154
Copyright © 2016 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
조피볼락
(Sebastes schlegeli)
은동북아연안,
일본홋가이도 이남및중국연안에분포하며,
수심100 m
내외의천해암초나 인공어초등에서식하기때문에자원증강이나레저용으로중요 한위치를차지하고있어경제적으로나사회적으로가치가큰 어종이다(Boehlert et al., 1986).
조피볼락은알을산란하는일 반어류와는달리체내에서수정하여새끼를산출하는태생어 류이다(Corey et al., 1983).
태생어류인볼락류에서산출된자 어들은인공사료에길들이기가어려운특징을갖고있어인공 종묘생산에있어서장시간에걸쳐서살아있는먹이생물을공 급해야한다는문제점을가지고있다(Lee et al., 1993a; Lee et al., 1993b).
현재인공종묘생산을위하여성립되어있는먹이생물의공급체계는 해수클로렐라
-
로티퍼(Rotifer)-
알테미아(Ar- temia)-
인공배합사료로의 공급체계가 구축되어 있다(Fujita, 1973; Kitajima, 1983; Hirayama, 1985; Yamasaki et al., 1987;
Yoshimura et al., 1996).
현재,
산업적인측면에서볼락류의인 공종묘생산에있어서가장큰문제점은불확실성이너무높다 는것이다.
이러한이유는여러가지가설이있지만,
가장유력 시되고있는가설은반세기걸쳐서그대로유지해온외줄타기 식의먹이생물공급체계에두고있다(Lee et al., 1993c; Lee et al., 2004).
현재구축된먹이생물중에서클로렐라와로티퍼를배양하여먹이로활용하는데는문제가없으나
, Artemia
는염호라는극한상황의환경에서자란갑각류의난을채집하여건 조통조림화한것으로서전량수입해서사용하고있다
(Kim et al., 1999).
또한Artemia
의지질영양학적인문제점에대해서는물벼룩(Moina macrocopa) 급여가 조피볼락(Sebastes schlegeli) 자어의 성장에 미치는 영향
정우철·Feng Jin·최종국·이정태
1·최병대·강석중
경상대학교 해양식품생명의학과, 1경상남도 수산자원연구소
Growth of Larval Rockfish Sebastes schlegeli Fed Moina macrocopa
U-Cheol Jeong, Feng Jin, Jong- Kuk Choi, Jeong-Tae Lee, Byeong-Dae Choi and Seok-Joong Kang
Department of Seafood and Aquaculture Science, Gyeongsang National University, Tongyeong 53064, Korea Fisheries Resources Resarch Institute, Gyeongnam, Tongyeong 50411, Korea
Several our studies have focused recently on the mass production of the freshwater Cladoceran Moina macrocopa which can substitute Artemia nauplii for the culture of larval marin fish. A 6 weeks experiment was conducted to investigate the effects of enrichment on the fatty acid composition of Moina macrocopa through feeding Schizo- chytrium sp. containing highly unsaturated fatty acids and to study the impacts of n-3-HUFA enriched Moina on improving survival rate and fatty acid compostion of larval rockfish Sebastes schlegeli . After feeding for 6 weeks, the Moina-fed fry resulted in a higher survival rate of 99.2% compared to the Artemia-fed fry 12.8%. In addition, the Moina-fed fry had the fast growth rate 45.6mm compare to the Artemia-fed fry 25.7 mm at the end of the experiment.
The Moina-fed fry showed significantly higher level of 16.47% DHA than their Artemia-fed fry counterparts of the level of 3.97% with respect to DHA. PL, the cell membrane components in living food organisms, constituted 63.8%
of the Moina, which was significantly higher than in the 40.1% of the Artemia. The present study indicate that Moina macrocopa can be used as Artemia substitute and improving the survival rate rockfish larvae through enchriment Schizochytrium sp.
Key words: Sebastes schlegeli, Moina macrocopa, Artemia, Rockfish, n-3 HUFA
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2016.0154 Korean J Fish Aquat Sci 49(2) 154-160, April 2016
Received 29 January 2016; Revised 18 March 2016; Accepted 11 April 2016
*Corresponding author: Tel: +82. 55. 772. 9154 Fax: +82. 55. 648. 3089 E-mail address: [email protected]
이미많은지적을하고있으며
(Choi et al., 1999), Artemia
는어 류의필수지방산으로알려져있는EPA, DHA
등의오메가-3
고 도불화지방산의함량이부족하기때문에이들성분을많이함 유한지질물질을보충하는 양양강화방법을사용하고있지만,
이를완전히해결하지못하고있는실정이다.
또한Artemia
의 양적수급문제,
가격폭등에대하여수십년에걸쳐서많은논 란이있음에도불구하고대체하지못하는이유는이를대신할 만한먹이생물을아직찾아내지못했기때문이다(Choi et al., 1999). Artemia
대체먹이생물로서물벼룩에대한연구가이루 어지고있으며(Kitajima, 1973; Ventura and Enderez, 1980;
Segawa and Yang, 1990; Balasubramanian and Bai, 1994).
물 벼룩(Moina macrocopa)
은대량배양의가능성이높은종중의 하나로고밀도사육이가능한종으로알려져있다(Jung et al., 2001; Shon, 2002; Kang et al., 2006).
그리고어류종묘생산의 초기먹이생물로서사용이시도된바가있어(Kitajima, 1973),
산업화가가능한먹이생물로기대되고있다.
따라서본연구는 어류종묘생산단계에서Artemia
급여에따른문제점을해결하기위해서
Artemia
대체먹이생물로서물벼룩을선정하여조피볼락자어에게급여하였을때성장과생존율에미치는영향을 알아보고자하였다
.
재료 및 방법
실험 물벼룩
실험에사용한물벼룩은국립경상대학교어류양식및사료연 구실에서보관중인
Moina macrocopa
종을사용하였다.
물벼룩 배양은30 L
투명원형아크릴수조를사용하여Seed culture
를 하였고,
직경1.6 m,
높이1 m
의수량1 ton FRP
원형탱크를사 용하여대량배양하였다.
배양에사용된먹이는DHA
를다량 함유하는해양미세조류Schizochytrium sp.
를사용하였다.
이 때배양수온은28.0℃
그리고pH 7.5
로유지하였다.
실험어 및 사육관리
실험은경남고성군삼산면소재의종묘배양장에서사육중이 던조피볼락자어를현장에서실시하였다
.
사육에사용된실험 어는종묘배양장에서부화시켜클로렐라로배양한로티퍼를2
주동안공급한조피볼락자어를사용하였다.
이때사용된조 피볼락자어의크기는평균체장11.2±0.36 mm
였다.
대조구는 알테미아급여구와실험구물벼룩급여구로하고,
각구당3
반 복구로1,000
마리씩수용하고6
주간사육하였다. 2
주마다성장(Body length, mm)
과생존율(Survival rate, %)
을측정하였다.
취급으로인한조피볼락치어의폐사를방지하기위하여10
마 리씩무작위추출하여바닥에모눈종이를붙인전용측정용기 에넣고사진을촬영하는사진법으로전장만을측정하는방법 을택하였다.
실험종료후에는전어체를사용하여Lipid class
와Fatty acid
를분석하였다.
사육장치
실험에사용한사육장치는유수식사육장치로서유입수의 첨가에의해서배설물이즉시사육수조밖으로배출될수있도 록하였으며
,
사육수조의크기는지름80 cm×
높이90 cm (
유 효수심60 cm)
의원형아크릴수조로수량은300 L,
주수량의 보충에의한순환률은5-6
회전/
일하였다.
사육수조에는에어 스톤을설치하여용존산소량은7 mL/L
를유지하였으며,
사육 기간중수온은20.3±0.8℃
였으며,
염분농도는32±0.5 psu
를유지하였다.
지방산 분석
총지질추출은
Bligh and Dyer
방법(1959)
에준하였다.
비커 에 균체5 g
을 취하여세포분쇄기(homogenizer AM-12, Ni- honseiki Kaisha Co. Ltd., Tokyo, Japan)
에서15,000 rpm
로5
분간분쇄한후, Chloroform
과Methanol
을2:1
로혼합한추출 용매를시료의2
배량넣어하루동안방치한다음chloroform
층만을분리하기위하여둥근플라스크위에깔때기를놓고,
그 위에Na
2SO
4를넣어서서히chloroform
층만흘러내리게하였 다.
분리된chloroform
층은진공회전농축기(Rotavapor R-114,
BUCHI)
를사용하여40℃
이하에서 용매를완전히증발시킨후
,
추출된총지질의무게를측정하였다.
모든작업은질소기 류하에서행하였다.
지방산methyl ester
유도체는시료일정량 과내부표준물질(C
23:0methyl ester) 1 mL (1 mg)
를cap tube
에 취하고, 0.5 N NaOH-methanol
용액1.5 mL
를가하여질소를 충진한다음, 100℃
에서8
분간가열하여검화하였다.
방냉후12% BF
3-methanol 2 mL
를가한후tube
의뚜껑을닫고, 100℃
에서
11
분간가열하여methyl
화하였다.
약30℃
로냉각한후Iso-octane 1 mL
를첨가하고30
초간vortex mixer
로혼합하였 다.
즉시3 mL
의포화식염수를가한다음흔들어방치하여iso- octane
층이분리되도록하였다. iso-octane
층을시료병(4 mL)
에옮긴후,
다시iso-octane 1 mL
를첨가한다음흔들어재추 출하여시료병에모으고이를지방산methyl ester
시료로하였 다.
지방산분석에사용하는GLC
는Omegawax
TM-320 fused- silica capillary column (30 m×0.32 mm×0.25 µm, i.d., Su- pelco Co., Bellefonte, PA, USA)
를장착한Clarus 600 (Perkin Elmer Co. Ltd., USA)
를이용하였다.
분석조건으로Column
은185℃
에서8
분간유지하고3℃/min
씩230℃
까지상승시킨후, 10
분간유지하였다.
이때주입기는250℃,
검출기는270℃
그 리고carrier gas
는He (1.0 kg/cm
2)
을사용하였다.
지방산의분 석은동일조건에서분석한표준품의ECL
과비교하여동정하 였고,
지방산표준품은14:0, 16:0, 18:1, 18:2, 18:3, 20:0, 22:1, 24:0 (Sigma Chemical Co., St. Louis, MO, USA)
과GC-MS
로동정된menhaden oil
을사용하였다.
Lipid class
Lipid classes
는TLC/FID
와Iatrorecorder TC-21 intergrator
가장착된
Iatroscan New MK-5 (Iatron Laboratory Inc., To- kyo, Japan)
를이용하여분석하였다.
분석조건에서공기의유 속은2 L/min
이며detector
수소량은160 mL/min
으로, scan- ning speed
는0.30 cm/s
로하였다.
분석과정은먼저Rod S-Ⅲ (0.9×150 mm,
석영봉규산코팅)
를5
분간수세한후,
다시증 류수10 mL
로헹군다음수분을증발시키기위하여아세톤10 mL
로씻고50℃
로조정한Rod-Dryer (TK-5 Iatron Lab. Inc., Tokyo, Japan)
에서5
분간건조시킨후Iatroscan
내에서수소염 이온화불꽃상에서3
회이상반복하여유기물을완전히제거하 였다. Rod
에시료1 μL
를Microdispenser (Drummond Scien- tific Co., Bromall, PA, USA)
로써점적하여전개조(NaCl
로포 화시킴)
에서10
분간포화시켰다.
전개용매는n-hexane : dieth- yl ether : acetic acid = 97:3:1 (v/v)
를이용하여약10 cm
까지 전개시킨후, Rod
를전개조에서꺼내고Rod-Dryer
에서5
분간 건조시켜서Iatroscan
으로분석하여지질획분의조성비를구 하고그함량을산출하였다.
동정은표준품인Cholesterol ester, Free fatty acid, Triglyceride, Cholesterol
및Phospholipid
에의 하여동정하였다.
통계처리
모든 자료는
3
회 반복 실험하여mean±SD
로 나타내었고, JMP (2002) Statistical Discovery Software™ version 5 (SAS Institute Inc., Cary, NC, USA)
를 이용하여Tukey-Kramer HSD test
로통계처리하였다.
결과 및 고찰
성장과 생존율
M. macrocopa
와Artemia
를조피볼락자어에게6
주간급여 한성장결과는Table 1
에나타낸바와같다.
실험초기조피볼 락크기는11.2 mm
에서M. macrocopa
와Artemia
를2
주간급 여한후,
각각18.2 mm
과15.0 mm
로성장차이가나타나기시 작하였다(P<0.05). M. macrocopa
와Artemia
를4
주간급여한 후에는각각32.0 mm
과21.1 mm
였으며, 6
주후에는각각45.6
mm
과25.7 mm M. macrocopa
를급여하였을때Artemia
급여 한실험구에서보다성장이우수한것으로나타났다(P<0.05).
M. macrocopa
와Artemia
급여에따른생존율은Table 1
에나타낸바와같이
Artemia
를급여한실험구의경우는실험시작후
2
주까지는생존율100%
였으나, 2
주가지난후부터는Arte- mia
를급여한실험구의폐사개체가나타나기시작하여4
주째 생존율은74.6%
로낮아졌으며(P<0.05), 6
주후에는생존율이12.8%
로급격히낮아지는것으로나타났다(P<0.05). Artemia
를 단독으로급여하였을때실험시작후2
주이후부터폐사 개체가나타나기시작했으며, 4
주째에는생존율이74.6%
로나 타났다.
이는부화후30-45
일사이로이시기에나타나는증 상으로는폐사까지는아니더라도생존하고있는개체들가운 데서일부는이시기이후부터바닥층으로들어눕기시작하 였다가사료급여시에는수면으로부상하는상태를나타내었 다.
일반적으로종묘생산현장에서는이문제를해결하기위하 여배합사료의전환을시도하는데이때부터는개체간의성장 차이와공식현상이나타나많은감모를일으키는시기이지만,
본실험에서는먹이를충분히공급하였기때문에공식현상은거의일어나지않은것으로사료된다
. Artemia
를급여한구에서는생존율이
12.8%
로나타난반면에M. macrocopa
를급여 한구의경우는실험시작후에4
주까지모두생존하였으며, 6
주째99.2%
의높은생존율을나타났으며0.8%
의폐사개체는 죽은개체를발견할수가없었기때문에영양상의결핍보다는 공식에의한현상이라고사료된다.
공급한M. macrocopa
는 담수물벼룩으로해수어류의종묘생산에적용함에있어서해수 에서의생존기간이8
분을초과하지못하여섭이후남은물벼 룩의사체가수질오염의우려등이제기되고있다.
그러나본 실험에사용한유영어류인조피볼락을대상으로실험하는동 안에물벼룩급여와동시에2
분이내에물벼룩을전량섭이하 였다. M. macrocopa
를급여하는즉시유속에따라서흐르는 물벼룩을섭이하기위해서조피볼락의군집이이동하면서M.
macrocopa
이바닥에도달하기전에전량섭이하는독특한기호성을나타내었다
.
이러한조피볼락의물벼룩에대한특이한 기호성으로보아활물벼룩이아닌냉동물벼룩의사용도충분 할것으로사료된다.
지질영양가
실험에서사용한
M. macrocopa
와영양강화한Artemia
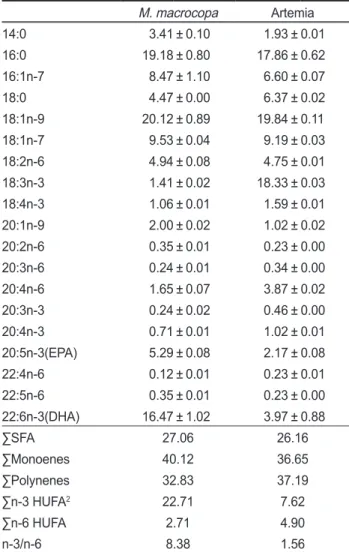
의지 방산조성은Table 2
에나타낸바와같다. M. macrocopa
의경 우는18:1n-9, 16:0, 22:6n-3, 18:1n-7
그리고16:1n-7
순으로높 게 나타났으며,
이때함량은각각20.12%, 19.18%, 16.47%, 9.53%
그리고8.47%
로나타났다.
영양강화Artemia
의경우 는18:1n-9, 18:3n-3, 16:0, 18:1n-7,
그리고16:1n-7
순으로높 게 나타났으며
,
이때함량은각각19.84%, 18.33%, 17.86%, 9.19%
그리고6.60%
로 나타났다.
포화지방산의 경우는M.
macrocopa
와Artemia
에서 각각27.06%
와26.16%
로 차이 Table 1. Growth and Survival rate of rockfish Sebastes schlegelifed on experimental diets for 6 week week
M. macrocopa Artemia
Body Length
(mm) Survival rate
(%) Body Length
(mm) Survival rate (%)
0 11.2±0.36 100 11.2±0.36 100
2 18.2±1.22 100 15.0±0.82 100
4 32.0±1.08 100 21.1±1.28 74.6
6 45.6±1.46 99.2 25.7±1.08 12.8
Values in the same row with same superscript are not significantly different (P<0.05).
가 없는 것으로 나타났다
(P<0.05). ∑n-3 HUFA
의 경우M.
macrocopa
와Artemia
에서 각각22.71%
와7.62%
로서M.
macrocopa
가높게나타났다(P<0.05).
두먹이생물간의차이는18:1
지방산계열과16:0
지방산이높아서먹이생물의일반적 인경향을나타냈으나, M. macrocopa
은DHA
함량이16.47%
로높은데비해서
Artemia
실험구는3.97%
로낮게나타났으 며(P<0.05),
영양강화를했음에도불구하고DHA
가낮게나타난것은
Artemia
자체의생리적현상인지방산역전환작용에의하여
DHA
가EPA
로역전환되었을가능성이있다(Barclay and Zeller, 1996).
또한Artemia
구에서18:1n-9
과18:3n-3
이양대지방산을이루고
18:3n-3
의지방산이특이적으로높은점은
Artemia
의특징을잘나타내고있다.
그러나먹이생물의지질영양가척도면에서
18:3n-3
의지방산의함량이높은것은담수어류에게는중요한성분이지만
,
참돔과같은해수어류에게는필수지방산으로의효과는매우낮은것
(Watanabe, 1993)
으로밝혀져있기때문에이점이Artemia
의한계점인것으로 생각한다.
두먹이생물의지방산종류에서중요한평가척도인∑n-3 HUFA
에서특히, DHA
함량에서차이가발생하는데이러 한차이점은두먹이생물의영양강화방법의차이에서오는것 으로사료된다. M. macrocopa
의배양방법은DHA
를다량함 유한Schizochytrium
을지속적으로급이하면서배양하여먹이 와영양강화제역활을동시에수행하는반면에Artemia
는부화 후유화오일에의한영양강화방법에서오는것으로사료된다. M. macrocopa
와Artemia
를급여한조피볼락자어의체지방 의지방산조성은Table 3
에나타낸바와같이M. macrocopa
를급여한경우16:0, 18:1n-9, 22:6n-3, 16:1n-7
그리고18:1n-7
순으로높게나타났으며,
이때함량은각각19.61%, 19.37%,
Table 2. Fatty acid composition of Moina macrocopa and Artemia
nauplii (% of total fatty acids)
M. macrocopa Artemia
14:0 3.41 ± 0.10 1.93 ± 0.01
16:0 19.18 ± 0.80 17.86 ± 0.62
16:1n-7 8.47 ± 1.10 6.60 ± 0.07
18:0 4.47 ± 0.00 6.37 ± 0.02
18:1n-9 20.12 ± 0.89 19.84 ± 0.11
18:1n-7 9.53 ± 0.04 9.19 ± 0.03
18:2n-6 4.94 ± 0.08 4.75 ± 0.01
18:3n-3 1.41 ± 0.02 18.33 ± 0.03
18:4n-3 1.06 ± 0.01 1.59 ± 0.01
20:1n-9 2.00 ± 0.02 1.02 ± 0.02
20:2n-6 0.35 ± 0.01 0.23 ± 0.00
20:3n-6 0.24 ± 0.01 0.34 ± 0.00
20:4n-6 1.65 ± 0.07 3.87 ± 0.02
20:3n-3 0.24 ± 0.02 0.46 ± 0.00
20:4n-3 0.71 ± 0.01 1.02 ± 0.01
20:5n-3(EPA) 5.29 ± 0.08 2.17 ± 0.08
22:4n-6 0.12 ± 0.01 0.23 ± 0.01
22:5n-6 0.35 ± 0.01 0.23 ± 0.00
22:6n-3(DHA) 16.47 ± 1.02 3.97 ± 0.88
∑SFA 27.06 26.16
∑Monoenes 40.12 36.65
∑Polynenes 32.83 37.19
∑n-3 HUFA2 22.71 7.62
∑n-6 HUFA 2.71 4.90
n-3/n-6 8.38 1.56
1HUFA: Highly unsaturated fatty acid (above 20 carbon fatty acid) Values are means of triplicate groups.
Table 3. Fatty acid composition of the rockfish Sebastes schlegeli fed different living food organisms (% of total fatty acids)1
Sebastes schlegeli M. macrocopa Artemia
14:0 3.37 ± 0.07 3.09 ± 0.03
16:0 19.61 ± 0.32 18.22 ± 0.22
16:1n-7 9.50 ± 0.03 6.04 ± 0.02
18:0 5.34 ± 0.02 6.57 ± 0.01
18:1n-9 19.37 ± 0.56 20.05 ± 0.36
18:1n-7 8.48 ± 0.08 7.22 ± 0.03
18:2n-6 7.27 ± 0.02 4.31 ± 0.02
18:3n-3 1.50 ± 0.01 19.05 ± 0.08
18:4n-3 1.54 ± 0.01 1.14 ± 0.03
20:1n-9 1.53 ± 0.10 1.10 ± 0.12
20:2n-6 0.22 ± 0.00 0.36 ± 0.01
20:3n-6 0.09 ± 0.00 0.13 ± 0.00
20:4n-6 1.55 ± 0.30 3.34 ± 0.32
20:3n-3 0.22 ± 0.00 0.39 ± 0.01
20:4n-3 0.50 ± 0.06 0.71 ± 0.02
20:5n-3(EPA) 5.71 ± 0.28 3.84 ± 0.14
22:4n-6 0.04 ± 0.00 0.11 ± 0.01
22:5n-6 0.22 ± 0.01 0.28 ± 0.01
22:6n-3(DHA) 13.94 ± 0.68 4.05 ± 0.96
∑SFA 28.32 27.88
∑Monoenes 38.88 34.41
∑Polynenes 32.80 37.71
∑n-3 HUFA2 20.37 8.99
∑n-6 HUFA 2.12 4.22
n-3/n-6 9.61 2.13
1Values are means of triplicate groups. 2HUFA: Highly unsaturated fatty acid (above 20 carbon fatty acid)
13.94%, 9.50%
그리고8.48%
였다.
그리고Artemia
를급여한 경우18:1n-9, 18:3n-3, 16:0, 18:1n-7,
그리고18:0
순으로높 게나타났으며,
이때함량은각각20.05%, 19.05%, 18.22%, 7.22%
그리고6.57%
였다.
포화지방산의경우는M. macroco- pa
와Artemia
에서각각28.32%
와27.88%
로차이를나타내지 않았으며, 20:5n-3(EPA)
와22:6n-3(DHA)
는M. macrocopa
에서각각5.71%
와13.94%
였으며, Artemia
는각각3.84%
와4.05%
였다.
특히∑n-3 HUFA
의경우M. macrocopa
와Arte- mia
에서각각20.37%
와8.99%
로서M. macrocopa
를급여한 구에서2
배이상높게나타났다(P<0.05).
각각의먹이생물을급 여한조피볼락체지방조성은섭이한먹이생물의지방산조성 과비슷한경향으로나타났으며,
이러한현상은해수어류의종 묘생산의필수지방산연구결과들과일치하고있다(Choi et al., 1999; Shon, 2002).
또한해산어류자치어의성공적인변태와 성장을하기위해서는DHA
가충분하게공급이되어져야만한 다고보고되고있으며(Watanabe, 1993), ∑n-3 HUFA
는어류 자치어용먹이의필수지방산공급원으로반드시필요한요소이며
, ∑n-3 HUFA
가풍부할때자치어의성장이빠르고체장증가의개선을가져온다고보고되고있다
(Koven et al., 1992;
Kang et al., 2006).
또한∑n-3 HUFA
는부레의정상적인수행 을하는것과관련이있는것으로밝혀졌으며, ∑n-3
지방산이 결핍된먹이는어류치어의성장을저해하고시각능력을손 상시키며,
신진대사에부조화를나타낸다고하였다(Castell et al., 1994).
최근에는∑n-3 HUFA
의대표적인지방산인EPA
와DHA
에서자치어의활력은EPA
보다DHA
의역할이더욱큰 것으로보고되고있다(Waranabe, 2008).
이러한점으로미루어 볼때본실험에서Artemia
구에서의성장과생존율이낮은점은DHA
함량의부족이하나의요인으로작용한것으로사료된다.
두먹이생물의
Lipid classes
조성은Fig. 1
에나타낸바와같이 극성지질인Phospholipids(PL)
함량과비극성지질의함량에서차이를나타내고있다
. PL
은M. macrocopa
와Artemia
는각각63.8%
와40.1%
로M. macrocopa
구가높게나타났다(P<0.05).
PL
은세포막의구성성분을이루고있는성분으로서인지질가 능성이매우높다.
이들인지질은해수어류자치어의중요한필 수지방성분으로삼투압조절에중요한역할을하는것으로알 려져있다(Watanabe, 2008).
또다른먹이생물인Copepoda
는Artemia
보다극성지질의비율이더높으며,
생리적으로중요한 지방산은극성지질에더많이존재하기때문에우수한먹이생 물로취급하고있으며(Shon, 2002),
이러한지질종류를많이함유하고있는천연산
Copepoda
를급여했을때성장이양호하게나타나는것도관련이있는것으로추정되고있다
(Kanazawa
et al., 1983; MacEvoy, 1998). Triglycerides (TG)
함량은M.
macrocopa
와Artemia
에서각각30.2%
그리고50.9%
로Arte- mia
급여구가높게나타났다(P<0.05). TG
는에너지원의성분 이면서중성지방의성분인점으로미루어보아,
부화한Artemia
유생의자체지질의성분보다는영양강화제의성분이기여한 것으로추정되는데이점에대해서는자세한연구가요망된다. Cholesterols (CHOL)
의함량의경우는M. macrocopa
와Arte- mia
에서각각5.9%
그리고4.0%
로M. macrocopa
구에서높게 나타났으며(P<0.05). Free fatty acid (FFA)
의경우는M. mac- rocopa
구와Artemia
구에서각각0.1%
그리고5.0%
로Artemia
를급여한조피볼락에서50
배높게나타났다(P<0.05).
이러한 차이는두먹이생물자체의유리지방산으로보기는어렵고,
영양강화과정중에서발생한것으로생각된다
. Artemia
유생의영양강화는산소부족에의한폐사를막기위하여강한에어레 이션과함께고수온에서이루어지기때문에
TG
의산화부산 물로생성된것으로추정되며,
또한유리지방산은생물에게독성을나타내기때문에
Artemia
를급여한조피볼락에게부정적영향을미친것으로생각된다
.
해수어류종묘생산시에먹이생 물의지질영양가의척도는먹이생물중에함유되어있는EPA
와DHA
의∑n-3 HUFA
함량과지질class
의극성지질함량이다(McEvoy et al., 1998; Watanabe, 1993).
이상의결과를종합해 볼때조피볼락의종묘생산시에Artemia
를단독으로사용했을 때의대량폐사원인은첫째, Artemia
자체의총지방산중에서 필수지방산인DHA
함량이낮은점,
둘째,
이를극복하기위하 여영양강화를실시하고있지만영양강화의효율의한계성,
셋 째,
영양강화과정중에생겨나는FFA
함량의증가넷째, Arte- mia
내의극성지질(PL)
이부족한점,
다섯째Artemia
자체의생 리적현상인DHA
에서EPA
로의역전환현상들이복합적으로 작용한결과로추정한다.
이번실험을통해서Artemia
가가지 고있는여러문제점들을해결할수있는대체먹이생물로서유 용한방안의하나로물벼룩을급여하였을때높은성장과생존 율그리고우수한영양학적인가치를가지는것으로나타났다.
References
Balasubramanian PR and Bai K. 1994. Utilization of anaerobi- Fig. 1. Lipid class profile of M. macrocopa and Artemia nauplii.
FFA, free fatty acid; TG, triglycerides; CHOL, cholesterols; PL, phospholipids. Vertical bars indicate mean±SD.
FFA TG CHOL PL
70 60 50 40 30 20 10 0
Wt/Wt, %
Moina macrocopa Artemia
cally digested cattle dung slurry for the culture of zooplank- ton, Daphnia similis Claus (Crustacea; Cladocera). Asian Fish Sci 7, 67-76.
Barclay W and Zeller S. 1996. Nutritional Enhancement of n-3 and n-6 Fatty Acids in Rotifers and Artemia Nauplii by Feed- ing spray-dried Schizochytrium sp. J World Aquaculture Soc 27, 314-322. http://dx.doi.org/10.1111/j.1749-7345.1996.tb 00614.
Bligh EG and Dyer WJ. 1959. A rapid method of total lipid ex- traction and purification. Can J Biochem Physiol 37, 911- Boehlert GW, Kusakari M, Shimizu M and Yamada J. 1986. En-917.
ergetics during embryonic development in the kurosoi, Se-
bastes schlegeli. J Exp Mar Biol Ecol 101, 239-256. http://
dx.doi.org/10.1016/0022-0981(86)90266-2.
Castell JD, Ghioni C and Sargent JR. 1994. Effects of purified diets containing different combinations of arachidonic and docosahexaenoic acid on survival, growth and fatty acid composition of juvenile turbot (Scophthalmus maximus).
Aquaculture 128, 315-333. http://dx.doi.org/10.1016/0044- 8486(94)90320-4.
Choi BD, Kang SJ and Kim KY. 1999. Mass production and nutritional value of Moina macrocopa in a various culture condition for Artemia substitution. The Third Asia-Pacific Marine Biotechnology Conference Philippines, O-2.
Corey PD, Leith DA and English MJ. 1983. A growth model for coho salmon including effects of varying ration allotments and temperature. Aquaculture 30, 125-143. http://dx.doi.
org/10.1016/0044-8486(83)90157-6.
Fujita S. 1973. Importance of zooplankton mass culture in pro- ducing marine fish seed for fish farming. Bull Plank Soc Jap 20, 49-53.
Hirayama K. 1985. Biological aspects of the rotifer Brachionus
plicatilis as a food organism for mass culture of seeding.
Coll France-Japan Oceanogr 8, 41-50.
Jung MM, Kim HS and Rho S. 2001. Survival and growth re- sponse on jumping of the each saline concentration of fresh- water cladoceran Moina macrocopa and estuarine cladocer- an Diapanosoma celebensis. Korean Fish Soc 34, 697-704.
JMP. 2002. Statistics and Graphics Guide. Version 5.0 SAS In- stitute Cary NC 179-209.
Kanazawa A, Teshima S, Kobayashi T, Takae M, Iwashita T and Uehara R. 1983. Necessity of dietary phospholipid for growth of the larval. Ayu Mem Fac Fish Kagoshima Univ 32, 115-120.
Kang CK, Park HY, Kim MC and Lee WJ. 2006. Use of ma- rine yeasts as an available diet for mass cultures of Moina
macrocopa. Aquacult Res 37, 1227-1237. http://dx.doi.
org/10.1111/j.1365-2109.2006.01553.x.
Kim NY, Choi BD, Kim KY and Kang SJ. 1999. Substitution live foods for Artemia II. Moina macrocopa production de- pend on the water temperature and fatty acid composition.
Preccdings of Korean Fisheries Society Autumn 1990, 363- Kitajima C. 1973. Experimental trials on mass culture of cope-364.
pods. Bull Plankton Soc Japan 20, 54-60.
Kitajima C. 1983. Mass culture-actual examples. In: The rotifer
Brachionus plicatilis biology mass culture. Jap Soc Sci Fish,
eds Koseisha Koseikaku, Tokyo, Japan, 102-128.Koven WM, Tandler A, Kissil WG and Sklan D. 1992. The importance of n-3highly unsaturated fatty acids for growth in larval Sparus aurata and their effect on survival, lipid composition and size distribution. Aquaculture 104, 91-104.
http://dx.doi.org/10.1016/0044-8486(94)90424-3.
Lee KW, Park HG, Lee SM, Han HS and Lim YS. 2004. Food value of freshwater rotifer (Brachionus calyciflorus) for cul- ture of sweetfish (Plecoglossus altivelis) larvae. J Kor Fish Soc 37, 7-12. http://dx.doi.org/10.5657/kfas.2004.37.1.007.
Lee SM, Lee JY, Kang YJ, Yoon HD and Hur SB. 1993a. n-3 highly unsaturated fatty acid requirement of the Korean rockfish Sebastes schlegeli. Bull Korean Fish Soc 26, 477- Lee SM, Lee JY, Kang YJ and Hur SB. 1993b. Effects of n-3 492.
highly unsaturated fatty acid on growth and biochemical changes in the Korean rockfish Sebasts schlegeli I. growth and body composition. J Aquaculture 6, 89-105.
Lee SM, Lee JY, Kang YJ and Hur SB , 1993c. Effects of n-3 highly unsaturated fatty acid on growth and biochemical changes in the Korean rockfish Sebastes schlegeli II. change of blood chemistry and properties of liver cells. J Aquacul- ture 6, 107-123.
McEvoy LA, Naess T, Bell JG and Lie O. 1998. Lipid and fatty acid composition of mormal and malpigmented Atlantic halibut (Hippoglossus hippoglossus) fed enriched Artemia:
a comparison with fry fed wild copepods. Aquaculture 163, 237-250. http://dx.doi.org/10.1016/s0044-8486(98)00237- Segawa S and Yang WT. 1990. Growth, moult, reproduction and 3.
filtering rate of an estuarine cladoceran Diaphanosoma ce-
lebensis, in laboratory culture. Bull Plankton Soc Japan 37,
145-155.Shon EJ. 2002. Nutritional lipids enrichment of water flea,
Moina sp. for seed production in marine fish. Gyeongsang
national university, master, Jinju, Korea.Ventura RF and Enderez EM. 1980. Preliminary studies on
Moina sp. production in freshwater tanks. Aquaculture 21,
93-96. http://dx.doi.org/10.1016/0044-8486(80)90129-5.Watanabe T. 1993. Importance of docosahexaenoic acid in ma- rine larval fish. J World Aquacult Soc 24, 152-161. http://
dx.doi.org/10.1111/j.1749-7345.1993.tb00004.x.
Watanabe T. 2008. Nutrition and feeding in fish and crustaceans.
Kousesha, Tokyo, Japan, 416.
Yamasaki S, Secor DH and Hirata H.1987. Population growth of two types of rotifer (L and S) Brachionus plicatilis at differ-
ent dissolved oxygen levels. Nippon Suisan Gakkaishi 53, 1303-1309.
Yoshimura K, Hagiwara A, Yoshiamtsu T and Kitajima C. 1996.
Culture technology of marine rotifers and the implications for intensive culture of marine fish in Japan. Mar Freshwater Res 47, 217-222. http://dx.doi.org/10.1071/mf9960217.