Effect of Ionizing Radiation on Rat Vascular Smooth Muscle Cell
12
0
0
전체 글
(2) 는 각각 87-2-11% vs 77-2-21%이었으며, 72시간째는 각각86-2-12% vs 71-2-27%로, 방사선 조 사군에서 24시간째에 G1기와 G2기의 arrest가 관찰되었고, 48시간째에 G2 arrest의 partial release가 관찰 되었다. 결 론:電離 放射線은 照射線量에 비례하여 혈관평활근세포의 증식을 억제시키는데, 이는 세포주기 중 G1/S, G2/M 期의 진행을 억제함에 의한 것이다. (Korean Circulation J 1998;28( (8) ):1322-1333) ) 중심 단어: 단어 혈관 평활근세포・전리 방사선・세포주기.. Introduction. system;Hannan et al used 7-9 Gy of γ-ray in 1994,15) and Fischell et al studied with the wire coated. Ionizing radiation inhibits cellular proliferation and. with 32P(0.00-20.006 μCi/cm) in 1994.16) There are many studies dealing with the mechanism. has been used extensively to treat proliferative neoplastic and nonneoplastic diseases. There are some. of cell death in various mammalian cells and cancer cells, but limited studies in VSMCs. In mammalian. precedent studies about radiation effects on non-cancer cells.1-3) Recently, many investigators have reported that. cells, ionizing radiation causes breaks in the DNA. Cells with damaged DNA stop DNA replication by. local irradiation could inhibit the SMC proliferation which is the cause of restenosis. Radiation therapy to. arresting in G1, or if the cells is already passed the restriction point in G1, by transiently arresting in S phase. inhibit SMC proliferation after arterial injury represents a reasonable and relatively new approach to. with both inhibition of replicon initiation and inhibition of replicon elongation. In addition, cells with damaged. inhibit restenosis after PTCA. A portion of the SMC in the vessel wall rapidly. DNA fail to progress into mitosis by arresting in G2.17) Cancer cells have mutation in p53 and lose the G1. entered the cell cycle during the first 48 hour following injury,4) and these cells were likely to be the progenitors. checkpoint. But they may be particularly sensitive to inhibition of the G2 checkpoint.18) In the study with. of continued proliferation, contributing to neointimal hyperplasia. Because radiation affects tissues primarily. fibroblast, ionizing radiation is reported to induce the G1 arrest via the expression of p53/p21 gene.17)19) In. by killing proliferating cells, these proliferating SMC may be particularly susceptible to the effects of ionizing. the study with VSMCs they reported that irradiation inhibits VSMCs proliferation,6)16) but it is not clear as. radiation after arterial injury. In previous experimental studies, several investigators have shown that irradiation. to mechanism in which radiation inhibits the VSMCs proliferation is cell cycle arrest or cell death. It is very. effectively inhibits neointimal hyperplasia.5-11) In recent years, several clinical trials using local, catheter-based. important to study the mechanism by which radiation inhibits VSMCs proliferation to establish radiation. ionizing radiation have demonstrated that radiation therapy could inhibit neointimal hyperplasia signific-. therapy as a new modality to prevent restenosis. This study was designed to investigate the effects of. antly.12)13) However, the optimal dose of irradiation to prevent. γ-radiation on the proliferation and cell cycle of VSMCs which are the main components of intimal. proliferation of VSMCs has not been studied in detail. If the dose is too high, aneurysmal dilatation may. hyperplasia in restenosis after PTCA.. occur in weak arterial wall, while if the dose is too low, VSMCs proliferation will not be inhibited effectively.. Materials and Methods. It was reported that the dose of 10-14 Gy of βparticle could inhibit SMC proliferation in animal. Cell culture. study.7)14) There are a few studies which evaluated effects of various radiation doses using in-vitro culture. Primary culture of SMC Rat aortic SMCs were isolated by collagenase dige1323.
(3) stion of thoracic aorta of 6-week-old Sprague-Dawley rat, and cultured in Dulbecco’s modified Eagle’s medium. with the dose of 8 or 16 Gy were examined on a light microscope after Papanicolaou (Pap) stain using. (DMEM : Gibco(supplemented with 10% fetal calf serum (Sigma) and 20 mmol/L HEPES, in 5% CO2/ air. Variation 24-3 automatic staining machine. Briefly, the staining method was as follows:The cells were. at 37℃. The medium was changed 2 times a week. The cells were passaged by trypsinization with 0.05% trypsin. hydrated in 50% of ethyl alcohol for 10 seconds and in running water for 1 minute. The nuclei were stained. EDTA and used between 5th and 7th passage.. with Hematoxylin. Then the cells were dehydrated in various concentrations of ethyl alcohol (50, 80, and. Identification of SMC. 95% ethyl alcohol for 20 seconds respectively). The cytoplasm was stained with orange G6 and eosin-. The SMCs were verified by observing“hill-andvalley” , characteristic growth pattern of SMCs with a. alcohol (EA)-36 for 3 minutes, each. The cells were dehydrated in 95% and 100% ethyl alcohol and. microscope. And the cultured cells were fixed in 4% paraformaldehyde and stained immunocytochemically. cleaned with xylene 3 times.. with monoclonal anti-α-smooth muscle actin antibody (DAKO)).. Analysis of cell proliferation Proliferation of cells were assessed by counting viable cells and measuring incorporation of [3H] thymidine into DNA. Analysis of radiation effect Dose-response analysis. Viable cell counting 137. The cultured cells were irradiated using a Cs source (irradiator:Shephard Co., Cesium-137:MK. Cells were cultured in 24-well culture plate. Initial seeding cell number was 0.25×105 cells/cm2. When. 01-68 8000 Ci, 1983) with the dose rate of 4 Gy/min. When the cells formed confluence of 50-60%, they. the cells had 60%-confluent, they were irradiated with the dose of 2, 8, or 16 Gy. After trypsinization. were exposed to the dose of 2, 8, or 16 Gy by changing exposure time. The effects of radiation were analyzed. with 0.05% trypsin-EDTA the cells were stained with trypan-blue. Suitable aliquots of the cell suspension. as described next at 12, 24, or 48 hours after irradiation.. were applied on hemocytometer and the number of cells which could exclude trypan-blue were counted on. Irradiation at various culture conditions. a light microscope.. The cells were cultured in aminoacid-rich medium (DMEM/F12, 315 mg/dl of glucose). They were exposed to radiation with a certain fixed dose, and. Incorporation of [3H] thymidine into DNA. harvested at 24 or 48 hours after irradiation. The same procedures were repeated at different culture condi-. thymidine incorporation. Cells were cultured as previously described. When the cells had 60%-confluent,. ions as follows; 1) cell culture in high glucose medium (DMEM. they were irradiated with the dose of 8 Gy, and subsequently incubated with [3H] thymidine (464 mCi/. with 400 mg/dl of glucose) 2) cell culture in low glucose medium (DMEM. mmol, New England Nuclear) in the presence of 1 μ Ci/ml thymidine. Following incubation for 24 hours in. with 100 mg/dl of glucose) 3) cell culture in the medium containing estrogen. DMEM, the reaction was terminated. The cells were rinsed with solution (10% Tris, 150 nM NaCl, pH 7.4). (DMEM with 180 nM of 17β-estradiol (Sigma)). at 40℃, and fixed with 10% trichloroacetic acid. Finelly they were rinsed with 100% ethanol and dried.. Microscopic observation. The incorporated radioactivity was measured by using a liquid scintillation spectrometer.. Morphological changes of the cells after irradiation 1324. DNA synthesis of VSMCs was measured by [3H]. Korean Circulation J 1998;28(8):1322-1333.
(4) Analysis of cell cycle. Results. The cells cycles were analyzed using Fluorescence Activated Cell Sorter (FACS).. Cell culture PI-FACS The cells were trypsinized to single cell suspension.. With microscopic observation, the cells grew to multilayered hill-and-valley pattern, the characteristic. After fixation with 70% ethanol at 4℃ for 30 minutes, the cells were centrifuged, then rinsed with phosphate-. property of SMCs (Fig. 1). Fig. 2 showed the cultured cells which were stained. buffered saline(PBS). They were incubated in 1mg/ml RNase A (Boeringer Mannheim) for 30 seconds at. with monoclonal anti-α-smooth muscle actin antibody. The cytoplasm of all presenting cells are stained in. 37℃. Immediately before flow cytometric analysis, the. brown color which means that all presenting cells are VSMCs containing α-smooth muscle actin. The α-. cells were incubated in 50 μg/ml propidium iodide (PI:Sigma) solution for 30 minutes at room tem-. smooth muscle actin is labeled with mouse monoclonal anti α-smooth muscle actin antibody. Secondary lab-. perature. The samples were analyzed using FACS flow cytometer (Becton-Dickinson)with the rate of 10,000. elling is achieved with horseradish peroxidase-conjugated rabbit anti-mouse antibody.Effect of radiation on. cells/second. Twenty thousand cells were scanned per each sample. Cell cycle was analyzed by the propo-. SMC proliferation The effect of γ-irradiation on the growth of rat. rtion of cells at G0/G1, S and G2/M phase.. VSMCs in culture was studied. Just before irradiation, the cell count was 0.4×105. BrdU+ +PI-FACS One ml of 5-bromo-2’-deoxyuridine (BrdU:. cells/cm2. After irradiation with the dose of 0, 2, 8, or 16 Gy (n=9 at each dose), the cell counts at 24 hour. Boe-ringer Mannheim) solution was added to culture medium and incubated for 10 minutes. The cells were. were 3.3±0.9, 2.3±0.6, 1.9±0.6, 1.3±0.2×105 cells /cm2, and those at 48 hour were 5.1±2.1, 2.0±0.8, 1.8. fixed with 95% ethanol. After treatment with formaldehyde, cells were heated in a 98℃ water bath for. ±0.6, 1.2±0.5×105 cells/cm2, respectively. The cells of control group grew by 56% at 48 hour compared. 10 minutes for partial DNA denaturation and then cooled immediately in an iced water bath. The cells. with those at 24 hour. However, the cell growth of irradiated groups was inhibited by radiation and this. were suspended in 1.0 ml of mouse monoclonal antiBrdU antibody (1:25 dilution, DAKO) and incu-. phenomenon was dose-dependent (Fig. 3).. bated for 1 hour at 37℃. After then they were rinsed and incubated with 1.0ml of fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse F anti-body (1:20 dilution, DAKO) at 37℃ for 75 minutes. Immediately before flow cytometric analysis the cells were resuspended in PI solution.. Data analysis Data were reported as mean±SEM. All results were analyzed by ANOVA test. Dose-response effect was analyzed by linear regression. The value within 95% confidence interval (p<0.05) was considered significant.. Fig. 1. Photomicrograph of cultured rat VSMCs showing typical hill-and-valley appearance (original magnification ×200).. 1325.
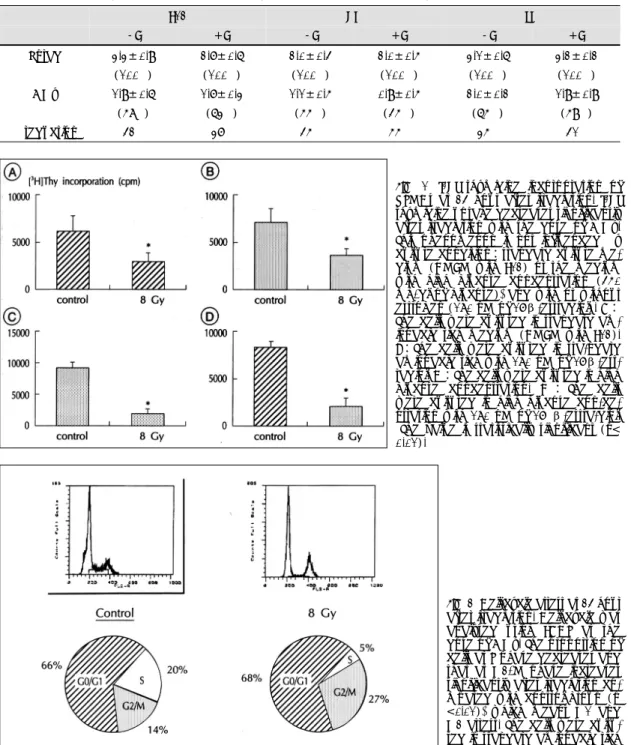
(5) Radiation effect in various culture conditions The cells were counted at 24 hour after radiation. this phenomenon was not influenced by culture conditions, such as with high glucose concentration or. with the dose of 0, 2, 8, or 16 Gy in various culture conditions. The inhibition of cell growth was consi-. with/without estrogen (Fig. 6).. stently observed regardless of culture conditions;high (450 mg/dl) or low (100 mg/dl) glucose concentration. Effect of radiation on SMC cycle. or with/without estrogen (180 nM of β-estradiol) (Figs. 4 and 5, Table 1).Effect of radiation on DNA. irradiation at a dose of 8 Gy. The proportion of cells at G0/G1-S-G2/M phase was 66-20-14% in control. synthesis of SMC DNA synthesis of VSMCs was analyzed at 24 hour. group, while 68-5-27% in irradiated group. The proportion of cells at S phase decreased and that at. after irradiation at a dose of 8 Gy. The [3H] thymidine uptake significantly decreased after irradiation. And. G2/M phase increased after irradiation (Fig. 7). Figure. Fig. 2. Photomicrograph of immunocytochemical stain for α-smooth muscle actin. Diamidinobenzidine substrate is used for color development which results in brown color (original magnification ×400).. Fig. 3. Sequential changes of VSMCs counts after irradiation. After irradiation with the dose of 2, 8, or 16 Gy, the cell growth is inhibited. The cell counts at 48 hour increases significantly compared with those at 24 hour (p<0.05) in control group, but the cell counts do not increase at 48 hour compared with those at 24 hour in irradiated group. The cells were cultured in aminoacid rich medium (DMEM with F12:standard culture condition).. 1326. The cell cycle was analyzed at 24 hour after. Fig. 4. Effect of estrogen on change of VSMCs count at 24 hour after irradiation. The cells were cultured in standard aminoacid rich medium (DMEM with F12) with/without estrogen (180 nM of 17β-estradiol). The cell counts after irradiation were significantly lower than those of unirradiated group at all radiation doses. The dose-dependent inhibitory effect on cell proliferation of radiation persists in standard aminoacid rich medium.. Fig. 5. Effect of high glucose concentration (450 mg/ dl) with/without estrogen (180 nM of 17β-estradiol) on change of VSMCs count at 24 hour after irradiation. The dose-dependent inhibitory effect on cell proliferation of radiation persists even in high glucose medium.. Korean Circulation J 1998;28(8):1322-1333.
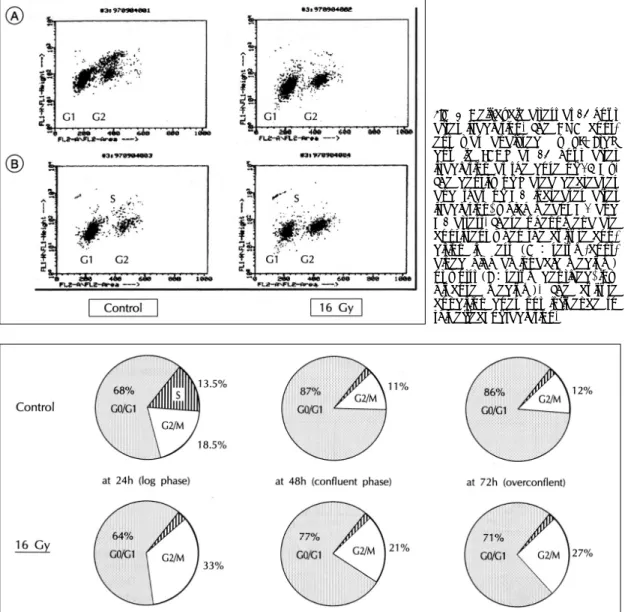
(6) 8 showed the DNA content analyzed by PI+BrdU double FACS at 24 hour after irradiation at a dose of. contained DMEM/F12)or worst (serum free, low glucose medium). These effects of radiation on cell. 16 Gy. The density of S area decreased and that of G2 increased after irradiation. This phenomenon was con-. cycle were similar in any culture condition. The change of cell cycle was observed serially after. sistent when the culture condition was best (serum-. irrradiation at a dose of 16 Gy.The proportion of cells. Table 1. Changes of VSMC counts at 24 hour after irradiation with the dose of 8 Gy F12. HG. LG. -E. +E. -E. +E. -E. +E. Unrad. 3.3±0.9. 2.7±0.6. 2.0±0.4. 2.0±0.5. 3.1±0.6. 3.2±0.2. (100%). (100%). (100%). (100%). (100%). (100%). 8 Gy. 1.9±0.6. 1.7±0.3. 1.1±0.5. 0.9±0.5. 2.0±0.2. 1.9±0.9. (58%). (63%). (55%). (45%). (65%). (59%). 42%. 37%. 45%. 55%. 35%. 41%. % reduction. Fig. 6. [3H] thymidine incorporation of VSMCs at 24 hour after irradiation. [3H] thymidine uptake decreases significantly after irradiation with the dose of 8 Gy. This phenomenon is not influenced by culture condition:standard culture medium (DMEM with F12) or the medium with high glucose concentration (450 mg/dl of glucose), and with or without estrogen (180 nM of 17β-estradiol). A: The cells were cultured in standard aminoacid rich medium (DMEM with F12). B:The cells were cultured in sta-ndard aminoacid rich with 180 nM of 17β-estradiol. C:The cells were cultured in high glucose concentration. D : The cells were cultured in high glucose con-centration with 180 nM of 17 β-estra-diol. *The value is statistically significant (p< 0.01).. Fig. 7. Cell-cycle arrest at 24 hour after irradiation. Cell cycle was analysed using FACS at the dose of 8 Gy. The proportion of cells at S phase decreases and that at G2/M phase increases significantly after irradiation compared with control group (p <0.01), which means G1 and G2 arrest. The cells were cultured in standard aminoacid rich medium.. 1327.
(7) in G0/G1-S-G2/M phase at 24, 48, 72 hour was 643-33, 77-2-21, 71-2-27% in irradiated group, and. which may be consequence of the partial release of G2 arrest after 24 to 48 hour of irradiation.. 68-14-18, 87-2-11, 86-2-12% in control group, respectively (Fig. 9). The proportion of cells at S. Effects on cell size and nuclear DNA amount. phase decreased and that at G2/M phase increased at 24 hour after irradiation, which means G1 and G2. The cell size and amounts of DNA increased after irradiation (Fig. 10). These changes were similar rega-. arrest. The proportion of cells in G2/M phase in irradiated cells decreased from 24 to 48 hour (33% vs. rdless of dose and time (8 or 16 Gy, 24 or 48 hours). According to microscopic observation, large, polygonal. 21%)but still higher than that of unirradiated cells,. cells and many binucleated cells with abundant cytop-. Fig. 8. Cell-cycle arrest at 24 hour after irradiation. The DNA content was analysed by PI+BrdU double FACS at 24 hour after irradiation at the dose of 16 Gy. The density of S area decreases and that of G2 increases after irradiation, which means G1 and G2 arrest. These phenomena are consistent when the culture condition is best (A : serum-contained high aminoacid medium) or worst (B:serum deprived, low glucose medium). The culture condition does not influence to this effect of radiation.. Fig. 9. Sequential patterns of cell-cycle of VSMCs after irradiation. The change of cell cycle was observed serially after irradiation at the dose of 16 Gy. The proportion of cells at S decreases and that at G2/M increases at 24 hour after irradiation, which means G1 and G2 arrest. The proportion of cells in G2/M phase in irradiated cells decreases from 24 to 48 hour (33 vs 21%) but still higher than that of unirradiated cells, which means the partial release of G2 arrest from 24 to 48 hour after irradiation. The cells were cultured in standard aminoacid rich medium.. 1328. Korean Circulation J 1998;28(8):1322-1333.
(8) Fig. 10. Change of cell size and nuclear DNA amount of VSMCs. The cell size and DNA amounts analyzed by FACS increased at 24 hour after irradiation with the dose of 8 Gy, which suggested that irradiation inhibited cell division and produced multinucleated cells. The cells were cultured in standard high aminoacid medium.. i. e., G1 and G2/M phase arrest.. Cultured VSMCs as a model of restenosis Cultured VSMCs have different characteristics from those in normal physiologic condition. And under the same physiologic condition, SMCs that migrated into intima have a different phenotype and different gene expression from those in normal medial wall of artery. It is believed that the cultured SMCs have similar physiologic characteristics to those in atherosclerotic Fig. 11. Photomicrograph of rat VSMCs 24 hours after irradiation with the dose of 8 Gy. Large polygonal and binucleated cell with abundant cytoplasm are observed after irradiation, which suggests that radiation inhibits cytokinesis, cytoplasmic division (original magnification ×400).. lasm appeared after irradiation (Fig. 11). These findings suggests that radiation inhibits cytokinesis, cytoplasmic division, and produces multinucleated cells.. Discussion In this study, the effects of γ-radiation on the proliferation and cell cycle of VSMCs were investigated using 137Cs in culture system. The proliferation of VSMCs was inhibited by γ-radiation in a dose related fashion. The mechanism by which irradiation inhibited VSMCs proliferation was via cell cycle arrest,. lesion, and Ross reported that in vitro cultured VSMCs have similar biologic characteristics to those of neointima following balloon catheter and other injury to the arterial wall, as they have passed several passage.20) Therefore using cultured VSMCs as a model of restenosis after PTCA is reasonable.. Radiation source β-and γ-ray are widely used in radiation therapy. β-particle irradiation with 32P offers the advantages of a short half-life (14.3 days) and limited range in tissue (3-4 mm). This would eliminate the risk of catheterization laboratory personnel and minimize the exposure of surrounding cardiac and pulmonary tissue to ionizingradiation. However, because of limited range in tissue, centering should be required when endovascularly applied. If not, the surface dose varies significantly resulting in eccentric effects and the 1329.
(9) proliferative VSMCs remained. γ-Ray needs long exposure time (30-60 minutes) to get therapeutic. and reservoir of SMCs that can proliferate. Brenner at al reported that a dose of around 25 Gy would be. effects, hense exposed total body dose is relatively high. This has some risk to surrounding cardiac and. needed to eliminate completely an assumed target population of 107 potentially clonogenic SMCs.22) This. pulmonary tissue, and operating personnel. But γ-ray has a even dose distribution and the centering is not. is a larger dose than can be realistically used, as it would be likely to produce an unacceptable risk of late. necessary. The histology and morphologic analysis of swine coronary arteries treated with the same doses of. sequelae. Waksman reported that a dose response relation was demonstrated, but further inhibitory effect. β-and γ-radiation demonstrated an overall similarity.21) Thus the biological effect of the radiation on. was absent beyond 28 Gy.8) In general, it is accepted that under the 20 Gy, radiation inhibits VSMCs more. neointimal formation depends on the absorbed dose and not on the specific type of isotope or the variation. potently as the dose is increased, and lower dose may be either stimulatory or less effective. Therefore it is. in treatment times seen with the different approaches.. reasonable to assume that the optimal dose of radiation applied to inhibit restenosis would be 15-20 Gy.. Effects on cell proliferation Radiation has a beneficial effect of inhibiting SMC. Mechanism of cell damage. proliferation. This study established that radiation inhibits VSMCs in a dose-dependent manner. This result. When the cell nucleus is exposed to ionizing radiation, DNA mutations and chromosomal aberrations can. is consistent with many other clinical data. Only one experimental study failed to show a beneficial effect of. occur. As a result, the ability of the cells to replicate is impaired, and ultimately cell death takes place. Damage. radiation on the restenotic process.5) Schwartz et al found that X-irradiation actually modestly increased the. to the DNA by radiation has been intensively studied. When nuclear DNA has been damaged, normal cells. neointimal proliferation seen after oversized stent placement in porcine coronary arteries. 5) This negative result. cease to progress through the cell cycle at one of several points, i. e., G1 and G2 arrest. Recently, there was a. was possibly due to methodological differences, which include the use of relatively low radiation doses (4 or 8. report of the effect of irradiation on cell cycle of fibroblast that has similar characteristics to VSMCs.19). Gy at day 1, and 4 Gy at day 1 and 8 Gy at day 4) in the setting of more extensive vessel injury (stent:. Nasrin et al reported that G1 arrest occurred in fibroblast after irradiation with the dose of 4 Gy.19) In this. artery oversize ratio was 1.5-2.0:1). As for the lowest limit of effective dose, Abbas et al. study, the proportion of cells at S decreased and that of G0/G1 and G2/M were increased after irradiation.. reported that there was no inhibitory effect with the dose of 6 Gy in his rabbit iliac model,6) and Waksman. Increasing the proportion of G0/G1 and decreasing the proportion of S phase indicate that the cells in G1. reported that SMCs proliferation would not be inhibited with 3.5 Gy.7) In the present study, there was a. phase cannot be passed to S phase, or G1 arrest, while increasing the proportion of G2/M phase means that. inhibiting effect on VSMCs proliferation at least with the dose of 2 Gy. This finding is consistent with other. the cells in G2/M phase cannot be passed to G1 phase, or G2 arrest. Radiation induces G1 and G2 arrest in. prior studies that have demonstrated that proliferating VSMCs in vitro are sensitive to relatively low doses. VSMCs. The increased proportion of cells in G2/M phase decreased as time elapsed (33% vs 21% in this. (2-5 Gy) of external beam (X-ray) irradiation.6)16) This difference between in vivo and in vitro experi-. study), which indicates partial release of G2 arrest. Yanagihara also reported similar results of the escape. ment may be due to differences in study systems. In animal study (in vivo study) there are many factors. from G2 arrest.23) This means the recovery of SMC proportion that can proliferate again. It is probable,. that could influence the radiation effects on SMCs proliferation, such as growth factors, various cytokines,. then, that the radiation doses applied both clinically and experimentally, do not entirely eliminate the. 1330. Korean Circulation J 1998;28(8):1322-1333.
(10) potential clonogenic SMC population. Consequently, it is likely that the remaining SMCs will grow back and,. inhibition when radiation was delivered at the time of injury, marked inhibition when radiation was applied. unless they reach the end of their proliferative capacity, they will eventually causes restenosis. Wiedermann. at 2days after injury;and no inhibition when the radiation was applied at 7days after injury.25) These. reported the inhibitory effects of radiation on SMC proliferation lasting 6 months in his porcine model.10). data suggest that radiation given near the peak of mitotic activity more effectively suppresses subsequent. The reduction of neointimal area seen in his irradiated animals, when compared with that in the control group. neointimal formation. In clinical conditions, radiation can be applied immediately before or after PTCA. This. at 6 months, was nearly identical to the differences in neointimal areas between groups at 30days.24) This. may reduce the effect of radiation and larger dose may be required to get an inhibiting effect than in culture. strongly suggests that proliferative process responsible for restenosis is essentially complete at 30days, with. system. With γ-radiation, the duration of radiation therapy is approximately 30-60 minutes to get a the-. little change over subsequent months. This difference of the results between in vitro and animal study can be. rapeutic dose. Coronary blood flow is disturbed during the therapy, and this may cause thrombus form-. further explained as follows;Even though SMC is not completely eliminated, irradiation may reduce the cells. ation which results in sudden arterial occlusion. Reendothelialization after PTCA or stent implan-. available for proliferation and migration into the neointimal zone in vivo and the bands of fibrosis after. tation may be affected by radiation. A delay in reendothelialization could, theoretically, adversely affect. irradiation may also act as a diffusion barrier for mediators of chemotaxsis and cellular proliferation as. the risk of subsequent stent thrombosis. Vscular lesions after irradiation may occur. And There are no. well as physical impediments to cell migration.. data about the long term side effects of irradiation. Wiedermann et al reported that there was no evidence. Conditions that can change radiosensitivity One preliminary study reported that the restenosis. of radiationinduced damage to the surrounding myocardium, no neoplastic transformation and no evid-. rate is low in women and the patients with DM who had taken radiation therapy (SCRIPPS trial:Teirstein PS. ence of radiation vasculopathy in his swine model at 6 month follow-up.10) Despite these encouraging data,. 1997). It is conceivable that the patients with DM or female patients are more sensitive to radiation. In this. the potential from injury from intracoronary irradiation on the vasculature and myocardium remains to. study, however, the degree of radiation-induced inhibition of cell growth or DNA synthesis was not signifi-. be established. The γ-radiation potently inhibits the proliferation. cantly different among various culture conditions. Of course, neither the effect of high glucose concen-. of VSMCs in a dose dependent manner by the mechanism of cell cycle arrest (G1 and G2/M arrest). These. tration in culture media can be extraplated to the clinical situation of DM nor that of estrogen in media. findings were consistently observed in the high glucose as well as the low glucose status with or without estr-. can be extrapolated to the female’s response to irradiation. Further studies should be done to investigate. ogen. These data suggest that radiation therapy adjunctive. whether the DM or female gender can modify the biologic response of vascular smooth muscle cells to. to PTCA would be effective in inhibiting neointimal hyperplasia.. irradiation.. Summary Considerations in clinical application Sheffer et al demonstrated that the inhibition of SMC proliferation in their model was critically dependent on the timing of the radiation exposure:minimal. Background: : Recently many investigators have reported the studies of radiation therapy to prevent proliferation of 1331.
(11) vascular smooth muscle cells (VSMCs) which is the main cause of restenosis. This work is designed to investigate the effects of γ-radiation on the proliferation and cell cycle of VSMCs. Method: : Rat aortic smooth muscle cells were isolated, and the. dependent manner by the mechanism of cell cycle arrest. KEY WORDS:VSMCs・γ-irradiation・Cell cycle.. REFERENCES 1) Borok TL, Bray M, Sinclair I. Role of ionizing irradi-. cells were used between 5th and 7th passage. The cultured cells were irradiated using Cesium-137 (137Cs). 2). with the dose of 2, 8, or 16 Gray (Gy), and harvested at 12, 24, or 48 hours after irradiation. Proliferation of. 3). cells was subsequently assessed. And the changes of DNA synthesis and cell cycle were analyzed by flow cytometry. Results: : 1) The number of inoculated cells was 0.4×105 cells/cm2. After irradiation with the dose of 0, 2, 8, or 16 Gy, cell counts at 24 hour were 3.3±0.9, 2.3±0.3, 1.9±0.6, 1.3±0.2×105 cells/cm2, and those at 48 hour were 5.1±2.1, 2.0±0.8, 1.8±0.6, 1.2±0.5×105 cells/cm2, respectively. The cells of control group grew by 56% at 48 hour compared with those at 24 hour, however the cell growth of irradiated groups was inhi-. 4) 5). 6). 7). bited in a dose-dependent manner. 2) The inhibitory effect of irradiation was consist-. 8). ently observed when the culture condition was changed to high (450 mg/dl) or low glucose concentration (100. 9). mg/dl) and with or without estrogen (180 nM of 17 βestradiol). 3) DNA synthesis was analysed by [ 3H] thymidine incorporation. Radioactivity of cells decreased significantly after irradiation (8 Gy), and this phenomenon was not influenced by various culture conditions; high aminoacid medium (with F12, high glucose concentration, or with or without estrogen). 4) The cell cycle was analyzed after irradiation. The proportions of G0/G1-S-G2/M phase in control and irradiated group were 68-14-18 vs 64-3-33% at 24 hour, 87-2-11 vs 77-2-21% at 48 hour and 862-12% vs 71-2-27% at 72 hour after irradiation. G1 and G2 arrest occurred at 24 hour and partial release of G2 arrest were observed at 48 hour after irradiation. Conclusion: : The results of this study indicate that γ-irradiation potentially inhibits the VSMCs proliferation in a dose1332. 10). 11). 12). 13). 14). ation for 393 keloids. Int Radiat Oncol Boil Phys 1988; 15:865-70. Kovalic JJ, Perez CA. Radiation therapy following keloidectomy: A 20-year experience. Int Radiat Oncol Biol Phys 1989;17:77-80. Sylvester JE, Greenberg P, Selch MT. The use of postoperative irradiation for the prevention of heterotopic bone formation after total hip replacement. Int J Radiat Oncol Biol Phys 1988;14:471-6. London S, Mayberg MR. Kinetics of bromodeoxy uridine uptake by smooth muscle cells after arterial injury. J Vasc Res 1994;31:247-55. Schwartz RS, Koval TM, Edwards WD, Camrud AR, Bailey KR, Browne K, et al. Effect of external beam irradiation on neointimal hyperplasia after experimental coroanry artery injury. J Am Coll Cardiol 1992;19:1106-13. Abbas MA, Afshari NA, Stadius ML, Kernoff RS, Fischell TAl. External beam irradiation inhibits neointimal hyperplasia following balloon angiography. Int J Cardiol 1994;44:191-202. Waskman R, Robinson KA, Crocker IR, Wang C, Gravanis MB, Cipolla GD, et al. Intracoronary low-dose βirradiation inhibits neointimal formation after coronary artery balloon injury in the swine restenosis model. Circulation 1995;92:3025-31. Waksman R. Local catheter-based intracoronary radiation therapy for restenosis. Am J Cardiol 1996;78(suppl 3A): 23-8. Wiedermann JG, Marboe C, Amols H, Schwartz A, Weinberger J. Intracoroanry irradiation markedly reduces restenosis after balloon angioplasty in a porcine model. J Am Coll Cardiol 1994;23:1491-8. Wiedermann JG, Marboe C, Amols H, Schwartz A, Weinberger J. Intracoroanry irradiation markedly reduces restenosis after balloon angioplasty in swine: Persistent benefit at 6-month follow-up. J Am Coll Cardiol 1995; 25:1451-6. Verin V, Popowski Y, Urban P, Belenger J, Redard M, Costa M, et al. Intra-arterial irradiation prevents neointimal hyperplasia in a hypercholesterolemic rabbit restenosis model. Circulation 1995;92:2284-90. Teirstein PS, Massullo V, Jani S, Popma JJ, Mintz GS, Russo RJ, et al. Catheter-based radiation therapy to inhibit restenosis after coronary stenting, N Engl J Med 1997;336:1697-703. King SB 3rd, Williams DO, Chougle P, Klein JL, Waksman R, Hilstead R, et al. Endovascular beta-radiation to reduce restenosis after coronary balloon angioplasty: Results of the beta energy restenosis trial (BERT). Circulation 1998;26:2025-30. Carter AJ, Laird JR, Bailey LR, Hoopes TG, Farb A, Fischell DR, et al. Effect of endovascular radiation from a β-particle emitting stent in a porcine coroanry restenosis model. Circulation 1996;94:2364-8.. Korean Circulation J 1998;28(8):1322-1333.
(12) 15) Hannan MA, Kjougeer F, Halees Z, Sanei AM, Khan. 16). 17). 18) 19). BA. Increased radiosensitivity and radioresistant DNA synthesis in culture fibroblasts from patients with coronary atherosclerosis. Arterioscler Thromb 1994;14:1761-6. Fischell TA, Kharma BK, Fischell DR, Loges PG, Coffey CW, Duggan DM, et al. Low-dose, β-particle emission stent’wire results in complete, localized inhibition from ‘ of smooth muscle cell proliferation. Circulation 1994;90: 2956-63. Dulic V, Kaufmann WK, Wilson SJ, Tlsty TD Lees E, Harper JW, et al. p53-dependent inhibition of cyclindependent kinase activities in human fibroblasts during radiation-induced G1 arrest. Cell 1994;76:1013-23. Kastan MB, Onyekwere O, Sidransky D, Vogelstein B, Craig RW. Participation of p53 protein in the cellular response to DNA damage. Cancer Res 1991;51:6304-11. Nasrin N, Mimish LA, Manogaran PS, Kunhi M, Sigut D, AlSedairy S, Hannan MA. Cellular radiosensitivity, radioresistant DNA synthesis, and defect in radioinduction of p53 in fibroblasts from atherosclerosis patients. Aterioscler Thromb Vasc Biol 1997;17:947-53.. 20) Ross R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993;362:801-9.. 21) Gravanis MB. Morphologic findings after intracoronary. 22) 23) 24). 25). irradiation (beta & gamma) in the swine model of restenosis. Eds. Vascular brachtherapy. Veenendaal, Nucletron BV;1996. p.75-82. Brenner DJ, Miller RC, Hall EJ. The radiobiology of intravascular irradiation. Int J Radiat Biol Phys 1996; 36:805-10. Yanagihara K. Radiation induced apoptosis in cultured humen epithelial tumor cells. Nippon Rinsho 1996;54: 1943-8. Wiedermann JG, Marboe C, Amols H, Schwartz A, Weinberger J. Intracoroanry irradiation markedly reduces restenosis after balloon angioplasty in a porcine model. J Am Coll Cardiol 1994;23:1491-8. Sheffer A, Eigler NL, Whiting JS, Litvack FI. Suppression of intimal proliferation after balloon angioplasty with local beta irradiation in rabbits. J Am Coll Cardiol 1993;21(suppl A):185A.. 1333.
(13)
수치


+2

관련 문서
