107
ABBREVIATIONS: PDGF, platelet-derived growth factor; RASMC, rat aortic smooth muscle cells; MAPK, mitogen-activated protein kinase; Hsp, heat shock protein; ERK, extracellular signal-regulated kinase; JNK, c-Jun N-terminal kinase; DMEM, dulbecco modified eagle medium; FBS, fetal bovine serum; OWE, olibanum water extract.
Corresponding to: Bokyung Kim, Department of Physiology, School of Medicine, Konkuk University, 322, Danwol-dong, Chungju 380-701, Korea. (Tel) 82-43-8403726, (Fax) 82-43-8519329, (E-mail) bkkim2@
kku.ac.kr
*Additional corresponding author: Dr. HM Lee, [email protected]
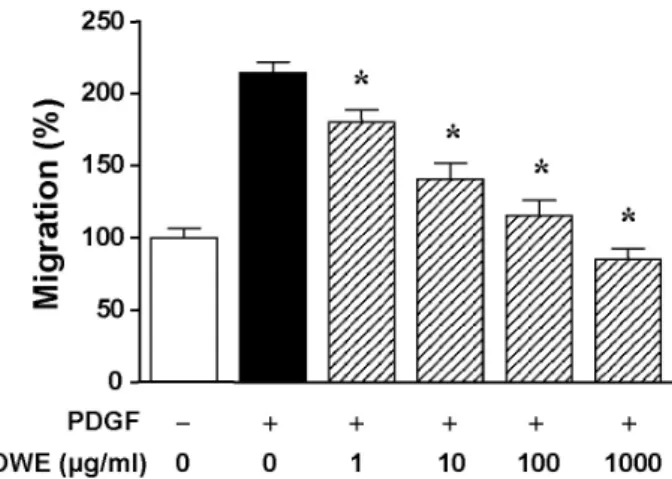
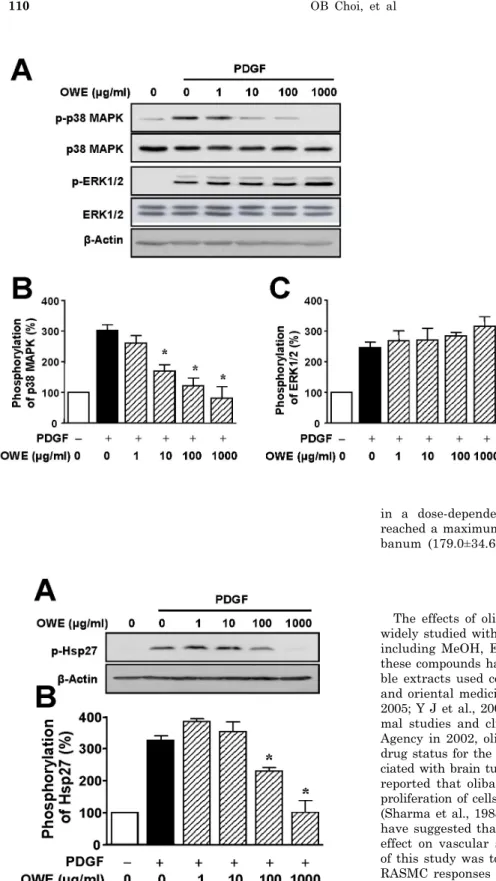
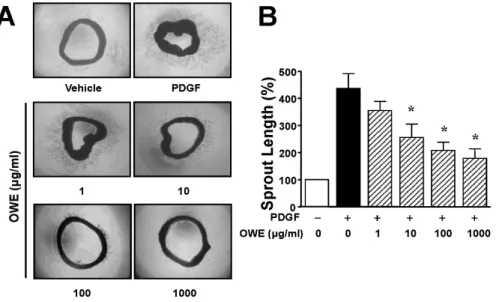
Olibanum Extract Inhibits Vascular Smooth Muscle Cell Migration and Proliferation in Response to Platelet-Derived Growth Factor
Ok-Byung Choi
1, Joo-Hoon Park
1, Ye Jin Lee
1, Chang-Kwon Lee
2, Kyung-Jong Won
2, Junghwan Kim
3, Hwan Myung Lee
1,*, and Bokyung Kim
21