10
책임저자:김성주, 서울시 강남구 일원동 50
135-710, 삼성서울병원 외과 Tel: 02-3410-3476, Fax: 02-3410-0040 E-mail: [email protected]
접수일:2007년 5월 7일, 게재승인일:2007년 8월 29일
중심 단어: 제대혈로부터 유래된 CD34+ cell, 태아의 흉선과 간 조 직, Rag2/γc/
생쥐, 인간화 생쥐 모델
태아의 간과 흉선 조직 그리고 조혈모 세포를 이식한 Rag2
/γ
c/Mice에 사람의 면역 세포 형성
성균관대학교 의과대학 삼성서울병원 1외과학교실, 2삼성생명과학연구소, 3펩트론(주)
강미진2ㆍ주성연2ㆍ최봉금2ㆍ정다연2ㆍ최호인3ㆍ박재범1ㆍ최규성1ㆍ권준혁1ㆍ김성주1ㆍ조재원1
Reconstitution of Human Immune Cells with Co-transplantation of Fetal Liver/Thymus Tissues and Cultured Umbilical Cord Blood-derived Hematopoietic Stem Cells in Rag2
/γ
c/Mice
Mijin Kang2, Sung-Yeon Joo2, Bong-Kum Choi2, Da- Yeon Jung2, Ho-In Choi3, Jae Berm Park1, Gyuseong Choi1, Choon Hyuck Kwon1, Sung-Joo Kim1 and Jae- Won Joh1
1Department of Surgery, Samsung Medical Center, Sung- kyunkwan University School of Medicine, 2Samsung Biosci- ence Research Institute, Seoul, 3Peptron, Inc., Daejeon, Korea
Purpose: Many researchers have tried to develop animal models that mimic the human immune system, e.g. a humanized mouse model, to improve the engraftment of hematopoietic stem cells and develop human immune cells in an animal model. This study evaluated the feasibility of the cultured human umbilical cord blood (hUCB)-derived CD34+ cells for cell expansion, in Rag2/γc/ mice, and establish co-transplantation with human fetal thymus/liver tis- sue (Thy/Liv) under the kidney capsule.
Methods: Co-transplantation of hUCB-derived CD34+ cells with Thy/Liv was performed. The hUCB-derived CD34+ cells were prepared by freshly thawing (G1) and culturing for 7 days with two types of cytokine combinations (G2, G3). The CD45+ cell populations were measured at 6, 8, 10 and 16 weeks in the peripheral blood. The splenocytes were cultured with mitogenic stimuli (PHA -L or IL-2) at 20 weeks post- transplantation, and the proliferation of human immune cells was evaluated.
Results: There were no significant differences in the human CD45+ cell populations at 6, 8, 10 and 16 weeks post-trans- plantation between the groups. In the cultured splenocytes at 20 weeks post-transplant with PHA-L or IL-2, there was remarkable expansion of CD3+ cells in the three groups.
Although no CD19+ cells were detected in the spleen, human Ig G was detected in the sera of these mice.
Conclusion: The cultured and expanded hUCB-derived cells with cytokine combinations might be a feasible cell source in humanized mouse modeling. In addition, human immune cells can be reconstituted from the co-transplantation of Thy/Liv and cultured hUCB-derived CD34+ cells. (J Korean Surg Soc 2008;74:10-18)
Key Words: Cord blood-derived CD34+ cell, Fetal thymus/
liver tissues, Rag2/γc/ mice, Humanized mouse model
서 론
질병에 대한 치료를 연구하는 방법으로, 사람과 유전적 으로 매우 유사하다고 알려진 동물에서 인위적인 사람 질 병 모델을 구축함으로써 많은 연구가 이루어져왔다. 그러 나 이런 동물 질병 모델은 치료 방법을 찾더라도 그것을 사람에게 임상 적용하기 전까지는 많은 단계를 거쳐야 하 며, 사람에게 임상 적용 시에 동일한 결과를 보장할 수 없다 는 제한점을 가지고 있다.
1980년대에 면역 세포가 결핍된 생쥐가 개발되면서, 사 람의 면역 체계 형성 및 면역 반응을 갖는 동물 모델에 대 한 연구로 가시화되기 시작했으며, 사람의 면역 체계를 갖 춘 동물 모델의 개발은 질병의 병태 생리뿐 아니라 치료법 및 치료에 대한 반응 연구에 획기적인 기여를 할 것으로 기대되어 왔다.(1) 이에 면역 결핍 생쥐에서 사람의 면역체 계를 갖춘, humanized-mice의 구축은 동물에서의 질병 연구 의 결과를 사람에게로 임상 적용할 수 있는 직접적인 근거 를 제공하여 임상 적용단계를 획기적으로 줄여줄 수 있다 는 점에서 반드시 필요한 연구과제로 여겨진다.
이러한 연구로 Severe combined immune deficiency (SCID) 생쥐에게 사람 CD34+ 세포를 이식한 결과 모든 생쥐의 조 직에서 사람으로부터 유래된 조혈모세포가 소량 발현되었 으며 조직적으로 재형성되어 있음이 확인되었다.(2) 그러나 이식한 생쥐에 형성된 소량의 사람 세포의 대부분이 B 세 포로, 모든 조직에서 약 90% 정도 차지하며 이식된 SCID 생쥐의 어떠한 조혈 기관에도 사람 세포인 T 세포가 전혀 존재하지 않았다. 이러한 점을 개선하기 위하여 SCID 생쥐 대신, natural killer 세포의 기능도 저하되어있는 non-obese diabetic/SCID (NOD/SCID) 생쥐를 이용한 연구들이 이루어 졌다. 그러나 NOD/SCID 생쥐에게 사람 CD34+ 세포를 이식 을 했을 때, SCID 생쥐에서보다 훨씬 더 많은 사람 세포를 조직적으로 재형성하였음(3-5)에도 불구하고 여전히 사람 T 세포가 발달하지는 않았다.(2,6) 사람 CD34+ 세포를 이식 한 SCID 또는 NOD/SCID 생쥐에서 사람 T 세포가 결여되어 있는 것과는 대조적으로 태아의 흉선과 간 조직을 이식한 SCID 생쥐에서는 사람 T 세포가 많이 형성되었다. 그러나 태아의 흉선과 간 조직을 이식한 SCID 생쥐에서도 비장을 제외한 다른 곳에서는 사람 T 세포가 1% 미만으로 소량 존 재하였으며 사람 B 세포는 거의 발현되지 않았으며 사실상 사람 세포가 흉선에만 제한되어 존재하였다.(7-9) 최근 NOD/
SCID 생쥐에 태아의 흉선과 간 조직을 먼저 이식한 후 간 조직으로부터 유래한 CD34+ 세포를 주입한 humanized mice 모델이 보고가 되었다.(10) 이 체계에선 사람의 T 세포와 B 세포뿐 아니라 그 외에 수지상세포(dendritic cell), 단핵구 (monocyte)와 대식세포(macrophage)도 생성이 확인되었다.
또한 in vivo상에서 Epstein-Barr virus (EBV)와 초항원인 tox- ic shock syndrome toxin 1 (TSST-1)에 대해 특정 면역 반응 을 확인한 바 있다.
사람 CD34+ 세포의 원천으로 간 조직으로부터 얻는 사람 CD34+ 세포는 그 양이 소량이며 태아의 조직을 구하는 것 은 매우 제한적이다. 사람 CD34+ 세포의 또 다른 근원으로, 최근에 이식과 유전자 치료 적용에 조혈모세포의 원천으로 써 주목을 받고 있는 제대혈을 들 수 있다. 제대혈은 발달 초기의 조혈모세포의 상당 부분을 포함하고 있으며 골수 줄기 세포보다 더 높은 확장과 잠재적인 증식성을 가지고, 기증자와 수혜자의 조직 적합성 항원(human leukocyte anti- gen)의 불일치 정도가 크더라도 면역 거부 반응이 잘 일어 나지 않는다는 점에서 장점을 가지고 있다. 또한 이식편 대 숙주병(graft-versus-host disease) 역시 골수를 이식하였을 때 보다 제대혈 이식 시 낮은 발생률을 보인다. 그러나 하나의 제대혈로는 세포 수가 충분치 않다는 한계가 있다. 따라서 제대혈로부터 CD34+ 세포를 분리 시, 생체 밖에서(ex vivo) long-term repopulating 할 수 있는 성질을 가지면서도 조혈 모세포의 확장과 자기 재생을 지지해 줄 수 있는 배양 조건 을 찾아내는 것이 중요하다.
배양 조건의 연구로, 조혈모세포의 발달이 사이토카인
수용체의 신호에 의해 조절된다는 점에 착안하여 많은 연 구자들이 조혈모세포의 확장에 있어 최적 배양 조건을 획 득하기 위한 여러 종류의 사이토카인 조합을 시도하였 다.(11,12) 특히 조혈모세포의 발달에 중요한 신호 전달을 한다고 알려진 stem cell factor (SCF)와 Flk2/Flt3 ligand (FL), 발달 초기의 조혈모세포의 확장을 자극시킨다고 알려진 thrombopoietin (TPO)은 조혈모세포 확장에 중요한 사이토 카인으로 잘 알려져 있다. 이 외에 다양한 사이토카인들을 혼합하여 NOD/SCID 생쥐에 이식한 결과 상당히 높은 비율 로 사람 CD45+ (common leukocyte marker) 세포가 감지되었 으며 배양된 세포가 자기 재생을 하는 능력을 유지하고 있 다는 보고가 잇따르고 있다.(11,13)
이에 저자는 재조합 활성 유전자 2 (recombinase activating gene 2)와 많은 사이토카인의 기능적 수용체에 대해 공통적 인 감마(gamma) 체인이 모두 결핍(common gamma γc/Rag2 double knockout mice)된, 즉 T 세포와 B 세포 그리고 NK 세포가 결핍되어 면역 거부 반응을 일으키지 않는 생쥐를 이용하여, 이 생쥐에게 태아의 흉선과 간 조직을 이식하는 것과 동시에 제대혈로부터 분리한 CD34+ 세포를 사이토카 인의 혼합 배양 후 꼬리 정맥을 통해 주입하여 더 높은 이 식 성공률과 더불어 사람의 면역 체계가 형성되는지 연구 하고자 하였다.
방 법
1) 실험동물
C57BL/6J x C57BL/10SgSnAi-[KO]c-[KO] Rag2 female mice (Taconic Inc, USA)는 국제 실험동물관리공인협회(AAALAC International, Association for Accreditation of Laboratory Animal Care International)의 인증을 취득하여 국제적으로 요구하는 실험동물 관리 및 사용에 관한 지침[ILAR Guide, 1996]에 따라 운영 중인 삼성생명과학연구소 실험 동물실 SPF (specific pathogen free) 사육실에서 사육되었다.
제대혈로부터 분리한 조혈모세포(CD34+ 세포)를 사이토 카인 혼합 배양(G2 및 G3)하여 생쥐에 주입하였을 때, 배양 하지 않은 fresh CD34+ 세포를 주입(G1)하는 것에 비해 이 식 성공률 및 사람 면역 체계 형성에서 어떠한 차이가 발생 하는 지 알아보고자 하였다. 전체 그룹에 대한 대조군으로 PBS (phosphate buffered saline)를 주입하였다(Table 1).
2) 태아의 흉선, 간 조직 및 제대혈의 준비
생쥐에 이식할 세포는 다음과 같이 준비하였다. 이식할 태아의 흉선과 간 조직은 advanced Bioscience Resources, Inc (Alameda, CA)로부터 받았으며. 동의서를 받은 건강한 산모 (삼성서울병원)로부터 제대혈을 얻었다. 제대혈은 Ficoll- Hypaque density gradient centrifugation 방법으로 단핵세포 (mononuclear cell)만을 분리하였다. CD34+ 세포만을 얻기
Table 1. Humanized mice modeling group
Control Only PBS
G1 Fetal thymus/liver+ fresh UCB*-derived CD34+ cells G2 Fetal thymus/liver+ cultured (combination 1†)
UCB*-derived CD34+ cells
G3 Fetal thymus/liver+ cultured (combination 2‡) UCB*-derived CD34+ cells
*UCB = umbilical cord blood, †combination 1: α-MEM me- dium+20% FBS+1% deionized fraction V BSA+1% penicillin and 1% streptomycin+cytokines1 (100 ng/ml SCF, 10 ng/ml TPO, 100 ng/ml FL, 10 ng/ml G-CSF, 10 ng/ml GM-CSF, 2 U/ml EPO),
‡combination 2: α-MEM medium+20% FBS+1% deionized frac- tion V BSA+1% penicillin and 1% streptomycin+cytokines2 (100 ng/ml SCF, 10 ng/ml TPO, 100 ng/ml FL).
위해 Direct CD34 Progenitor Cell Isolation Kit (Miltenyi Biotec GmbH Bergisch Gladbach, Germany)를 사용하였다. 단 핵세포 1×108 세포 당 100μl blocking buffer와 100μl CD34 bead를 첨가하고 40분간 4oC에서 반응시켰다. 반응 후, AutoMACS (Miltenyi Biotec GmbH Bergisch) 기계를 이용하 여 CD34+ 세포만을 획득하였다. CD34+ 세포의 순도 (hLin hCD34+) 측정은 유체세포 측정법(Flow Cytometry)을 이용 하였으며 90∼95%였다. 획득한 사람 CD34+세포를 냉동 보 존액(freezing solution; 90% FBS+10% DMSO)에 침수시켜 freezing container에 넣고 70oC에서 overnight 한 후 액체 질 소에 보관하였다. 실험에 사용 시, 37oC water bath에서 녹인 후 실험에 사용하였다.
3) 사람 CD34+ 세포의 배양
이렇게 분리한 CD34+ 세포의 수 제한을 해결하기 위해 냉동보존을 하였던 사람 CD34+ 세포를 해동시켜 배양을 하 였다. 조혈모세포 확장에 따른 최적 배양 조건을 찾기 위해 2가지 사이토카인 조합(combination 1, 2)을 이용하였다(G2, G3). 기존에 조혈모세포 확장에 중요하다고 알려진 SCF, FL, TPO, 그 외에 erythropoietin (EPO), granulocyte colony stimulating factor (G-CSF), granulocyte-macrophage colony stimulating factor (GM-CSF) 등의 사이토카인들(R&D sys- tems, USA)을 첨가하였다. 24 well에 2×104 CD34+ cell을 분 주하고 사이토카인을 첨가하여 7일 동안 37oC, 5% CO2 배 양기에서 배양하였다. 배양 후, 그들의 순도 검사 (hLin hCD34+)는 제대혈 분리할 때와 동일하게 수행하였으며, 세 포 배양 시, combination 1은 초기 세포 수에 비해 세포 수 증가가 약 40∼50배 정도, combination 2는 약 20∼30배 정 도 증가가 되었다. CD34+ 세포의 발현(hLinhCD34+)은 com- bination 1에서는 40%, combination 2는 20∼30% 정도로 나 타났다. 이 때 사용한 배지는 α-MEM medium에 20% FBS,
1% deionized fraction V BSA, 1% penicillin과 1% streptomy- cin가 첨가된 것을 사용하였으며 사이토카인 조합은 Table 1과 같았다.
4) 사람 CD34+ 세포의 이식
이식하기 전 6∼8주 된 생쥐들을 치사량 미만인 3 Gy로 방사선을 조사하였다. 24시간 안에 생쥐를 마취하고, 1 mm3 로 자른 태아의 흉선과 간 조직을 왼쪽 콩팥 캡슐에 이식을 하였다. CD34+ 세포의 이식은 냉동 보존하였던 세포를 해 동만하여(G1) 또는 해동 후 combination 1과 2의 사이토카인 조합에서 7일간 미리 배양하여(G2, G3), 2×105개의 CD34+ 세포를 태아의 흉선과 간 조직 이식과 동시에 꼬리 정맥에 주입하였다. 대조군은 조직을 이식하지 않은 왼쪽 콩팥 캡 슐을 실험군과 동일하게 수술만 하였으며 꼬리에는 PBS를 주입하였다. 이식 후 생쥐들에게 항생제(Ciprofloxacin, 100 mg/L)을 경구 투약하였다.
5) 이식 후 말초 혈액 검사를 통한 이식 성공률의 비교 이식 후, 생쥐의 꼬리에서 6주, 8주, 10주, 16주 채혈하였 으며, 채혈 후 lysing buffer (BD Pharm LyseTM, BD bio- science, USA)를 이용하여 적혈구를 제거하고 hCD45+ 세 포 및 multilineage 세포들의 발현을 알아보기 위해 다양한 CD 표지들로 염색을 하고 유체세포 측정법으로 분석하였 다.
6) 골수 및 비장에서의 이식 성공률의 비교
이식 후 20주에는 생쥐를 희생하여 제1차 림프기관인 골 수와 제2차 림프기관인 비장으로부터 세포를 획득하였다.
비장의 경우, 슬라이드를 이용하여 조직을 부수고 nylon fil- ter에 통과시켜 잔해를 제거하였다. 원심분리하고 얻은 pel- let에 lysing buffer를 넣고 세포를 풀어준 후 상온에서 5분간 방치하였다. PBS를 넣어 반응을 멈추게 하고 상온에서 5분 간 원심분리하여 비장림프구를 얻었다. 얻은 세포의 일부 는 다양한 CD 표지들로 염색을 하고 나머지는 배양을 하였 다. 골수는 주사기를 이용하여 골수 안에 존재하는 세포를 얻어 염색을 하고 유체세포 측정법으로 분석했다: FITC- conjugated anti-human CD45, CD3; PE-conjugated anti-human CD19; PEcy5-conjugated anti-human CD56 and anti-mouse CD45 antibody.
7) 항원 자극에 대한 사람 면역 세포의 반응 실험 생쥐의 면역 체계에서 실제로 성숙한 사람 T 세포를 생산 해내는지 확인하기 위해, 이식 20주에 사람세포와 생쥐 세포 를 분리하지 않은 비장세포를 5×105 cells로 분주하고 Phytoagglutinin-L (PHA-L; Sigma, USA) (1μg/ml) 또는 human recombinant IL-2 (R&D systems, USA)로 7일 동안 자극을 주 고 후에 CD 표지들로 염색을 하고 유체세포 측정법으로 분
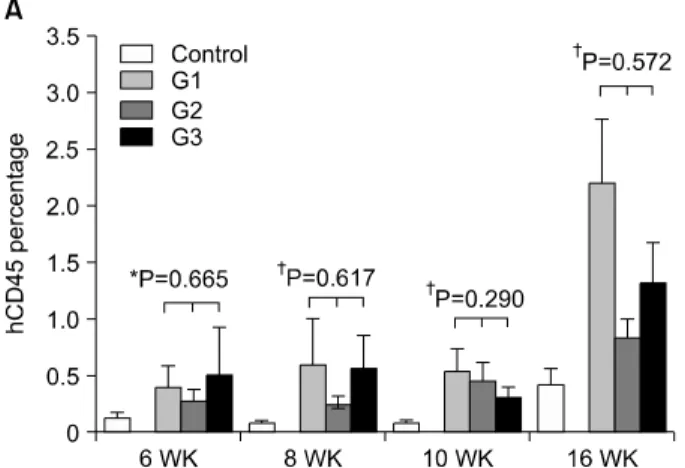
Fig. 1. Human cell reconstitution of recipient peripheral blood. (A) Time course of hCD45+ cells of peripheral blood from recipients after transplantation. Expression of hCD45+ cells was gradually increasing with time. (B) Phenotypic analysis of peripheral blood at 16 weeks after transplantation. Only hCD3+ T cells were present in all recipients. hCD45+ cells were analyzed using flow cytometry.
석하였다: FITC-conjugated anti-human CD3; PE-conjugated an- ti-human CD19; PEcy5-conjugated anti-human CD56 antibody.
PHA는 당에 붙는 렉틴(lectin)이라고 불리는 단백질의 한 종 류로써, 림프구 배양에서 세포 증식의 자극을 하는데 사용되 며, 특히 그 중 PHA-L은 T-림프구의 기능을 분석할 때 매우 유용한 시약이다. IL-2는 면역 체계의 넓은 범위에 작용하며 면역 활성화 조절과 항상성 조절에 중요한 역할을 하며 IL-2 역시 T 세포 성장 인자로 잘 알려져 있다.(14)
이식 후 20주에 심혈을 채혈하여 그로부터 혈청을 얻고 그 안에 존재하는 면역 글로불린을 조사하였다. Anti-human Ig G-Fc 특이적 항체를 10μg/ml 농도로 0.1 M sodium carbo- nate-bicarbonate buffer (pH 9.5) 희석한 후 100 ul씩 NUNC maxisorp plate에 넣고 4oC에서 overnight 반응시키며 coating 하였다. 반응 후 PBS-T (tween-20 0.05%)로 3회 씻어낸 후 0.5% Bovine Serum Albumin (BSA)가 포함된 PBS-T를 400 μl/well씩 넣고 37oC에서 1시간 동안 blocking 시켰다. Human Ig G standard를 0.5% BSA가 포함된 PBS-T에 serial dilution 하고, 채취한 혈청도 0.5% BSA가 포함된 PBS-T에 serial di-
lution하여 준비하였다. 준비된 시료를 각 well당 100μl씩 넣고 상온에서 1시간 동안 반응시키고, 이를 PBS-T로 3회 씻은 후 HRP labeled anti-Human IgG-Fab를 0.2μg/ml 농도로 0.5% BSA가 포함된 PBS-T에 희석한 후 100μl/well씩 넣고 상온에서 1시간 동안 반응시켰다. 마지막으로 이를 PBS-T 로 3회 washing한 후 안정화된 Tetramethylbenzidine (TMB) substrate를 100μl씩 넣어준 후 20∼30분간 반응시키고 2N HCl을 100μl/well씩 넣고 반응을 정지시키고 이를 ELISA reader에서 Optical Density (OD)450을 측정하였다.
8) 통계적인 분석
통계적인 분석은 Statistical Package for Social Science (SPSS for Window 15.0; SPSS Inc, Chicago, IL)를 이용하였 다. 각 개체 및 각 군의 주 수에 따른 사람 CD45+ cells에 대한 분석으로 Repeated Measure Analysis of Variance (ANOVA)를 이용하였다. 각 주 수에서 군 간의 비교는 일원 배치 분산분석(one way analysis of variances, ANOVA) 또는 Kruskal-Wallis test를 시행하였다. P value<0.05를 통계적으
Fig. 2. Establishment of Humanized-mice models. We sublethally irradiated the mice. Within 24h, we transplanted them with fetal liver and thymus tissues under the kidney capsule and intravenously with fresh or cultured 2×105 CD34+ cells. The left kidney capsule was changed by implanted tissues at 20 weeks after transplantation.
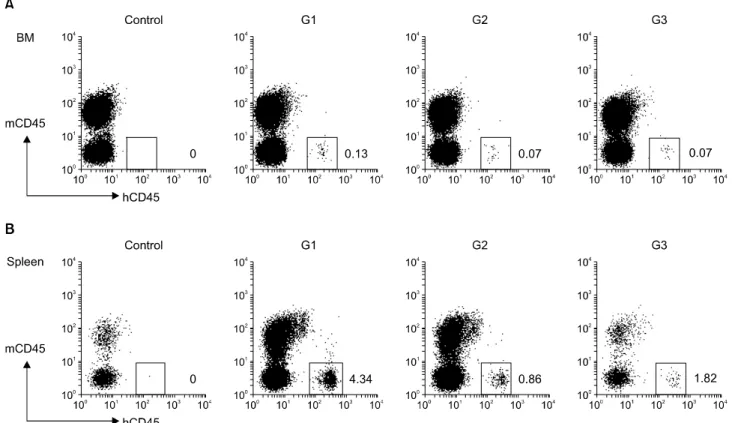
Fig. 3. Human cell reconstitution of recipient lymphoid organs at 20 weeks after transplantation. (A) Expression of hCD45+ cells in primary lymphoid organ BM is shown. (B) Expression of hCD45+ cells in secondary lymphoid organ spleen is indicated. hCD45+ cells in Spleen were expressed more than compared to BM. hCD45+ cells were analyzed by flow cytometry.
로 유의한 것으로 판단하였다.
결 과
1) 이식한 생쥐의 말초 혈액에서 사람 세포 존재 각 개체는 주 수에 따라 사람 CD45+ 세포가 통계적으로 유의하게 차이가 있었다(Fig. 1A) (P<0.001, Pillai's trace 및 Wilks' Lamda). 그러나 각 군의 주 수 에 따른 사람 CD45+ 세포는 통계적으로 유의한 차이를 나타내지 않았으며 (P=
0.638, Pillai's trace; P=0.664, Wilks' Lamda), 각 군에 따른 사람 CD45+ 세포는 통계적으로 유의한 차이를 나타내지는 않았다. 또한 각 주 수에서, 군 간의 차이를 비교하였을 때, 군 간의 차이는 각각 통계적으로 유의하지 않았다(6주
P=0.665, Kruskal-Willis Test, 8주 P=0.617; 10주 P=0.290; 16 주 P=0.572, ANOVA). 비록 통계적인 수치로 각 그룹 간의 차이가 유의하지는 않으나, 각각의 humanized mouse 모델 링 조건하에서 사람 CD45+ 세포가 형성되었다는 것을 확인 할 수 있다. 또한 각 개체가 주 수에 따라 통계적으로 유의 하게 차이가 나는 것으로 시간이 지남에 따라 사람 CD45+ 세포의 수가 증가했다는 것을 알 수 있다.
10주 이후로는 조혈모세포의 분화를 조사하였다. 사람 CD45+ 세포들이 발견되는 점을 미루어 림프구 계통과 mye- loid 계통의 하부 세포들이 존재할 것이라고 예상되어 다양 한 CD 표지들로 염색을 하였다. 16주에 모든 실험군에서 림파구의 사람 CD3+ T 세포들이 발견되었다. 사람 CD3+ T 세포들이 G1 그룹에서 1.45%, G2 그룹이 0.67%, G3 그룹이
1.47%로, G1 그룹과 G3 그룹은 거의 동일한 양이 발현되었 으며 G2 그룹에 비해 약 2.2배 정도 많은 양이 존재하였다 (Fig. 1B). 사람 CD19와 사람 CD56도 염색을 하였으나 감지 되지 않았으며 myeloid 계통의 세포들도 전혀 감지되지 않 았다.
2) 이식한 생쥐의 장기에서 사람 세포 존재
우선 이식 후, 20주에 태아의 조직이 이식 된 생쥐의 콩팥 을 육안으로 살펴보았다(Fig. 2). 태아의 간과 흉선 조직을 이식한 세 그룹 모두 왼쪽 콩팥 일부분이 기존의 선홍색이 아닌 흰색을 띠며 지방과 달리 단단하였다. 또한 오른쪽 콩 팥에 비해 상당히 부풀어 있는 것을 관찰할 수 있었다. 대조
Fig. 4. Human immune cell reconstitution in environment of Rag2-/-γc-/-
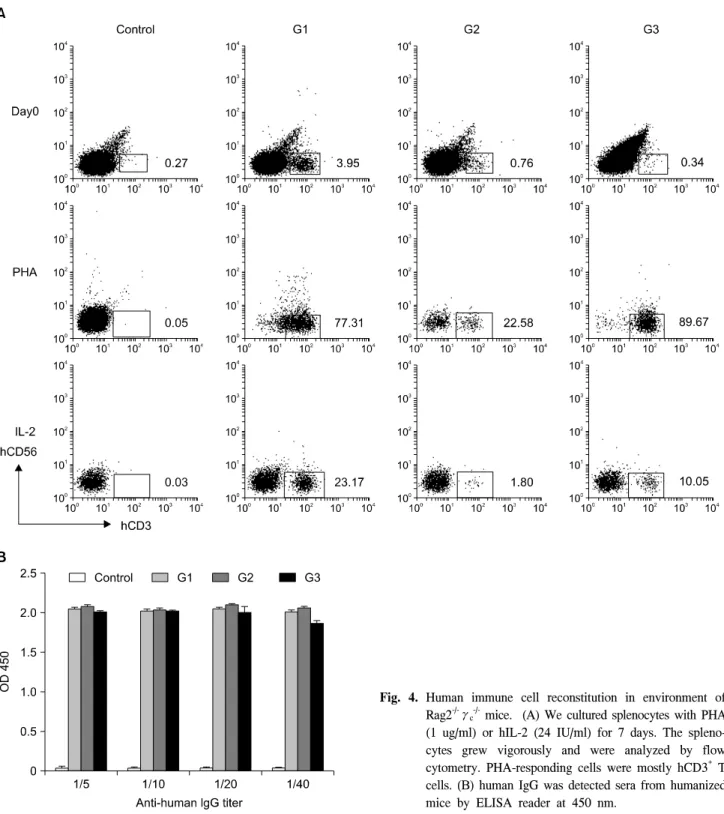
mice. (A) We cultured splenocytes with PHA (1 ug/ml) or hIL-2 (24 IU/ml) for 7 days. The spleno- cytes grew vigorously and were analyzed by flow cytometry. PHA-responding cells were mostly hCD3+ T cells. (B) human IgG was detected sera from humanized mice by ELISA reader at 450 nm.
군의 콩팥은 건강한 생쥐인 B6의 콩팥과 유사하였으며 정 상적이다.
이식한 생쥐의 장기에서 사람 세포 존재를 확인하기 위 해 우선 골수 세포를 살펴보면(Fig. 3A), 전반적으로 사람 CD45+ 세포가 1% 미만으로 상당히 낮게 존재하지만 대조 군에서 전혀 발현되지 않는 것과 달리 나머지 실험군에서 는 특정 집단을 형성하는 것을 관찰할 수 있다. G1 그룹에 서는 0.13%로, 재형성이 제대혈로부터 분리된 CD34+ 세포 를 배양한 세포를 주입한 그룹 둘 다(0.07%)에서보다 약 2 배 정도 재형성이 잘 되는 것을 관찰하였다. 비장림프구에 서는 사람 CD45+ 세포가 G1 그룹이 가장 높은 4.34%, G2 그룹에서는 0.86%, G3군에서는 1.82%로 전반적으로 골수 세포에 비해 12∼33배 정도 높게 발현이 되었으며 배양한 군 간에는 G3 그룹이 약 2배 정도 더 많이 발현이 되었다 (Fig. 3B).
3) 사람 면역 세포의 자극에 대한 반응
생쥐 체계에서 실제로 성숙한 사람 T 세포를 생산해내는 지 확인해 보았다(Fig. 4A). 우선 PHA-L 또는 hIL-2로 자극 하기 전에 여러 종류의 CD 표지들로(CD3, CD19, CD56) 염 색하였지만 말초혈액에서처럼 전혀 감지가 되지 않았고 비 장 림프구에서 사람 CD3+ T세포들만이 발견되었다. 대조군 에서 0.27%로 G3 그룹이 0.34%인 것에 비해 차이가 별로 나지 않는다. 그러나 대조군에서는 특정 집단을 형성하지 않는 것과 달리 G3 그룹에서는 발현 양이 적지만 특정 집단 을 형성하는 것을 볼 수 있다. G1 그룹에서는 3.95%, G3 그 룹에서는 0.76%가 발견되었다. 비장세포에 PHA-L (1μg/
ml) 또는 hIL-2 (24 U/ml)로 7일 동안 자극을 주고 후에 CD marker로 염색을 하여 유체세포 측정법으로 분석하였을 때, G1 그룹에서 77.31%, G2 그룹에서는 22.58%, G3 그룹에서 는 89.67%로 모든 실험군에서 T cell의 발현이 증가하였다.
특히 G3 그룹은 사람 CD3+ 세포가 0.34%밖에 존재하지 않 음에도 불구하고 생체 밖에서(in-vitro) PHA-L에 의한 자극 에 대해 반응이 89.67%로 매우 효과적이었다. 가장 높게 hCD3+ 세포를 발현한 G1 그룹에서는 초기에 비해 19.6배 정도 증가하였고 G2 그룹은 초기에 비해 약 30배 정도 증가 가 되었다. 또한 IL-2 처리 7일 후에는 G1 그룹에서 23.17%, G2 그룹에서는 1.80%, G3 그룹에서는 10.05%로 발현이 증 가하였다. PHA-L에 반응하는 세포와 유사하게, G3 그룹에 서 29.6배로 가장 높게 증식이 일어났으며 G1 그룹은 5.9배, G2 그룹은 2.4배로 나타났다. 태아의 조직들과 더불어 CD34+ 세포를 주입한 생쥐로부터 polyclonal T 세포가 생성되었으 며, 이러한 T 세포가 활발하게 증식하는 것과 더불어 기능 적인 반응을 한다는 것을 보여주었다.
또한 생쥐의 심혈로부터 혈청을 얻고 사람 면역글로불린 이 존재하는지 살펴보았다(Fig. 4B). Anti-human IgG-Fc 특 이적 항체를 이용하여 자극을 준 후, ELISA reader로 OD값
을 측정한 결과 면역 글로불린 IgG가 검출되었다. 면역 결 핍 생쥐의 배경이 되는 정상 C57BL/6생쥐와 humanized mice의 대조군과 OD수치가 거의 유사하게 나타났다. 반면 세 개의 모든 실험군에서는 비록 말초혈관이나 장기에서 사람 CD19+ B 세포가 감지되지 않음에도 OD 수치 2로, 40 배 정도 희석에도 지속적으로 유지가 되었다. 비록 다른 myeloid 계열의 세포나 lymphocyte 계열의 NK 세포나 B 세 포가 감지되지는 않았지만 이렇게 수립된 humanized-mice 환경 내에서 사람 면역 세포가 형성되었다는 것을 알 수 있었다.
고 찰
우리는 C57BL/6J-Rag2/γc/ double knockout mice에 태 아의 간과 흉선 조직과 제대혈로부터 유래된 CD34+ 세포를 이식하여 lymphoid와 myeloid 계열의 세포 생성 및 분화가 일어나며 기능적인 면역 시스템을 갖추고자 하였다. 이식한 조직과 세포가 생체 안에서 repopulating되어 생쥐의 말초 혈 관, 비장 그리고 골수에서 사람 CD45+ 세포가 감지되었다.
그러나 이는 이미 보고된 다른 문헌들에 비하여 상당히 적 게 발현이 되었다. 다른 문헌들의 경우는 NOD/SCID mice, NOG (NOD/SCID IL2rγ/) mice, BALB/c-Rag2/IL2rγ/
mice에 제대혈로부터 유래된 fresh 또는 배양된 CD34+ 세포 또는 태아의 간과 흉선 조직을 이식하면 비장, 흉선, 골수 등 여러 장기에서 사람 CD45+ 세포가 적어도 40% 이상 발 현이 되었다.(10-13,15-19) 이는 생쥐 종에 따른 차이로 보여 지며 C57BL/6J를 background로 하여 humanized mice를 구축 한 문헌은 아직 보고된 바가 없다.
현재까지 보고된 문헌들은 제대혈 CD34+ 세포를 ex vivo 에서 사이토카인 조합을 다양하게 하여 분화를 최소화하면 서 stem cell로서의 성질을 최대화하는 배양 조건의 연구가 활발하게 진행 중이다. 본 연구에서는 두 가지의 사이토카 인 조합으로 배양을 하였다. 사이토카인을 첨가하여 배양 을 하게 되면 세포 수는 원하는 만큼 증가가 되나 CD34+ 세포의 발현이 약 20∼40% 정도로 낮아진다. 이는 배양하 는 동안 줄기 세포 및 전구 세포로서의 성질을 fresh CD34+ 세포에 비해 잃어버리고 granulocyte, megakaryocyte, macro- phage progenitor 세포로 더욱 분화가 되어서 나타나는 현상 이라고 볼 수 있다. 그러나 주 차별로 이식 수준은 배양 유 무에 상관없이 군 간의 차이가 유의하지 않는 것을 알 수 있다. 반면 사람의 면역 체계 형성은 G1 그룹과 G3 그룹은 전반적으로 비슷하나 G2 그룹은 이 두 그룹에 비해 낮은 수치를 갖는다. 즉, 배양을 하여 CD34+ 세포의 발현이 낮아 지더라도 이식 수준 및 면역 체계 형성이 잘 이루어지므로 세포 수 증가 측면을 고려하면 배양을 하지 않는 것보다 하는 것이 유리하며 배양 시, 사이토카인은 SCF, FL, TPO를 사용하는 것이 효과적인 것으로 생각한다.
이식 후, 6주부터 10주까지는 사람 CD45+ 세포의 수준이 비슷하다가 16주에 높은 수준을 갖는 것을 볼 수 있다. 이는 이식을 하고 생쥐 내의 환경에 세포들이 골수에 정착을 하 고 골수로부터 세포들을 성숙화 시키고 다른 기관으로 내 보내는 역할을 하는 데 그만큼의 시간이 걸리는 것이라고 볼 수 있으며 이러한 일들이 반복이 되어 16주에서 높은 수 준을 보인다고 볼 수 있다. 또한 16주에 림프구 계통의 사람 CD3+ T 세포가 감지되는 것에 반해 B 세포와 NK 세포가 감지가 되지 않았다. 이는 이식한 생쥐에 존재하는 사람 CD45+ 세포가 건강한 사람 세포(hCD45+가 약 99% 이상 존 재) 비해 현저하게 낮은 수준으로 존재하므로 T 세포에 비 해 적은 비율로 존재하는 B 세포와 NK세포가 이식한 생쥐 에서 존재하더라도 감지가 되지 않았을 가능성을 고려해 볼 수 있다.
비장 그리고 골수에서 1차 임파계장기인 골수와 2차임파 계장기인 비장에서 사람 CD45+ 세포가 발견되었다는 것은 이식한 조직과 세포가 생쥐 환경 내에 골수에 정착하여 면 역 반응에 관여하는 세포를 만들어 낼 가능성을 시사하는 것이며 비록 흉선에서 T 세포의 발달이 이루어지는지 알 수는 없었지만 비장에서 사람 CD3+ T 세포가 감지되는 것 으로 가능성을 좀 더 뒷받침할 수 있다.
In vitro에서 이러한 T 세포를 mitogenic stimulator인 PHA- L 또는 IL-2을 처리하면, 비장 내에 존재하는 사람 poly- clonal T 세포가 활발하게 증식이 일어나는 것을 볼 수 있다.
T 세포의 기능을 간접적으로 mixed lymphocyte reaction (MLR) assay, TCR repertoire expression과 cytotoxicity를 통하 여 알 수 있어 본 humanized mice에서는 MLR assay를 수행 하였으나 의미를 찾을 수 가 없었다. T 세포는 항체를 생산 하는 B 세포에 작용하여 항체를 만들도록 하며 대식 세포 에 작용하여 대식 세포를 활성화시키기도 하고, cytotoxic T lymphocyte (CTL)에 작용하여 그들을 활성화시키기도 한다.
T 세포보다 상대적으로 적게 존재하는 B 세포의 경우, 비록 사람 CD19+ 세포가 어느 기관에서도 관찰되지 않았지만, 이식 후 20주에 혈청에 Anti-human IgG-Fc 특이적 항체를 이용함으로써 그 안에 human IgG가 존재하는 것을 알 수 있었다. 이는 특정 항원으로 자극을 주었을 때, 그것을 방어 하려는 항체가 생성이 되어 면역 반응이 일어날 수 있는 가능성이 있다. 일반적으로 IgG는 1차 감염 후기와 2차 감 염 시 혈액에 존재하며 보체(complement)를 활성화 시킨다 고 알려져 있다. 또한 IgG의 반감기가 in vivo serum에서 23 일임을 고려하면, 흥미롭게도 20주라는 오랜 시간 동안 IgG 가 존재하였으며 이에 대해서는 심층적인 연구가 이루어져 야 할 것이다.
현재 저자 등은 앞서 언급된 단점을 보완하고자 NOG mice를 사용하여 fresh CD34+ 세포를 주입하여 실험을 진행 하고 있으며, 문헌에 보고된 것(15-18)과 같은 양상으로 사 람 CD45+ 세포가 발현하는 것을 확인하고 있다. 또한 사이
토카인을 사용하여 배양을 하게 되면, CD34+ 세포를 잃어 버리는 경향을 있는데, 배양을 하더라도 CD34+ 세포를 높 게 유지하기 위해 CD34+ 세포와 MSC (mesenchymal stem cell) 또는 mouse stromal cell line과 co-culture하는 방법 (20-23) 등을 시도하고 있다.
또한 humanized mice를 구축하고 그 것이 특정 항원에 반 응하는 완전한 기능적인 면역 체계를 갖추었더라도 그 것 이 지속되기 위해서는 조혈모세포의 self-renewal이 요구된 다. 최근 보고된 바에 의하면, chromatin-modifying agent인 5azaD (DNA demethylation agent)와 TSA (histone deacetylase inhibitor)를 처리함으로써 CD34+CD90+ 세포들이 약 10배 가량 증가를 하였으며, self-renewal에 관여한다고 알려진 HoxB4, Bmi1 등을 포함한 여러 유전자들의 mRNA 및 단백 질 발현이 증가가 되었다.(24-27) 이러한 chromatin-modify- ing agent들을 사용함으로써 조혈모세포의 self-renewal를 높 일 수 있을 것으로 생각한다.
본 연구에서 조직과 세포를 같이 이식했을 때, 기대했던 것과는 달리, 사람 CD45+ 세포 및 다른 계열의 세포들의 발 현이 미흡하였다. 그러나 저자들이 이전에 시도했던 세포 또는 조직만을 이식했을 때 보다 사람 CD45+ 세포의 발현 및 다른 면역 세포들의 발현이 향상 된 것을 확인할 수 있 었다. 이를 증진시키기 위하여 향후 배양 시 CD34+ 세포를 유지하는 방법 및 조혈모세포의 self-renewal을 높이는 방법 들을 이용하고 이를 NOG 또는 BALB/c-RaG1/IL2rγ/
mice 등 다른 생쥐 종에 적용함으로써 humanized mice를 구 축을 시도해 보고자 한다.
REFERENCES
1) Shultz LD, Ishikawa F, Greiner DL. Humanized mice in trans- lational biomedical research. Nat Rev Immunol 2007;7:118-23.
2) Greiner DL, Hesselton RA, Shultz LD. SCID mouse models of human stem cell engraftment. Stem Cells 1998;16:166-77.
3) Bente DA, Melkus MW, Garcia JV, Rico-Hesse R. Dengue fever in humanized NOD/SCID mice. J Virol 2005;79:13797-9.
4) Cravens PD, Melkus MW, Padgett-Thomas A, Islas-Ohlmayer M, Del P Martin M, Garcia JV. Development and activation of human dendritic cells in vivo in a xenograft model of hu- man hematopoiesis. Stem Cells 2005;23:264-78.
5) Palucka AK, Gatlin J, Blanck JP, Melkus MW, Clayton S, Ueno H, et al. Human dendritic cell subsets in NOD/SCID mice engrafted with CD34+ hematopoietic progenitors. Blood 2003;102:3302-10.
6) Islas-Ohlmayer M, Padgett-Thomas A, Dolmiati-Saad R, Melkus MW, Cravens PD, Martin Mdel P, et al. Experimental infection of NOD/SCID mice reconstituted with human CD34+ cells with Epstein-Barr virus. J Virol 2004;78:13891- 900.
7) Aldrovandi GM, Feuer G, Gao L, Jamieson B, Kristeva M,
Chen IS, et al. The SCID-hu mouse as a model for HIV-1 infection. Nature 1993;363:732-6.
8) McCune JM, Namikawa R, Kaneshima H, Shultz LD, Lieberman M, Weissman IL. The SCID-hu mouse: murine model for the analysis of human hematolymphoid differ- entiation and function. Science 1988;8241:1632-9.
9) Vandekerckhove BA, Krowka JF, McCune JM, de Vries JE, Spits H, Roncarolo MG. Clonal analysis of the peripheral T cell compartment of the SCID-hu mouse. J Immunol 1991;
146:4173-9.
10) Melkus MW, Estes JD, Padgett-Thomas A, Gatlin J, Denton PW, Othieno FA, et al. Humanized mice mount specific adap- tive and innate immune responses to EBV and TSST-1. Nat Med 2006;12:1316-22.
11) Piacibello W, Sanavio F, Severino A, Dane A, Gammaitoni L, Fagioli F, et al. Engraftment in nonobese diabetic severe combined immunodeficient mice of human CD34+ cord blood cells after ex vivo expansion: evidence for the amplification and self-renewal of repopulating stem cells. Blood 1999;93:
3736-49.
12) Ueda T, Tsuji K, Yoshino H, Ebihara Y, Yagasaki H, Hisakawa H, et al. Expansion of human NOD/SCID-repopulating cells by stem cell factor, Flk2/Flt3 ligand, thrombopoietin, IL-6, and soluble IL-6 receptor. J Clin Invest 2000;105:1013-21.
13) Suzuki T, Yokoyama Y, Kumano K, Takanashi M, Kozuma S, Takato T, et al. Highly efficient ex vivo expansion of hu- man hematopoietic stem cells using Delta1-Fc chimeric protein. Stem Cells 2006;24:2456-65.
14) Gaffen SL, Liu KD. Overview of interleukin-2 function, pro- duction and clinical applications. Cytokine 2004;28:109-23.
15) Yahata T, Ando K, Nakamura Y, Ueyama Y, Shimamura K, Tamaoki N, et al. Functional human T lymphocyte develop- ment from cord blood CD34+ cells in nonobese diabetic/
Shi-scid, IL-2 receptor gamma null mice. J Immunol 2002;
169:204-9.
16) Hiramatsu H, Nishikomori R, Heike T, Ito M, Kobayashi K, Katamura K, et al. Complete reconstitution of human lympho- cytes from cord blood CD34+ cells using the NOD/SCID/gam- mac null mice model. Blood 2003;102:873-80.
17) Ishikawa F, Yasukawa M, Lyons B, Yoshida S, Miyamoto T, Yoshimoto G, et al. Development of functional human blood
and immune systems in NOD/SCID/IL2 receptor {gamma}
chain (null) mice. Blood 2005;106:1565-73.
18) Watanabe S, Terashima K, Ohta S, Horibata S, Yajima M, Shiozawa Y, et al. Hematopoietic stem cell-engrafted NOD/
SCID/IL2Rgamma null mice develop human lymphoid sys- tems and induce long-lasting HIV-1 infection with specific hu- moral immune responses. Blood 2007;109:212-8.
19) Gorantla S, Sneller H, Walters L, Sharp JG, Pirruccello SJ, West JT, et al. Human immunodeficiency virus type 1 pathobi- ology studied in humanized BALB/c-RaG1-/-gammac-/- mice.
J Virol 2007;81:2700-12.
20) Noort WA, Kruisselbrink AB, in’t Anker PS, Kruger M, van Bezooijen RL, de Paus RA, et al. Mesenchymal stem cells promote engraftment of human umbilical cord blood-derived CD34+ cells in NOD/SCID mice. Exp Hematol 2002;30:870-8.
21) Zhang Y, Chai C, Jiang XS, Teoh SH, Leong KW. Co-culture of umbilical cord blood CD34+ cells with human mesen- chymal stem cells. Tissue Eng 2006;12:2161-70.
22) Kawada H, Ando K, Tsuji T, Shimakura Y, Nakamura Y, Chargui J, et al. Rapid ex vivo expansion of human umbilical cord hematopoietic progenitors using a novel culture system.
Exp Hematol 1999;27:904-15.
23) Nolta JA, Thiemann FT, Arakawa-Hoyt J, Dao MA, Barsky LW, Moore KA, et al. The AFT024 stromal cell line supports long-term ex vivo maintenance of engrafting multipotent hu- man hematopoietic progenitors. Leukemia 2002;16:352-61.
24) Tadokoro Y, Ema H, Okano M, Li E, Nakauchi H. De novo DNA methyltransferase is essential for self-renewal, but not for differentiation, in hematopoietic stem cells. J Exp Med 2007;204:715-22.
25) Araki H, Mahmud N, Milhem M, Nunez R, Xu M, Beam CA, et al. Expansion of human umbilical cord blood SCID-re- populating cells using chromatin-modifying agents. Exp Hematol 2006;34:140-9.
26) Araki H, Yoshinaga K, Boccuni P, Zhao Y, Hoffman R, et al. Chromatin-modifying agents permit human hematopoietic stem cells to undergo multiple cell divisions while retaining their repopulating potential. Blood 2007;109:3570-8.
27) Akala OO, Clarke MF. Hematopoietic stem cell self-renewal.
Curr Opin Genet Dev 2006;16:496-501.