al., 2002
Ad. vector 20 Prostate ca. I IT Reduction in serum PSA in 13
(including 4 PR) DeWeese TL et al., 2001
Ad. vector 16 Prostate ca. I IT 7 MR, 3 PR Freytag SO et al., 2002
Ad5-Δ24-
RGD 21 Ovarian ca. I IP 14 SD, 5 PD, 0 P/CR Kimball KJ et al., 2010
CG7870 23 Hormone
refractory I IV 5 declain in PSA Small EJ et al., 2006
Ad5-D24-
RGD 8 Advanced solid
tumors ATAP IT+IV 3 showed progress Pesonen S et al., 2010
Ad5/3-
Cox2L-D24 14 Advanced solid
tumors ATAP IT+IV 10 showed progress Hemminki A et al., 2010
H103 27 Advanced solid
tumors I IT 13 (PR/MR/SD) Li JL et al.
KH901 19 H&N ca. I IT 12 SD Chang J et al., 2009
Ad5/3-D24-
GMCSF 12 Advanced solid
tumors ATAP IT+IV 8 (MR/SD) Koski A et al., 2010
Ad5-D24-
GMCSF 16 Advanced solid
tumors ATAP IT+IV 8 (CR/MR/SD) Cerullo V et al., 2010
Abbreviations: CR, complete response; PR, partial response; MR, minor response; SD, stable disease; PD, progressive disease; H&N, head and neck; SCCHN, squamous cell carcinoma of the head and neck; NSCLC, non-small-cell lung cancer; HCC, hepatocellular cancer; Ad., Adenoviral; CTx, chemotherapy; RTx, radiotherapy; TR, tumor regression; PTR, partial tumor regression; GR, good response; LR, local response; PSA, prostate specific antigen; ATAP, Advanced Therapy Access Program; inj., injection; IT, intratumoral; IV, intravenous; IP, intraperitoneal; IHA, intrahepatic artery.