34 책임저자:김종석, 560-180, 전주시 덕진구 금암동 산 2-20
전북대학교 의학전문대학원 생화학교실 Tel: 063-270-3085, Fax: 063-274-9833 E-mail: [email protected]
접수일:2010년 2월 16일, 1차수정일: 2010년 2월 19일, 2차수정일: 2010년 2월 23일, 게재승인일:2010년 2월 25일
Correspondence to:Jong-Suk Kim
Department of Biochemistry and Institute for Medical Sciences, Chonbuk National University Medical School, San 2-20, Geumam-dong, Deokjin- gu, Jeonju 560-180, Korea
Tel: +82-63-270-3085, Fax: +82-63-274-9833 E-mail: [email protected]
Ethanol Upregulates PTEN via Nuclear Factor-kappa B (NF-κB) Signaling Pathways in HL-60 Cells
Young-Ju Jeong1 and Jong-Suk Kim2
Departments of 1Obstetrics & Gynecology, 2Biochemistry and Institute for Medical Sciences, Chonbuk National University Medical School, Jeonju 560-180, Korea
The phosphatase tensin homolog deleted on chromosome 10 (PTEN) has various cellular functions including cell growth, proliferation, migration and cell survival. PTEN also plays an important role in ethanol-induced cytotoxicity. The present study was performed to determine the role of ethanol in PTEN expression of HL-60 cells. Here we show that ethanol up-regulates PTEN via activation of nuclear factor-kappa-light-chain-enhancer of activated B (NF-κB) in HL-60 cells. Western blot and RT-PCR results revealed that ethanol increased PTEN expression at HL-60 cells in a dose-dependent manner. In addition, we found that ethanol activated the NF-κB pathways, evidenced by NF-κB DNA binding activity in ethanol-treated cells. However, ethanol-induced PTEN expression was completely blocked by disruption of NF-κB with pyrrolidine dithiocarbamate (PDTC). We conclude that ethanol induces up-regulation of PTEN expression through NF-κB activation in HL-60 cells. (Cancer Prev Res 15, 34-38, 2010)
Key Words: PTEN, NF-κB, Ethanol, HL-60 cells
INTRODUCTION
The novel tumor suppressor gene, PTEN/MMAC1/TEP1, on the chromosome 10q23 protein product (PTEN), is a lipid phosphatase.1∼3) Phosphatidylinositol 3-kinase (PI3K) is the central enzyme of an intracellular signaling pathway that promotes the survival and proliferation, and differentiation.4∼6) The function of PI3K can be antagonized by PTEN that dephosphorylates the main PI3K product phosphatidylinositol 3,4,5-triphosphate (PIP3).1,4∼6) Many reports have shown that PTEN has various cellular function including cell growth, proliferation, migration and cell survival.5∼7)
Recently, it has been shown that PTEN expression is associated with neuronal differentiation,8) and in particular, the PTEN induced G1 phase cell cycle arrest in several cell lines.9) HL-60 cells, a human promyelocytic leukemic cell line, control
the progression of its cell cycle through the activation of the PI3K signaling pathway.10) Recently, it has been shown that PTEN induced G1 phase cell cycle arrest in HL-60 cells.11∼14) In addition, dimethyl sulfoxide (DMSO), a representive HL-60 differentiation agent, could affect the PTEN signaling path- way.13) These findings indicate that the differentiation power of HL-60 cells is dependent on their PTEN expression ability.
On the other hand, ethanol induces cellular injury through its negative action on PI3K signaling pathway.15) In fact, it has been known that cellular levels of PTEN account for the sensitivity of ethanol-exposed cells to cytokine-induced cyto- toxicity.16) These findings indicate that ethanol could control PTEN expression in mammalian cells. In this study, we therefore investigated whether ethanol could regulate PTEN in HL-60 cells. We found that ethanol induced expression of PTEN through NF-κB activation in HL-60 cells.
MATERIALS AND METHODS 1. Materials
Anti-PTEN monoclonal antibody and p85 antibody were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). M-MLV RNase H-reverse transcriptase, TRIzol reagent, RPMI 1640, and fetal calf serum were obtained from Gibco- BRL (Gaithersburg, MD, USA). HBSS (Hanks balanced salt solution), ethanol, anti-β-actin antibody, and PDTC were obtained from Sigma Chemical Co. (St. Louis, MO, USA).
2. Cell culture
A human leukemia cell line (HL-60) was obtained from the American Type Culture Collection (Rockville, MD, USA). The cells were cultured in RPMI 1640 medium containing 10%
fetal calf serum, 2 mM glutamine, antibiotics (Penicillin G 60 mg/L, Streptomycin 100 mg/L, Amphotericin B 50μl/L) under a humid atmosphere (5% CO2, 95% air).
3. Reverse transcription polymerase chain re- action (RT-PCR) for PTEN expression
To examine the contribution of transcriptional control in PTEN regulation, RT-PCR was performed using RNA PCR Kits (GeneAmp, Applied Biosystems, USA). Total RNA was isolated from cells using TRIzol reagent following the manu- facturer’s instructions. Five micrograms of total RNA was transcribed into cDNA in a 20μL final volume of reaction buffer (10 mM Tris-HCl, 50 mM KCl, 1.5 mM MgCl2, 1 mM each dNTP) and 2.4 M oligo-d (T)16-primer, 1 units RNase inhibitor, and 2.5 units M-MLV RNase H-reverse transcriptase by incubation for 15 minutes at 70oC and 50 minutes at 42oC.
The reaction was stopped by incubation at 95oC for 10 minutes. PCR aliquots of the synthesized cDNA were added to a 45μL PCR mixture containing 10 mM Tris-HCl, 50 mM KCl, 1.5 mM MgCl2, 0.2 mM each dNTP, and 2 units Taq DNA polymerase, and 0.4μM of each PCR primer: sense primer, human PTEN (5'-CCGGAATTCATGACAGCCAT- CATCAAAGA-3'), and antisense primer, human PTEN (5'- CGCGGATCCTCAGACTTTTGTAATTTGTG-3'). Amplifica- tion for PTEN or (PPAR) was initiated with 3 minutes of denaturation at 94oC followed by 26 cycles at 94oC for 1 minute, 94oC for 1 minute, 55oC for 1 minute, and 72oC for 3 minutes. After the last cycle of amplification, the samples
were incubated for 5 minutes at 72oC. β-actin PCR was performed with 2.5μL aliquots of synthesized cDNA using primers at a concentration of 0.15μM: sense primer, human β-actin (5'-CCACGAAACTACCTTCAACTCC-3'), antisense primer (5'-TCATACTCCTGCTGCTTGCTGATCC-3'). The ob- tained PCR products were analyzed on ethidium bromide- stained agarose gels (2%).
4. Western blot analysis for PTEN
Human leukemia cells (1×106 cells) were seeded in 100-mm culture dishes and harvested in phosphate buffered saline (PBS).
After washing with PBS, cell pellets were lysed with lysis buffer (20 mM Hepes pH 7.2, 1% Triton X-100, 150 mM NaCl, 0.1 mM phenylmethylsulfonyl fluoride (PMSF), 1 mM EDTA, 1μg/mL aprotinin). After incubation for 30 minutes at 4oC, cellular debris was removed by centrifugation at 100,000 g for 30 minutes, and supernatants were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The protein concentration was determined using protein assay kits (Bio-Rad Laboratories, USA). Samples (50μg) were prepared with four volumes of 0.5 M Tris buffer (pH 6.8) containing 4% SDS, 20% glycerol and 0.05% bromophenol blue at 95oC for 5 minutes. SDS-PAGE was performed in 10% slab gels.
Proteins were then transferred to nitrocellulose paper. The membrane was washed in blocking buffer (10 mM Tris-HCl pH 8.0, 150 mM NaCl, 5% fat-free milk) for 60 minutes at room temperature with shaking and then washed with TBST (TBS, 0.01% Tween 20). Blots were incubated with the primary antibodies against PTEN or p65 followed by alkaline phosphatase-conjugated secondary antibody.
5. Preparation of nuclear extract
HL-60 cells (2×106 cells) were treated with 50 mM ethanol for 24 h. Cells were immediately washed twice, scraped into 1.5 ml of ice-cold PBS (pH 7.9), and then pelleted at 12,000×g for 30 s. Cytoplasmic and nuclear extracts were prepared from cells using the NE-PERⓇ Nuclear and Cyto- plasmic Extraction Reagents (Pierce Biotechnology, Rockford, IL).
6. Electrophoretic mobility shift assay (EMSA)
The activation of NF-κB was assayed by a gel mobility shift assay using nuclear extracts. An oligonucleotide containing the κ-chain binding site (κB, 5’-CCGGTTAACAGAGGG-
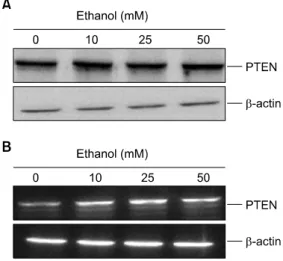
Fig. 1. Effect of ethanol on the PTEN expression in human leukemia cells. HL-60 cells (1×106) were stimulated with various concentrations of ethanol for 24 h. Western blot (A) and RT-PCR (B) for PTEN expression was performed as described in “Materials and Methods”. β-actin was used as a loading control.
Fig. 2. Effect of ethanol on the pAKT expression in human leukemia cells. HL-60 cells (1×106) were stimulated with various concentrations of ethanol for 24 h. Western blot for pAKT expression was performed as described in “Materials and Methods”. β-actin was used as a loading control.
Fig. 3. Effect of ethanol on the NF-κB activation in HL-60 cells. HL-60 cells (1×106) were stimulated with 50 mM ethanol for 24 h. Nuclear extracts were prepared as described in Materials and Methods. NF-κB DNA binding was analyzed by electrophoretic mobility shift analysis as described in Materials and Methods.
GGCTTTCCGAG-3’) was synthesized and used as a probe for the gel retardation assay. The two complementary strands were then annealed and labeled with [α-32P]dCTP. Labeled oli- gonucleotides (10,000 cpm), 10μg of nuclear extracts, and binding buffer (10 mM Tris-HCl, pH 7.6, 500 mM KCl, 10 mM EDTA, 50% glycerol, 100 ng poly (dIㆍdC), 1 mM DTT) were then incubated for 30 min at room temperature in a final volume of 20μl. The reaction mixtures were analyzed by electrophoresis on 4% polyacrylamide gels in 0.5X Tris-borate buffer, and the gels were then dried and examined by autoradiography.
RESULTS
1. Ethanol induces up-regulation of PTEN ex- pression in HL-60 cells
HL-60 cells, a human promyelocytic leukemic cell line, control the progression of its cell cycle through the activation of PI3K/Akt signaling pathway.14) To address such a possi- bility, HL-60 cells were treated with various concentrations of ethanol (0∼50 mM) for 24 h, and then RT-PCR and western blot analysis was performed. Ethanol caused an increase of PTEN protein level in a dose-dependent manner (Fig. 1A). At the same time, we also determined the mRNA levels of PTEN.
As shown at Fig. 1B, PTEN mRNA level in leukemia cells
was also increased after treatment of ethanol.
An increased expression of PTEN could induce the inhibition of Akt phosphorylation. Therefore, Western blot was perform- ed to detect the phosphorylated Akt in the ethanol- treated HL-60 cells (Fig. 2). This reduction in phosphorylated Akt was correlated with the increase in PTEN expression. Thereby, we confirmed that PTEN activity was enhanced within ethanol-treated leukemia cells. These findings suggest that ethanol upregulates PTEN expression in HL-60 cells.
Fig. 4. Effect of PDTC on ethanol-induced PTEN expression in human leukemia cells. HL-60 cells (1×106) were stimulated with 50 mM ethanol in the presence of 10μM PDTC. Cell extraction and western blot for PTEN level were performed as described in “Materials and Methods”. β-actin was used as a loading control.
2. Ethanol induces NF-κB activation in HL-60 cells
Our previous results show that PTEN expression is enhanced during DMSO-induced NF-κB activation in HL-60 cells.13) To clarify the mechanism of ethanol-mediated up-regulation of PTEN expression, ethanol-induced activation of NF-κB was evaluated using EMSA. As shown in Fig. 3, ethanol increased NF-κB binding activity. These results suggest that ethanol potentially induces NF-κB activation in HL-60 cells.
3. NF-κB activation is essential for ethanol- induced upregulation of PTEN in HL-60 cells
In order to determine definite role of NF-κB in ethanol- induced upregulation of PTEN, pyrrolidine dithiocarbamate (PDTC), a chemical inhibitor of NF-κB was used to inhibit the activation of NF-κB in leukemia cells treated with ethanol. HL-60 cells (1×106 cells) were treated with 50 mM ethanol in the presence of 10μM PDTC for 24 h. PDTC blocked the stimulation of PTEN expression by ethanol (Fig.
4). In addition, PTEN levels of leukemia cells in the presence of PDTC were even less than those of control cells. These results show that ethanol-induced PTEN expression is de- pendent on NF-κB activation, and NF-κB is consequently a key regulator of the upregulation of PTEN by ethanol.
However, the role of NF-κB in PTEN expression is con- troversial; some results suggested that NF-κB inhibits PTEN expression in the human colon or cervix cancer cell lines.17) Our previous finding suggested that NF-κB activity is positive regulator of PTEN expression in leukemia cells.13) Thus, these findings indicate that the profile of PTEN expression controlled
by NF-κB is different by cell natures.
DISCUSSION AND CONCLUSION
In conclusion, we found that ethanol induces NF-κB- dependent PTEN expression in leukemia cells. Our result is the first evidence that PTEN is target of ethanol/NF-κB pathways in leukemia cells. This paper might help us to understand the mechanism by which ethanol induces differentiation of leu- kemia cells.
REFERENCES
1) Myers MP, Pass I, Batty IH, Van der Kaay J, Stolarov JP, Hemmings BA, Wigler MH, Downes CP, Tonks NK. The lipid phosphatase activity of PTEN is critical for its tumor supressor function. Proc Natl Acad Sci U S A 95, 13513- 13518, 1998.
2) Tonks NK, Myers MP. Structural assets of a tumor sup- pressor. Science 286, 2096-2097, 1999.
3) Wen S, Stolarov J, Myers MP, Su JD, Wigler MH, Tonks NK, Durden DL. PTEN controls tumor-induced angiogenesis.
Proc Natl Acad Sci U S A 98, 4622-4627, 2001.
4) Kim JS, Peng X, De PK, Geahlen RL, Durden DL. PTEN controls immunoreceptor (immunoreceptor tyrosine-based acti- vation motif) signaling and the activation of Rac. Blood 99, 694-697, 2002.
5) Stambolic V, Suzuki A, de la Pompa JL, Brothers GM, Mirtsos C, Sasaki T, Ruland J, Penninger JM, Siderovski DP, Mak TW. Negative regulation of PKB/Akt-dependent cell survival by the tumor suppressor PTEN. Cell 95, 29-39, 1998.
6) Maehama T, Dixon JE. The tumor suppressor, PTEN/
MMAC1, dephosphorylates the lipid second messenger, pho- sphatidylinositol 3,4,5,-triphosphate. J Biol Chem 273, 13375- 13378, 1998.
7) Tang Y, Eng C. PTEN autoregulates its expression by stabilization of p53 in a phosphatase-independent manner.
Cancer Res 66, 736-742, 2006.
8) Lachyankar MB, Sultana N, Schonhoff CM, Mitra P, Poluha W, Lambert S, Quesenberry PJ, Litofsky NS, Recht LD, Nabi R, Miller SJ, Ohta S, Neel BG, Ross AH. A role for nuclear PTEN in neuronal differentiation. J Neurosci 20, 1404-1413, 2000.
9) Furnari FB, Huang HJ, Cavenee WK. The phosphoinositol phosphatase activity of PTEN mediates a serum-sensitive G1 growth arrest in glioma cells. Cancer Res 58, 5002-5008, 1998.
10) Cappellini A, Tabellini G, Zweyer M, Bortul R, Tazzari PL, Billi AM, Fala F, Cocco L, Martelli AM. The phosphoinositide 3-kinase/Akt pathway regulates cell cycle progression of HL60 human leukemia cells through cytoplasmic relocalization of the
cyclin-dependent kinase inhibitor p27(Kip1) and control of cyclin D1 expression. Leukemia 17, 2157-2167, 2003.
11) Lee YR, Yu HN, Noh EM, Kim JS, Song EK, Han MK, Kim BS, Lee SH, Park J. Peroxisome proliferator-activated receptor gamma and retinoic acid receptor synergistically up-regulate the tumor suppressor PTEN in human promyeloid leukemia cells. Int J Hematol 85, 231-237, 2007.
12) Lee YR, Yu HN, Noh EM, Youn HJ, Song EK, Han MK, Park CS, Kim BS, Park YS, Park BK, Lee SH, Kim JS. TNF- alpha upregulates PTEN via NF-kappaB signaling pathways in human leukemic cells. Exp Mol Med 39, 121-127, 2007.
13) Lee YR, Shim HJ, Yu HN, Song EK, Park J, Kwon KB, Park JW, Rho HW, Park BH, Han MK, Kim JS. Dimethy- lsulfoxide induces upregulation of tumor suppressor protein PTEN through nuclear factor-kappaB activation in HL-60 cells. Leuk Res 29, 401-405, 2005.
14) Cappellini A, Tabellini G, Zweyer M, Bortul R, Tazzari PL,
Billi AM, Falà F, Cocco L, Martelli AM. The phospho- inositide 3-kinase/Akt pathway regulates cell cycle progression of HL60 human leukemia cells through cytoplasmic relo- calization of the cyclin-dependent kinase inhibitor p27(Kip1) and control of cyclin D1 expression. Leukemia 17, 2157-2167, 2003.
15) Shulga N, Hoek JB, Pasorino JB. Elevated PTEN levels account for the increased sensitivity ethanol-exposed cells to tumor necrosis factor-induced cytotoxicity. J Biol Chem 280, 9416-9424, 2005.
16) He J, Monte S, Wands JR. Acute ethanol exposure inhibits insulin signaling in the liver. Hepatology 46, 1791-1800, 2007.
17) Kim S, Domon-Dell C, Kang J, Chung Dai H, Freund J, Evers BM. Down-regulation of the tumor suppressor PTEN by the tumor necrosis factor-/nuclear factor-κB (NF-κB)- inducing kinase/NF-κB pathway is linked to a default I-κB- autoregulatory loop. J Biol Chem 279, 4285-4291, 2004.