약학회지 제41권 제 6 호 782~788(1997)
Yakhak Hoeji
Vol. 41. No. 6일차 배양 혈관 평활근 세포에서 포도당 농도에 의한 엔도톡신 유도 프로스타글란딘 합성 변화
이수환* • 우현구 • 김지영 ■백은주 • 문창현
아 주 대 학 교 외 과 대 학 생 러 학 교 실
(Received November 10. 1997)
E n h a n c e m e n t o f E n d o t o x i n - i n d u c e d P r o s t a g l a n d i n S y n t h e s i s b y E l e v a t i o n o f G l u c o s e C o n c e n t r a t i o n i n P r i m a r y
C u l t u r e d R a t V a s c u l a r S m o o t h M u s c l e C e l l s
Soo Hwan Lee*, Hyun Goo Woo, Ji Young Kim, Eun Joo Baik and Chang-Hyun Moon
Department of Physiology, School of Medicine, Ajou University, Suwon 442-749, Korea
Abstract— This study was designed to characterize glucose-enhancing effects on endotoxin-induced prostaglandin production in primary cultured rat vascular smooth muscle cells (VSMC). High glucose treatment significantly augmented prostaglandin (PG) synthesis in lipopolysaccharide (LPS)-stimu- lated VSMC and this effect was maxima] at the concentration of 4 mg/ml. It has been reported that in creases in glucose metabolism through sorbitol pathway could alter the cytosolic NADH/NAD"^ ratio an d this change favors de novo synthesis of diacylglycerol (DAG) and, in turn, results in the activation of protein kinase C (PKC) in vascular tissues. Protein kinase C (PKC) inhibitors, staurosporin and H7.
blocked the glucose enhancing effect, and DAG. a PKC activator, significantly increased the PG pro
duction stimuated by LPS. Sodium pyruvate, which can reverse the alteration in cytosolic NADH/NAD^
ratio, reduced the high glucose effect on PG production. And also, zopolrestat. a strong aldose reduc
tase inhibitor, almost completely blocked the augmentation effect of glucose on PG synthesis. Ara
chidonic acid release was significantly increased in high glucose treated group, which implied the in crease in PLA2 activity was gissociated with glucose enhancing effect. Metabloic labeling study clearly showed that de novo synthesis of prostaglandin H synthase-2 (PGHS-2) is greatly increased in high glucose treated group and this was mitigated by the treatment of zopolrestat. Taken together, the ac
tivation of PKC through sorbitol pathway increased the activities of PLA2 and PGHS which resulted in the augmentation in LPS-induced PG production in high glucose treated VSMC.
Keywords □ High glucose. Vascular smooth muscle cells. Endotoxin. Prostaglandin.
E ic o s a n o id
란
p r o s ta g la n d in , th ro m b o x a n e , 15- h y d ro x y e ic o s a te tra e n o ic a c id {H E T E s )등과 같은
a r a c h id o n ic a c id
대사산몰을 통칭하는 말로써
.이들
e ic o s a n o id
류들은 염증 반응
,면역조절 반옹 및 세포 분화
.증식등과 같은 다양한 생체 반응에 관여하고 었는 것으로 알려져 있다
. ^ E ic o s a n o id의 생성은
phos-*^ 본 논문에 관한 문외는 이 저자에게로
( 전화) 0331-219-5043 ( 팩스) 0331-219-5049
p h o lip a s e
에 외해 세포막 인지질로 부터
a ra c h i-d on ic a c id
가 유러됨으로 부터 시작되는데
.이 단계가
e ic o sa n o id ,
특히
p r o s t a g la n d in의 생성에 었어 가장 중요한 조절 단계인 것으로 인식되어져 왔다
.^"'유리된
a r a c h id o n ic a c id
는
p r o s t a g la n d in G / H s y n th a s e( P G H S )
에 외해
P G H 2로 번환되며 이 대사체는
th r o m b o x a n e
을 비롯한 모든
p r o s ta n o id외 전구물질 로 작용하게 된다
.이
P G H S에는 두가지
is o fo rm이 존
재하는 것으로 알려져 있으며
.이 중 최근
S im m o n s등
782
에 외해 발견된
in d u c ib le P G H S (P G H S - 2)는 기존 의
iso fo rm (P G H S - 1)과는 다른 조절기전을 통해 발
현되고 있으며
,"'®*각종 질병의 발현에 있어서 중요한
역할을 할 것으로 추정되고 있다. ®이
is o z y m e은 여 러 세포내,외의 자극에 외해 발현이 증가되는 것으로 알 려져 있는데
.'*^*"*특히 다양한 세포들에서
m ito g e n ic s t im u la t io n외 결과로 나타나는
p r o s ta g la n d in합성
중가와 밀접한 관련이 었는 것으로 보고되는 등
,*"^고
**지금 까지의 연구 결과에 의하면
P G H S가 관여하는 단
계는 이전에 주론 되었던 것과는 달러
p r o s ta g la n d in합성에 있어 가장 중요한 조절 단계일 것으로 추정되고 있다
.따라서 최근에는
P G H S - 1및
P G H S - 2외 생리학 적 기능 및 발현 조절 기전에 대한 연구가 매우 활발히 진행되고 있다
.연구자등은
P G H S is o z y m e들의 생체내 역할에 대 한 일련의 연구를 수행하여 오고 있으며
,이러한 연구의
일환으로 일차 배양 혈관 평활근 세포에
b a c te ria llip o p o ly s a c c h a r id e (L P S )
를 처리한 후
p r o s t a g la n d in
합성능 변화틀 측정한 결과
,시간 경과에 따라
p r o s t a c y c lin
외 합성이 현저히 증가되며
,이는
P G H S - 2
외 선택적 발현 증가에 기인함을 확인한 바 있
다
22 24)또한 동일한 실험 조건하에서 행한 예비 실험
결과
,세포 배앙액의 포도당 농도를 증가 시키면
, en-d o to x in
에 외한
p r o s ta g la n d in합성이 현저허 증가됨 을 관찰할 수 있었다
.이와같은 사실은
p ro s ta c y c lin의 강력한 혈관 확장 작용을 감안할 때 매우 중요한 의미를 내포하고 있는 것으로
,당뇨병 상태에서의
e n d o to x in에 의한
sep tic s h o c k발현증가는
p ro s ta c y c lin의 생 성 증가와 밀접한 관련성이 있을 것으로 추정된다
."""^*이와 관련하여
,배양액중의 포도당 농도를 증가시키는 경우 신장 내피세포나 사구체 세포등에서
P K C가 활성 화되며 이는 당뇨병의 혈관 함병증의 한 원인이 될 수 있다는 보고는
,^'®* P G H S is o z y m e의 발현이
P K C s tim u la to r인
p h o rb o l estei■에 외하여 증가된다는 연 구 결과와*®^더불어
,앞서외 추정에 시사하는 바 크다 할 수 있다
.본 연구 에서는 이러한 추론을 확인하기 위 한 연구외 일환으로
.포도당 농도를 달리한 배양조건하 에서 일차 배양 혈관 평활근 세포에서
p r o s ta g la n d in함성에 머처는
e n d o to x in의 영향을 검토하고
,그 작용
기전의 일단을 검토 하였으며
, p r o s ta g la n d in합성 경 로에 관여하는 효소계에 미처는 영향들을 살펴 보고자 하였다
.실험방법
시약- B a c te ria l lip o p o ly sa cc h a rid e (E . coli 0 1 1 1
:
B 4 ), a s p ir in , a ra c h id o n ic acid, 6- ke to - P G F i„
등은
S ig m a C h e m ic a l C o .
에서
. D ulb ec c c /s M o d ifie d M E M , fe ta l c a lf s e r u m등은
G I B C O사에서
, ['H]-6- k e tc rP G F ja , ( ^ ] - a r a c h id o n ic a c id는
N e w E n g la n d N u c le a r ( N E N )에서
, T R A N ^ S - L A B E L T M은
I C N
사에서 구입하였고
, zo p o lre sta t은
P fiz e r사에서 공급 받았으며
,기타외 시약은
1급 이상의 것을 용하 였다
.혈관평활근세포의분리및배양- S p r a g u ^ D a w - le y
계
(1 8 0 - 1 0 0 g)웅성 랫드률 실혈 처사 시킨후 홍강
을 열어
th o rac ic a o rta롤 무균적으로 적출하여
H a n / s F 1 2
배지
( G IB C O , G r a n d Is la n d , N Y , U S A )에 넣어 이후의 분리 배양에 이용하였다
.혈관 평활근 세
포의 분리 및 배양은
T h y b e rg등의 방법에
®준하여 행
하였다
.무균적 조건 하에서
c o nn e ctive tis s u e와 세동
맥돌을 제거한 후
, a o rta를 길이 방향으로 자른 다옴
,0 .1 % c o lla g e n a s e
가 함유된
H a m 's 12배지에서
1시간 동안 전배앙 하였다
. A o r ta의
in n e r lin in g을
r u b b e r p o lic e m a n으로 긁어 제거한 뒤
, m e d ia l layer-1- 1 m m역 크기로 잘게 세절하고 다시
0 .1 % c o lla g e n a s e롤
o v e r n ig h t처리하였다
.분리된 세포를
3,000 r p m에 서
5분간 원심분러 한 후
10% fe ta l c a lf s e r u m이 함유 된
D ulbeccc/s m o d ifie d e s s e n tia l m e d iu m ( D M E M , G IB C O )에
r e s u s p e n d하여
5% 0 0 / 9 5 % a ir, 37°C조 건에서 배앙하였다
.배지는 일주일에
2번씩 교체하였으 며 본 실험에서는
2회
-3회 계대한 세포를 이용하였다
.실험 개시전
48시간 동안의
s e r u m d e p r iv a tio n을 통해
g ro w th a rre s t
시킨 조건하에서 세포 배양액에
2, 4. 6, 8 m g / m/외
g lucose가 포함되도록한 루
, l ( H i g / m/의
L P S
를 가하여
p r o s ta c y c lin (P G l2)외 생성에 미처는 영 향을 검토하였다
.6-keto-PGF,„ 생성능과 PGHS 활성의측정 -
배 양세 포에
10% fe ta l c a lf s e r u m (F C S , G I B C O B R L , U S A)이 포함된
D M E M배지를 가하고
b a c te r ia l lip o p o ly s a c c h a r id e (L P S , E. coli, 0 1 1 1:
B 4 , 10 [ig/m l. S ig m a , U S A
)와 함께
37°C에서 배양하였다
.일정
시간 동안 배양한 후 배지를 전량 취하여
-20°C에 보관
하였다가
,생성된
6-keto-PGF\a외 양을
r a d i o i m m i r n o a s s a y ( R IA)로 측정하였다
. P G H S의 활성은
F u등
®Vol. 41 No. 6, 1997
0
Fig. 1 — Effects of glucose on endotoxin-induced pros
taglandin production in primaiy cultured rat vas
cular smooth muscle cells.
Cells were preincubated in the absence of serum for 48 hours and further incubated with or without lipopolysachharide (LPS. 10 ug/m/) in 10% fetal bovine serum contained fresh media for 16 hours. Supernatants were collected and the contents of 6-ketcrPGF i were quantified with radioimmunoassay. G l • 1 mg/m/ glucose
;
G2
:
2 mg/m/ glucose;
G4:
4 mg/m/ glucose:
G8:
8 mg/m/ glucose. p<0.05 vs G l+ L P S
작용 특성등에 관한 기초 연구를 수행하였다
.일차 배양
혈관 평활근 세포에
L P S를 처리한 경우 이전의 보고에
서와 마찬가지로
p r o s ta g la n d in합성이 현저허 중가 되었으며,""이러한 증가는 배양액내외 포도당 농도 가 증가될수록 더욱 현저하여 포도당 농도
4 m g / m/의 경우
1 m g /m Z인 경우에 비헤
2배 이상의
p r o s ta g la n d in
이 함성됨을 관찰할 수 었었다
.포도당 농도를
4 m g / m /
이상 증가시
? !경우
,더 이상의
p ro s t a g la n d in합성 증가는 관찰되지 않았다
(F ig . 1).따라 서 이후외 실험에서는 포도당을
1 m g / m Z ( G l)및
4m g / m
;
(G 4)의 농도로 처리하여 수행하였다
. W il-l ia m s
등의 보고^**에 외하면
,고포도당 배지에서 배양한
혈관세포에서는 세포질내의
N A D H /N A E T ^비율이 현 저히 중가하며
,이는 세포내에서의
diac y lg ly cer이의 생합성을 증가시키는 환경을 제공하게 되고
,이에 따라
p r o te in k in a s e C ( P K C
)의 활성화가 초래된다고 알려 져 있다
.따라서 다옴 실험에서는 포도당 농도외 증가에 외한
L P S유도
p r o s ta g la n d in합성 증폭 현상이
P K C
외 활성화에 매개되는지의 여부를 확인하기 위하
여 동일한 실험조건하에서
P K C저해제인
s ta u - ro s p o rin및
H 7을 처리하여 보았다
(F ig . 2). P K C저 해제는 고농도 포도당 처리군에서의
p r o s ta g la n d in증
J. Pharm. Soc. Korea 에 의한 실험 방법에 준하여 즉정하였다
.즉
. 6-keto-P G F V
생성능을 측정하기 위해 배지를 제거한 세포에
.과량의
a ra c h id o n ic a c id (10 |xg/m/, S ig m a , U S A )가
함유된
P B S를 가하여
10분간 배양한 후
,배지틀 취하
여 생성된
6 - ke to - P G F V익 양을 정량함으로써
P G H S활성외 지표로 하였다
.6-keto-PGF,„의정량- 6- k e to - P G F i
„의 정 량은 전 보^*에 기술한
r a d io im m u n o a s s a y방법에 따라 실시
하였으며
6-keto-PGF\„항체는 루이지애나 주립대학
의 황 동호 박사로부터 공급 받아 사용하였다
.Arachidonic acid release - 24 w e ll tis s u e c u lt u re p la te ( N u n c , U S A )
에 배양한 혈관 평활근 세포에
[
크
H ] - a r a c h id o n ic a c id (0 .2 5 |xC i/w ell)를 가하고
20
시 간 동안 배양한 다옴
,수회 세척하였다
.포도당 농 도를 달리한 조건하에서 일정시간 동안 배양한 후
.배지 를 취하여
r a d io a c itiv ity틀 측정하였다
.Metabolic labeling -
혈관 평활근 세포를
6 w e ll tis s u e c u ltu r e p la te (F a lc o n , U S A )에
c o n flu e n t하 도록 배양한 다옴
48시간 동안
s e r u m s ta r v a t io n을 행
한 후
,포도당 농도를 달리한
D M E M배지에서 일정 시
간 동안 배양하였다
. M e th io n in e - fre e D M E M ( G I B C O B R L , U S A)으로 배지를 교환하고 여기에
(® S ]- m e th io n in e (5 0 n C i/m Z , IC N , U S A
)틀 가한 후
37°C
에서
2시간 더 배양하였다
. S o lu b iliz a tio n b uff- e r(5 0 m M T ris, p H 7.4, \% T w e e n 20)틀 가하고 초 음파 분쇄기로 세포틀
s o lu b iliz e한 다옴
10,000 rp m ,4°C
에서
5분간 원심분러 한 후 얻은 상층액에
P G H S -2
항체를 가해
im m u n o p r e c ip ita io n을 행하였다
. R a - d k r l a b e l된
P G H S생외 양을
S D S p o ly a c ry la m id e gel e le c tro p h o re sis를 거쳐
flu o ro g ra p h y에 외해 비 교하였다
.통계-
모든
d a t a는
m e a n ± S E로 나타내었으며
, s t u d e n t V t - t e s t에 의해 유의성을 검정하였다
.결과 및고찰
본 연구에서는 포도당 농도를 달리한 배양조건하에서 세균 감염시 병리학적 제 증상을 발현하는 원인 물질인
b a c te r ia l lip o p o ly s a c c h a r id e (L P S
)이
p r o s ta g la n d in
합성에 미처는 영향을 일차 배양 혈관 평활근 세포
에서 검토함으로써
,당뇨병에서외
sep tic s h o c k증상 발현 증가 현상과외 상관성을 살펴 보고자 하였으며
,그
I 1 -LPS * *
HPS T J
Jlili
784 이수환• 우 현 구 • 김지영• 백은주• 문창현
0
5
0
5 2
1
1
f/Blo8-d£>d.ol-9
J
-LPS +LPS +L+Stau +L+H7 Fig. 2 — Effects of PKC inhibitors on endotx)xin-inducedprostaglandin production in primary cultured rat vascular smooth muscle cells
Cells were preincubated in the absence of serum for 48 hours and further incubated with or without lipopolysachharide (LPS. lO ug/m /) in 10% fetal bovine serum contained fresh media for 16 hours. PKC inhibitors, stau
rosporine (Stau
:
100 ng/m/) and H7 (50 uM ) were added with LPS. Supernatants were collected and the contents of 6-ketcrPGFia were quantified with radioimmunoassay. p<0.01 vs G 4+LPS
폭 현상을 현저히 억제함을 관찰할 수 있었다
.그러나
.저농도 포도당 처리군에서외
L P S작용에는 별다른 영 향이 없거나
( s ta u r o s p o r in처리군
),상대적으로 작은 정도의 억제현상만이
(H 7처리군
)관찰 되었다
.이러한
P K C
의 영향이 전술한 바와 같은 경로를 통한 것인지외
여부를 확인하기 위해
,동일한 실험 조건하에서
d i-acylg ly cerol
을 처리한 결과
,저농도 포도당 처리군에
서의
p r o s ta g la n d in함성은 유의적인 증가를 보였으 나
,고농도 포도당 처러군에서는 통계적으로 유의한 번 화를 보이지 않았다
(F ig . 3).이는
F ig . 1에서 보인 바
와 같이
G 4이상의 포도당 농도에서는 더 이상외
6-k e to - P G F i„
생성 증가가 관찰되지 않는 현상과 일치하
고 있다
.세포질내에 증가된
N A D H / N A i y버율은 세
포질내
p y r u v a te의 농도와 역상관관계가 있는 것으로
보고된 바 있으며,크인 이는
p y r u v a te외
la c ta te로의 대 사과정에서
N A D H가
N A I T로 산화됨에 기인하는 것 으로 알려져 있다
.따라서
,이번에는 배양액중의
r u v a te
농도를 번화 시켰을 경우외
p r o s ta g la n d in합 성에 미치는 영향을 검토하였다
. F ig . 3에 요약한 바와 같이
, s o d iu m p y r u v a te처리에 의 해 고농도 포도당에 의한
p r o s ta g la n d in합성 증폭 현상이 유의적으로 억 제됨을 관찰할 수 있었다
.전술한 바와 같은 세포질내에
Vol. 41, No. 6, 1997
6 1 6 4
Fig. 3 — Effects of diacylglycerol (DAG) a n d pyruvate on endotoxin-induced prostaglandin production in primary cultured rat vascular sm ooth m u s cle cells
Cells were preincubated in the absence of serum for 48 hours and further incubated w ith or without lipopolysachharide (LP S. 10 ug/m l) in 10% fetal bovine serum contained fresh m edia for 16 hours. Diacylglycerol (D A G. 10 uM ) and sodium .pyruvate (Pyr, 11 m M ) were added with LPS. Supernatants were collected and the contents of 6-ketcrPGFia were quan
tified w ith radioimmunoassay. p<0.05
서외
N A D H / N A iy ^비율 번화는 과잉의 포도당이
s o rb ito l p a t h w a y
틀 통해 대사됨에 기인하는 것으로 알려져 있다
. A ld o s e re d u c ta s e저해제는 실험적 당뇨 모델에서 변형된 혈관 반응성을 개선시키는 것으로 보 고된 바 있으며
.이는
sorb ito l p a t h w a y저해를 통한
P K C
활성화 억제 작용에 기인한 것으로 알려져 있다
.본 실험에서는
a ld o s e re d u c ta s e저해제인
zo p o h r e s ta t을 처 리 함으로써
s orb ito l p a t h w a y를 차단한 후
p r o s ta g la n d in
합성에 미치는 영향을 검토하였으며 그
걸과
.고포당 처리군에서의 증폭작용이 현저히 억제됨 을 관찰할 수 있었다
( G l + L P S:
0 . 4 5 ± 0 .1 0 n g / m/;
G 4 + L P S • 1.45± 0.047 n g / m
/;
G 4 + L P S + z o p o lre s ta t:
0 .5 4 ± 0 .0 7 4 n g / m / ) .이상의 실험 걸과들로 부 터 고포도당에 의한
p r o s ta g la n d in합성 증폭 현상은
s orb ito l p a t h w a y
를 통한
P K C외 활성화에 기인함을 확인할 수 있었다
.P r o s ta g la n d in
은 다단계의 효소작용을 거쳐 생합성
되며
.이중
.세포막 인지질로부터의
a r a c h id o n ic a c id유러에 관여하는
p h o s p h o lip a s e A2단계와 유리된
a ra c h id o n ic a c id
를
P G H ^로 대사시 키는
p r o s t a g la n d in H s y n th a s e외 단계가 전체 반응 속도틀 규율하고 있는 것으로 알려져 있다/■인 따라서
.다옴 실험에서는
] +LPS I ^LPS-^DAG I ♦LPS*Pyr
0
5
0
5 2
1
1
{luv6u}o
J9d.0la>l<w하
이수환• 우 현 구 • 김지영• 백은주• 문창현
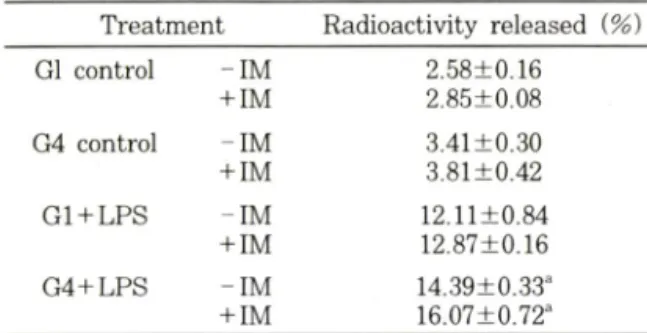
Table I — Effects of glucose on arachidonic acid release from rat vascular smooth muscle cells
T reatm ent Radioactivity released {%)
Gl control - IM 2.58+0.16
+IM 2.85±0.08
G4 control - IM 3.41±0.30
+IM 3.81±0.42
G l+ L P S - IM 12.11+0.84
+IM 12.87±0.16
G 4+ L P S - IM 14.39±0.33"
+IM 16.07±0.72"
고포도당 처리에 의한
p r o s ta g la n d in합성 증폭 현상이 이 두가지 주요 조절 단계에 미처는 영향을 검토하여 보
았다
. C o rn w e ll등®에 외하면 일차 배양 혈관 평활근 세
포에
L P S를 처러한 경우
a ra c h id o n ic ac id release가 현저히 증가되는 것으로 알려져 있으며
,이는 본 실험 결
과에서도 재확인 되고 었었다
(T able I). L P S자국에 의
한
a ra c h id o n ic acid release는 고포도당 처리에 외해 유의적인 증가를 보였으며
.또한 이러한 현상은 유의적
이지는 않았지만
. L P S를 처리하지 않은 군에서도 비숫
한 양상을 보였다
.그러나 이전의 보고와는 달러
, i n d o m e th a c in
처 러에 의한
a ra c h id o n ic acid release차 단 작용은 본 실험에서는 관찰되지 않았다
. L P S에 외한
a ra c h id o n ic acid release
증가는 혈관 평활근 세포에서
외
secretory P L A2활성 중가에 기인하는 것으로 보고
되고 있으나
,본 실험 결과만으로는 고포도당에 의한
a ra c h id o n ic acid release
중가현상이
sec즉
, S ecretory retory P L A2
활성 증가와 관계 있을 것으로 추
정할 수는 없었다
.즉
,전슬한 바와 같이
.고포도당에 외 한
p r o s ta g la n d in생성 증폭 현상이
sorbitol p a th -w a y
를 통한
P K C의 활성화에 기인한다면
,이로 인한
cytosolic P L A2
의 활성화 역시 관여될 가능성을 배제할
수 없기 때문에,이러한 문제점둘에 대해서는 추후 진 전된 연구를 통해 확인되어져야 할 것이다
. L P S에 의하
여 유도되는
P G H S - 2의 생합성에 머처는 포도당의 영
향을 검토한 결과는
F ig. 4에 나타낸 바와 같으며
b a n d
외 상대적 강도는
G 1 + L P S:
100. G 4 + L P S:
178
및
G 4 + L P S + Z o p o lre s ta t:
136이었다
.일반적으 로
P G H S - 2는
m ito ge n ic s tm u li에 의해 발현이 증가되
는 것으로 알려겨 있으며
,"'®본 실험 결과에서도 이를
재확인 할 수 있었다
.즉
, fe ta l c a lf s e r u m에 외해 유도 되는
P G H S - 2외 생합성은
L P S처리에 의해 현저히 증 폭되고 있으며
,이는 고포도당 처리군에서 더욱 현저하
Fig. 4 — Effects of glucose and zopolrestat on endotoxin- induced de novo synthesis of prostaglandin H2
synthase 2 in primary cultured rat vascular smooth muscle cells.
Cells were preincubated in the absence of serum for 48 hours and further incubated with or without lipopolysachharide (LPS. 10 in 10% fetal bovine serum contained fresh media for 12 hours. Medium was changed with methionine free D M E M and further incubated in the presence of [^]-methionine for 4 hours.
Zopolrestat (50 ^M ) was added at the beginning of second incubation. Cell lysates were im- munoprecipitated with specific PGHS-2 antibody and committed to electrophoresis. Fluorographic images were developed on Kodak X - O M at film.
Lane 1 '■ G l control. Lane 2
:
G l LPS treated.Lane 3
:
G l LPS and zopolrestat treated. Lane 4:
G4 control. Lane 5;
G4 LPS treated. Lane 6 ■ G4 LPS and zopolrestat treated.였다
.이 때
. aldose red uc ta se in h ib ito r인
zopol-r e s ta t
을 처리한 경우 고포도당 처리군에서 더욱 증폭되
었던
P G H S - 2의 생합성은 감소됨을 관찰할 수 있었으
며
.이러한 결과는
P G H S활성을 측정한 결과와도 일처
하였다
(G1 + L P S:
9 8 ± 5 4;
G 4 + L P S:
390± 14 0:
G 4 + L P S + z o p o lre s ta t:
1 7 0 ± 7 4: 단위 :
pg/m Z/10 m in ) .그러나 저포도당 처러군에서는
aldose re d u c ta se저해제 처러에 의해 오히려
P G H S - 2생함성이 증
가되는 양상을 보였으며
.이는
P G H S활성 측정 결과와
는 일치하지 않았다
(3 4 ± 1 8 p g /m //1 0 m in ).이러한 불 일처에 대해서는
tim e course s tu d y등을 비롯한 보*다 정밀한 실험을 통하여 그 이유를 추적할 예정이다
.이상 의 결과로 부터 고포도당에 의한
p r o s ta g la n d in합성 증폭작용은
P K C활성화에 따른
PLA2의 활성 중가 및
P G H S - 2
외 생합성 증가에 기인하는 것으로 추정할 수
있었다
.감사의 글
이 논문은
1996년도 한국학슬진흥재단의 공모과제
/.
Pharm. Soc. Korea
연구비
(04-FM)079)에 의하여 연구되었으며 이에 감사 드럽니다
.문 헌
1) S h im izu . T. a n d Wolfe, L. S.
:
Arachidonic acid cascade a n d s ig n al tra n sd u c tio n . J. Neuroche - mistry. 55, 1 (1990).2) S m it h . W . L .
:
P ro s ta n o id b io sy n th e s is a n d m e chan ism s of action. Am. J. Physiol 263. F181 (1992).3) W illia m s . C . S. a n d D u B o is , R . N .
:
P ro s ta g la n d in endoperoxide synthase:
W h y tw o isoforms? Am. /. Physiology, 270. G393 (1996).
4) H e rs c h m a n . H . R . ■ P ro sta g la n d in synthase 2, Biochim. Biophys. Acta, 1299. 125 (1996).
5) S m ith . W . L.
:
P ro sta g la n d in biosynthesis a n d its c o m p a rtm e n ta tio n in vascular sm ooth m u s cle a n d endothelial cells Ann. Rev. Physiol. 48.251 (1986).
6) N eedlem an. P.. T urk. J ., Ja k sc h i. B. A.. M o rris
on. A. R . a n d L e fko w ith. J . B.
;
A rac h id o nic a c id m e ta b o lis m . Ann. Rev. Biochem. 55. 69(1986).
7) S im m on s, D . L.. Levy. D . B ., Y a n n o n i. Y . a n d E rikso n, R . L.
:
Identification o f a phorbol ester- r e s p o n s ib le v .s rc - in d u c ib le gene. Proc. Natl.Acad. Sci. USA 86. 1178 (1989).
8) Xie. W .. C h ip m a n . J . G .. Robertson. D . L., Erik- son. R. L. an d Sim m ons. D . L.
;
Expression of a m itogen responsive gene encoding prostaglandin synthase is regulated by m R N A splicing. Proc.Natl. Acad. Aci. USA 88. 2692 (1991).
9) F u . J . Y ., Masferrer. J .F .. Seibert. K , R az. A.
a n d N eedlem an, P.. The induction a n d suppres
sion o f p ro s ta g la n d in H2 sy n th a s e (cyclooxy- genase) in h u m a n monocytes J. Biol. Chem, 265, 16737 (1990).
10) Riese, J .. Hoff. T.. Nordhoff, A .. D eW itt. D . L., Resch. K. a n d K aever V.
;
T ransient expression o f pro stag lan din endoperoxide synthase-2 d u r in g m ouse m acrophage activatio n. J. Leukocyte Biol. 55. 476 (1994).11) Siro is, J .
:
In d u c tio n o f p r o s ta g la n d in e n d o peroxide synthase-2 by h u m a n chorionic g on adotropin in bovine preovulatory follicles in vivo.
Endocrinology 135. 841 (1994).
12) R ester, M .. Coroneos. E .. T h o m a s P . J . a n d D u n n . M . J . '■ E n d o t h e lin s t i m u l a t e s p r o s ta g la n d in endoperoxide synth ase ^2 m R N A e x pression a n d protein synthesis th r o u g h a ty ro sine k in a s e s ig n alin g p a th w a y in r a t m e s a n g ia l cell. /. Biol Chem. 269. 22574 (1994).
13) B a za n , N . G .. Fletcher. B. S.. H e r s c h m a n . H . R . a n d M u k h e ije e , P. K . P la te le t a c tiv a tin g fac-.
to r a n d retinoic acid synergistically a c tiv a te th e in d u c ib le p r o s ta g la n d in s y n th a s e g e n e . Proc.
Natl. Acad. Sci. USA 91 5252 (1994).
14) Lee. S. H .. Soyoola, E .. C h a n m u g a m . P ., H a r t.
S., S u n , W .. Zhong, H .. L ion, S.
:
S im m o n s . D . a n d H w a n g . D .. S ele ctive e x p re s s io n o f m i- togen-inducible cyclooxygenase in m a c ro p h a g e s s tim u la te d w ith lipopolysaccharide J. Bioi Chem.267. 25934 (1992).
15) Jones. D . A .. C arlton. D . P., M c In ty re . T. M ..
Z im m e rm a n , G . A. a n d Prescott, S. M .
:
M olecu la r c lon in g o f h u m a n p r o s ta g la n d in e n d o p e roxide synthase type II a n d d em on stratio n o f ex
pression in response to cytokines. ]. Biol Chem.
268, 9049 (1993).
16) K u ju b u , D . A . a n d H e rs c h m a n . H . R .. D e x a m e th a s o n e in h ib its m ito g e n in d u c tio n o f th e T IS IO p r o s ta g la n d in sy n th as e /c y clo o x y g e n a se gene. J. Biol Chem. 267. 7991 (1992).
17) H a m a s a k i, Y .. K itzler. J .. H a r d m a n . R .. N et- te she im . P. a n d E lin g , T. E. '■ P h o rb o l e ster a n d e p id e r m a l g r o w th fa c to r e n h a n c e t h e e x pression o f tw o in d ucible p ro s ta g la n d in H s y n th a se genes in r a t trache al e p ith e lia l cells. Arch.
Biochem. Biophys. 304. 226 (1993).
18) O 'S u lliv a n . M . G .. C h ilto n . F. H .. H u g g in s . E . M . a n d M c C a ll. C . E . • L ip o p o ly s a c c h a r id e p r im in g o f alveolar m acrophages for e n h a n c e d syn th esis o f prostanoids involves in d u c tio n o f a novel pro stag la n d in H synthase. J. Biol. Chem.
267. 14547 (1992).
19) D uBois, R . N .. Aw ad, J .. Morrow. J .. Roberts. L.
J . a n d B ishop. P. R .
:
R e g u latio n o f eicosanoid production a n d mitogenesis in ra t in te s tin a l epithelial cells by transform ing grow th factor-a a n d
Vol 4h No. 6, 1997
이수환• 우 현 구 • 김지영• 백은주• 문창현
phorbol ester. /. Clin. Invest. 93. 493 (1994).
20) D e W itt. D . L. a n d M eade. E . A.
:
S e ru m a n d glucocorticoid re g u la tio n o f gene tra n sc rip tio n a n d expression o f p ro stag la n din H synthase-1 a n d pro stag lan din H synthase-2 isozym es. Arch.Biochem. Biophys. 306. 94 (1993).
21) Coyne, D . W .. N ickols, M .. B ertrand. W . a n d M orrison. A . R .
;
R e g u la tio n o f m e s a n g ia l cell cyclooxygenase synthesis b y cytokines a n d g lu cocorticoids. Am. J. Physiol. 263, F97 (1992).22) Lee, S. H .. H a rt. S.. Q ia o D . C h a n m u g a m P..
S o y o o la E .. L io u S .. K a n e m a r u , Y .
:
a n d H w a n g . D . Expression o f cyclooxygenase (C O X ) in s m o o th m u sc le cells (S M C s ). FASEB /. 7 s u p p l 1. A661 (1993).23) F e n g. L .. S u n . W .. X ia . Y .. T ang, W . W .. C h a n m u g a m , P .. S o yo o la. E ., W ils o n E . B . a n d H w a n g . D . C lo nin g o f tw o isoforms o f r a t cy
clooxygenase- differential reg ulation o f th e ir ex
pression. Arch. Biochem. Biophys. 307. 361 (1993).
24) Lee. S. H . E ffects o f b a c te r ia l lip o p o ly s a c charid e on p ro stag lan din production in prim ary c u ltu re d ra t v ascular sm ooth m uscle cells. Kor. J.
Food Hyg. Safety 11. 227 (1996).
25) L a n g . C . H .. D obrescu. C .. B a g y G . J . a n d S p itz e r J . J .
:
A lte re d glucose k in e tic s in d iabetic rats d u rin g g ra m negative infection. Am.
J. Physiol. 253. E123 (1987).
26) R a e tz , C . R . H . B io c h e m is try o f endotoxins.
Ann. Rev. Biochem. 59. 129 (1990).
27) M orrison. D . C. a n d R y a n , J . L.
:
E n dotox ins a n d disease m echanism s. Ann. Rev. Med. 38. 417 (1987).28) Lee. T. A .. S a lts m a n K . A ., O h a s h i H . a n d K in g G . L. '■ A ctivation o f protein kinase C by e levation o f glucose concentration
:
proposal for a m e c h a n is m in th e d e v e lo p m e n t o f d iabetic v a s c u la r c o m p lic a tio n s Proc. Natl. Acad. Sci.USA 86. 5141 (1989).
29) C raven. P. A. a n d D eR ub ertis, F. R .
:
Protein kin a se C is activated in glom eruli from strepto zo to c in d ia b etic rats-possible m e d ia tio n by
glucose. /. Clin. Invest. 83, 1667 (1989).
30) Thyberg, J .. H edin, U .. Sjolund, M .. Palm beg. L.
a n d Bottger, B. A .
:
R egulation of differential properties a n d proliferation of arterial sm ooth m u s cle cells. Arteriosclerosis 9, 413 (1990).
31) W illia m s , B . a n d S c h rie r, R . W . C h a r a c t e rizatio n o f glucose-induced in s itu protein k i
n a se C a c tiv ity in c u ltu re d v a s c u la r sm o oth m uscle cells Diabetes 41, 1464 (1992).
32) W illia m s o n , J . R ., C h a n g . K . Frangos. M .. H a sa n, K .. Ido, Y .. K a w a m u ra . T.. N yengaard. J . R .. E n d e n . M . V. D .. Kilo, C. a n d Tilton. R . G .
:
H y p e rg ly c e m ic p s e u d o h y p o x ia a n d d ia b e tic com plications Diabetes 42. 801 (1993).
33) O k u d a , Y . , K a w a s h im a , K .. S u z u k i , S ..
A s a k u ra , Y .. A sano. M .. T s u ru m a ru , K ., D ai.
H .. T a c h i. Y .. B a n n a i. C .. S a ito h , M . a n d Y a m a s h ita K.
:
R estoration o f nitric oxide prod uc tio n b y aldose reductase in h ib ito r in h u m a n e ndo the lial cells cultu re d in h ig h glucose m e d i
u m Life Sci. 60. 53 (1997).
34) Z ahng. H ., Kaseki, H ., Davis. W . B.. W hisler, R.
L . a n d C ornw ell. D . G .
:
M e c h a n is m s for th e s t im u la t io n o f p r o s ta n o id s y n th e s is b y c y closporine an d bacterial lipopolysaccharide Transplantation 47, 864 (1989).
35) N a k a n o . T.. O h a ra . O .. Teraoka. H . a n d A rita, H . G ro up I I phospholipase A 2 m R N A synthesis is s tim u la te d b y tw o d is tin c t m e c h a n is m s in ra t vascular sm ooth m uscle cells FEBS-Lett. 261.
171 (1990).
36) Flynn. J . T. a n d Hoff. H . Lipopolysaccharide in duces time-dependent increases in prostaglandin H synthas6~2 a n d cytosolic ph osp ho lipase A2
m R N A in cultu re d h u m a n microvessel-derived endothelial cells Shock 4. 433 (1995).
37) P ritchard. K . (T B a n io n M . K., M ia no . J . M ..
Vlasic. N .. B h a tia . U . G .. Y o un g . D . a n d Stem - e rm a n . M . B. In d u c tio n of cyclooxygenase-2 in r a t vascular sm o oth m uscle cells in vitro a n d in vivo J. Biol. Chem. 269, 8504 (1994).
/.