www.earticle.net
배아줄기 세포의 치료능력수프리야 시와니1 · 왕명현1 · 나레시 쿠마 싱2*
강원대학교 의생명공학과1, 강원대학교 동물생명공학과2
Therapeutic Potential of Embryonic Stem Cells-An Overview
Supriya Shiwani
1, Myeong Hyeon Wang1 and Naresh Kumar Singh2*1
Dept. of Medical Biotechnology, College of Biomedical Sciences,
2
Dept. of Animal Biotechnology, College of Animal Life Sciences, Kangwon National University
ABSTRACT
1)Embryonic stem (ES) cells are pluripotent cells derived from the inner cell mass cells of blastocyst with the potential to maintain an undifferentiated state indefinitely. Fully characterized ES cell lines express typical stem cell markers, possess high levels of telomerase activity, show normal karyotype and have the potential to differentiate into numerous cell types under in vitro and in vivo conditions.
Therefore, ES cells are potentially valuable for the development of cell transplantation therapies for the treatment of various diseases in animals as well as in humans. However, important problems associated with ES cells from in vitro fertilized blastocysts particularly from humans must be resolved before taking up its therapeutic applications. Current techniques for directed differentiation into somatic cell populations remain inefficient and yield heterogeneous cell populations. This review therefore focuses on ES cells with respect to in vitro propagation and differentiation in basic cell and developmental biology for successful use of these cells in therapeutics.
(Key Words: Embyronic stem cells, Pluripotency, Therapeutic potential)
Introduction
During embryonic development and tissue regeneration stem cells have been observed and considered as natural physiological units of building blocks, in particular, the embryonic stem cells. Embryonic stem (ES) cells possess a nearly unlimited self–renewal capacity and developmental potential to differentiate into virtually any cell type of an organism. Initially, with the establishment of mouse ES cells as permanent cell lines major advances in cell and developmental biology took place and was considered a breakthrough in biomedical sciences. ES cells, which are
derived from the inner mass of blastocyts or primordial germ cells, can be maintained continuously in an undifferentiated pluripotent state. They have the potential to contribute to a variety of differentiated cell lineages in the fetus including germ cells (Evans and Kaufman, 1981;
Martin, 1981). Targeted mutations in ES cells in the past have been introduced into the mouse germ lines to elucidate gene function invivo and create mouse models to study human genetic diseases and abnormalities (Smithies et al., 1985; Cawpecchi,1989). The availability of embryonic stem celll ines in mammalian species has greatly advanced the field of biological research by enhancing our ability to
* Corresponding Author: N. K. Singh, Dept. of Animal Biotechnology, Kangwon National University, Chuncheon 200-701, Korea. Tel:
+82-33-250-8637, E-mail: [email protected]
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM
www.earticle.net
manipulate the genome and by providing model systems to examine cellular differentiation.
Major breakthroughs for cell and developmental biology that has changed our views on life processes are;
1) the establishment of embryonic stem cell lines derived from mouse (Evans and Kaufman, 1981; Martin, 1981) and human embryos (Thomson et al., 1998), 2) creation of genetic mouse models of disease through homologous recombination in ES cells (Thomas and Capecchi, 1987), 3) The reprogramming of somatic cells after nuclear transfer into enucleated eggs (Wilmut et al., 1997), and 4) demonstration of germ line development of ES cells in vitro (Gassman et al., 1995; Hubner et al., 1993;
Toyookaetal., 2003). Because of these break throughs, cell therapies base do nan un limited, renewable source of cells have become an attractive concept of regenerative medicine.
Furthermore, the fertilized egg has the ability to generate an entire organism as it has been observed and found that ES cell has self renewal capacity that depends on a stoichiometric balance among various signaling molecules present in them (Hosler et al., 1993). Any minute imbalance in any one of these known and unknown signaling molecules could cause ES cell identity to be lost due to their innate pluripotent potentials. Few such molecular markers that defines pluripotentiality include Rex-1 (Hosler et al., 1993; Rogers et al., 1991), Sox-2 (Avilion et al., 2003), UTF1 (Okuda et al., 1998), Genesis (Sutton et al., 1996), GBX2 (Chapman et al., 1997), Pem (Fan et al., 1999) and L17 (Rodda et al., 2001).
Embryonic stem cell based therapies
Three properties that make ES cells relative to adult stem cells very attractive for replacement therapies (Hochedlinger and Jaenisch, 1999) are;
1. Human ES cells can be grown indefinitely in culture 2. ES cells can be genetically manipulated and loss of
functional genes (eg. CTFR) can theoretically be repaired by the introduction of transgene into ES cells either by random transgenesis or through gene
targeting.
3. Numerous differentiation protocols have already been established that permit the generation of almost any cell type either through the use of established culture conditions or when coupled with genetic manipulation.
Embryonic stem cells for cardiac repair
Embryonic stem (ES) cells have taken centre stage in the scientific debate due to their intrinsic capacity to transdifferentiate into endoderm-derived cardiomyocytes (Klug et al., 1996; Kehat et al., 2001; Mummery et al., 2003) and its characterization by well-defined protocols during in vitro expansion (scalable for potential human application) (Zandstraetal., 2003). The successful generation of expandable, fully differentiated force- generating cardiomyocytes, capable of excitation–
contraction coupling, (Kehat et al., 2001) renders them a particularly attractive cell type for post-infarction repair.
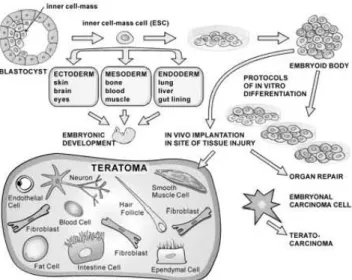
However, irrespective of ethical considerations, major technical hurdles continue to concern researchers and doctors that includes the risk of uncontrolled neoplastic transformation, potential development of teratomas, immune responses and disruptive growth within the host tissue primed with paracrine signals from ES cell-derived cytokines (Nussbaum et al., 2007) (Fig. 1).
Leor and coworkers (2007) performed many experiments with hES cells on cardiac tissues and demonstrated that proper cardiac specification is must before transplanting hES cell in a hostile atmosphere such as ischaemic or acellular environment. Undifferentiated human ES cells injected into the myocardial wall do not transdifferentiate into cardiac or endothelial lineage as observed by Foley and Mercola (2004) and this finding was in corroboration with the findings of Leor et al.
(2007) and which could be concluded as that the ES cells specification is limited and transplantation outside the embryonic environment disrupts their normal differentiation programme (Rohwedel et al., 1996).
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM
www.earticle.net
Fig. 1. Embryonic stem cells. During development, embryonic stem cells (ESCs) give rise to the ectoderm, mesoderm, and endoderm. ESCs are obtained from the inner cell mass of the blastocyst and in vitro form embryoid bodies (EBs) that contain undifferentiated (yellow), ectodermal (green), mesodermal (blue), and endodermal (red) cells.Implantation of EBs in vivo can repair damaged organs and generate tumors.
Recently, Behfar and colleagues identified a key role for tumor necrosis factor α in the release of cardio- inducive signals from the intact murine endoderm (Behfar et al., 2007), and that resulted in the generation of murine cardiopoietic cells. The scientific challenge in the years to come will be to investigate effective and safe cardiopoietic programming of human ES cells feasible for cardiac repair (Fig. 1). Cardiac restricted promoters have been used to select cardiomyocytes from differentiating ES cells (Fijnvandraat et al., 2003; Klug et al., 1996; Meyer et al., 2000). Min et al. (2002) reported improved left ventricular function in post-infarcted rats after transplantation of beating cells derived from ES cells. The engrafted cardiomyoctyes expressed sarcomeric alpha-actin, alpha-myosin heavy chain and troponin I and were rod-shaped with typical striation suggesting differentiation into mature cardiomyocytes. ES cell-derived cardiomyocytes expresses myofilament proteins and were able to form normal coupling units with endogenous
cardiomyocytes. However, many questions obviously remain, most notably about the evidence or not for immune tolerance of ES cells or their progeny in the heart (Nussbaum et al., 2007) and about the long-term follow- up after human ES cell transfer.
ES cells for invitro formation of vascular structures
Blood vessel formation is a complex process that results in the elaboration of a vasculature with a distinct pattern (reviews Wagner, 1980; Noden, 1991). Blood vessels are one of the first organ systems to develop during embryogenesis, and the vasculature continues to develop and expand into the postnatal period (Stewart and Wiley, 1981; Bar, 1983; Coffin and Poole, 1988;
Pardanaud et al., 1989). A key control point in blood vessel formation is the response of the endothelial cell to developmental signals, and this response has two components. Vascular development by in situ primary differentiation of endothelial cells from an undifferentiated precursor cell is called vasculogenesis, whereas the subsequent "sprouting" and organization of endothelial cells into blood vessels is called angiogenesis (Houser et al., 1961; Risau and Lemmon, 1988). Angiogenes is involves endothelial cell migration, division, and the establishment of junctional connections. Vasculogenes is occurs when aputative precursor cell differentiatest of or manangioblast and a stem cell for the hematopoietic lineages (Sabin, 1920; Noden, 1991).
In vitro differentiation of ES cells to form blood island-containing CEBs as a model for the study of embryonic blood vessel formation revealed that the blood islands of CEBs are lined by endothelial cells. Moreover, it was further revealed that the blood islands of the embryoid bodies in suspension culture mature to form connections and channels that resemble blood vessels.
Thus, signals for both the primary differentiation of endothelial cells from pluripotent stem cells and for some of the cellular processes involved in subsequent vascular development are intrinsic to the ES cell culture system.
During differentiation, transcripts characteristic for
醫뚫쫓뚫옳앓 융
뒀:쏟cι
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM
www.earticle.net
endothelial cells were detected that includes GATA-2, PECAM-1, Flk-1 and VE-cadherin. PECAM-1 antibodies were used to isolate endothelial cells from the 13 day EBs after enzymatic dissociation and the isolated PECAM-1 cells were seeded onto highly porous PLGA biodegradable polymer scaffolds (Putnam and Mooney, 1996) and sponges were implanted subcutaneously into SCID mice (Leon-Queent et al., 2004). After development in SCID mice as well as after in vitro differentiation in matrigel, micro vessels developed from human ES cells.
ES cells for neuropathies
Embryonic stem cells (ESCs) have the potential to become a limitless source of tissue for cell therapies and drug testing. Specifically, efficient generation of neuronal cells from human ESCs (hESCs) has major impact in transplantation therapies to replace aged or diseased neural tissue. In the recent past, several studies based on transdifferentiation of ES cells have improved our knowledge concerning the specific signals directing the differentiation of ESCs to neural progeny. Large scale differentiation of hESCs in a bioreactor as a way of producing neuronal cells in quantities has been found to be suitable for clinical therapies. The integrative nuclear fibroblast growth factor (FGF) signaling (INFS) has been shown to mediate the activation of genes thereby coaxing the differentiation of umbilical cord blood cells and human neuronal precursors to neuronal cells. Treatment of mouse ESCs with agents such as retinoic acid (RA) and cyclic adenosine monophosphate (cAMP) has been found to activate INFS, in part by nuclear translocation of an FGF receptor (Brustle et al., 1999). Immuno-positivity for glial fibrillary acidic protein (GFAP) has revealed an increased propensity of mouse ESCs for astrocyte differentiation upon stimulation with cAMP. In contrast, RA prompted the differentiation towards neurons as shown by enhanced expression of tyrosine hydroxylase and β3-tubulin. Methods have been developed for the directed differentiation of ESCs to neuronal cells with high efficiency and in large quantities to be utilized in
clinical therapies. Cells formed neurosphere-like aggregates in the bioreactor and in a serum-free medium. Such transition to neuroepithelial cells and then radial glial cells has been noted recently. Finally, neuron-like cells emerged upon plating of the neurospheres and the differentiated cell has displayed increased gene/protein levels of Pax6, Nestin, β3-tubulin, MAP2 and GABA compared to undifferentiated hESCs (Fig. 2). This scalable system has the ability to produce mass quantities of neurospheres. In conjunction with an INFS-based method for directing the differentiation of these cells further to neuronal cells may facilitate the production of cellular material suitable for therapies against Parkinson and Alzheimer diseases (Brustle et al., 1999). Successful generation of apparently normal neural cell types from in vitro differentiated EScells has naturally led to intense interest in their potential use to repair or limit the damage associated with in farctor neuro degenerative diseases. Brustle et al. (1999) first demonstrated that EScells–derived neural cells could survive, respond to environmental signals and exhibit aspect so fregion specific differentiation when introduced into developing mouse brain. They also showed that following in vitro generation of precursors of oligo-dendrocytes and astrocytes, these transplanted cells interact with hostneuronsand myelinate the axons in brain and spinal cords in a rat model of human dysmyelinating Pleizaeus-Meizhacherdisease (Brustle et al., 1999). The resulting remyelination of axons led to are covery of the pathological phenotype in the animals.
ES cells for the treatment of diabetes
Embryonic stem cells (ES) can be differentiated into insulin-producing cells by manipulating culture conditions. In-vitro differentiation of mouse ES can generate embryoid bodies, which, after selection for nestin expressing ES could be stimulated to differentiate towards a insulin producing cell like phenotype (Lumelsky et al., 2001). The addition of phosphor-inositide kinase inhibitors have also been shown to promote larger numbers of ESC
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM
www.earticle.net
towards functional cells (Hori et al., 2002). Variations in ES-culture conditions generate cells with properties insulin producing cells (Kania et al., 2002; Kahan et al., 2003; Stoffel et al., 2004). With manipulation of culture conditions and use of pax4 or pdx-1, transcription factors associated with cell lineage (Blyszczuk et al., 2003;
Miyazaki et al., 2003) could yield promising results.
Recently, some doubt has been casted on whether ESC differentiation protocols truly yield cells that produce insulin, or cells that merely absorb insulin from the medium (Rajagopal et al., 2003). Later, it was concluded that these differentiated cells must actively synthesize and secrete insulin rather than insulin being detected.
Moreover, functioning molecular components of regulated secretion of insulin and insulin-containing vesicles were further taken up as essential additional features that indicate a cell phenotype. Transplantation of ES derived insulin-producing cells has reversed diabetes in rodents indicating that these cells do synthesize and release insulin (Assady et al., 2001). Regulated expression of introduced transcription factors that can be turned on during in-vitro differentiation of ES might be more successful in secreting insulin. ES, genetically selected for insulin expression and injected into diabetic rats, improve glucose control more efficiently (Soria et al., 2001) and human ESC also produces insulin under different culture conditions (Assady et al., 2001; Segev et al., 2003).
Techniques that do not require murine feeder cells have been developed, allowing for single species propagation of ES and avoiding possible zoonotic infection of cells intended for clinical use (Moritoh et al., 2003). Problems in control of differentiation and teratoma formation from ES-derived insulin-producing cells still remain to be overcome (Sipione et al., 2004). Existing ES lines are not assumed to be identical or ideal for generating islets or cells, and hence additional ES lines needs to be generated continuously (Cowan et al., 2004). Mouse embryonic stem cells differentiate into functional islet like cells that are able to rescue experimentally induced diabetes in mouse models (Blyszczuk et al., 2003; Hori et al., 2002). Efficient selection methods with pancreatic lineage specific
promoters will be necessary to overcome current limitations, such as tumor formation of grafts and low insulin levels. Ethical concerns about the use of ESC need to be addressed and resolved in the face of this powerful technology.
Therapeutic cloning
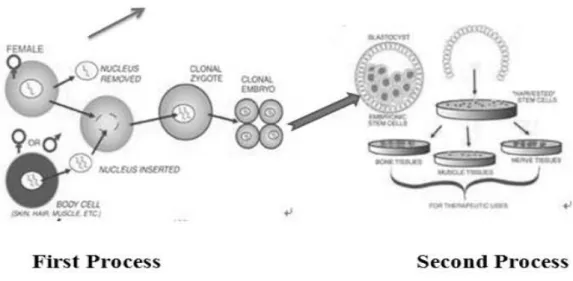
Cloning is defined as the production of a set of individuals with same genotype. This occurs naturally by asexual reproduction in hydra, sea anemone, planarians, and annelids (Yanagimachi, 2002), but cloning is also possible with mammalian somatic cells as demonstrated by birth of “Dolly” the sheep in 1997 (Wilmut et al., 1997). Therapeutic cloning utilizes nuclear transfer techniques to produce pluripotent ES cells with genome of the nucleus of origin. Specifically the nucleus of an adult donor cell is introduced into an enucleated donor oocyte to generate a cloned embryo. The somatic cell nucleus at a low frequency and depending on the donor cell type may regain its pluripotentiality to initiate the earliest stages of embryonic development. ES cells are amenable to genetic manipulation and when combined with therapeutic cloning, ES cell derivatives offer the potential for both genes and cell based therapies (Fig. 2).
Therapeutic cloning for ESC therapy is another stem cells research area which is gaining momentum. It involves transfer of somatic nuclei from non-reproductive cells of patient’s body to enucleated donated eggs (oocytes) and production of embryos from these fused cells. The embryos produced thus are used for collecting pluripotent embryonic stem cells, which can differentiate into any cell type identical to that of a part of the body of the patient. Cells and tissues derived from such embryonic stem cells for the purpose of transplantation will not be rejected by the patient’s body, since the nucleus was derived from the patient. This technology therefore has far reaching implications in using ESCs for therapeutic purposes.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM
www.earticle.net
Fig. 2. Schematic presentation shows that the clonal embryo resulting from the first process from blastocyst source and in the second process the cultivation of stem cells and resultant specialized cells have been shown for therapeutic use.
Production of high yielding animals
The introduction of specific genes into the genome of livestock and their stable transmission into the genome of the resultant offspring has been regarded as major technical advances in animal production. Transgenic technology is a rapid method to introduce new genes into cattle, swine, sheep and goats without cross breeding (Pursel and Rexroad, 1993). Methods that have been used to produce transgenic animals include:
1. DNA transfer by retroviruses
2. Microinjection of genes into pronuclei of fertilized ova 3. Injection of embryonic stem cells exposed to foreign
DNA into the cavity of blastocysts
4. Sperm mediated exogenous DNA transfer during the in vitro fertilization
5. Liposome mediated DNA transfer into cells and embryos
6. Electroporation of foreign DNA into sperm, ova or embryos
7. Biolistics
8. Nuclear transfer with somatic or embryonic cells (Wheeler and Walters, 2001)
Production of transgenic animals has numerous practical applications which include increased growth rate, improved feed utilization, improved carcass composition, increased disease resistance, enhanced reproductive performance, increased prolificacy and altered cell and tissue characteristics for biomedical research (Wheeler and Choi, 1997) and manufacturing. Alteration in composition of milk through transgenic technology led to improvement in production of Keratin proteins and / or growth factors that are deficient in milk (Bremel et al., 1989).
Improvement of nutrients or therapeutic value of milk may have a profound impact on survival and growth of newborns in both humans and animals. In swine, 44% of the growth rate of the developing piglets can be attributed to yield and composition of sow milk (Lewis et al., 1978).
How do transgenic animals contribute to human welfare?
The benefits of transgenic animals to human welfare have been grouped into three categories and that are as follows;
1. Agriculture applications
. / ’
“
Fi rst P ..
oce~‘s‘ ... ‘-
; p
Second Process
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM
www.earticle.net
· Breeding: Farmers have always used selective breeding to produce animals that exhibit desired traits (e.g., increased milk production, high growth rate).
Traditional breeding is at time-consuming, difficult task. When technology using molecular biology was developed, it became possible to develop traits in animals in a shorter time and with more precision.
Inaddition, it offers the farmer an easy way to increase yields.
· Quality: Transgenic cows could produce more milk or milk with less lactose or cholesterol.
· Disease resistance: Scientists are attempting to produce disease-resistant animals, such as influenza-resistant pigs, but a very limited number of genes are currently known to be responsible for resistance to diseases in farm animals.
2. Medical Applications
· Xenotransplantation: Patients die every year for lack of a replacement of heart, liver, or kidney. Transgenic pigs may provide the transplant organs needed to alleviate this shortfall. Currently, xenotransplantation is hampered by a pig protein that can cause donor rejection but research is underway to remove the pig protein and replace it with a human protein.
· Nutritional supplements and pharmaceuticals: Products such as insulin, growth hormone, and blood anti- clotting factors may soon be or have already been obtained from the milk of transgenic cows, sheep, or goats. Research is also underway to manufacture milk through transgenesis for treatment of debilitating diseases such as phenylketonuria, hereditary emphysema, and cystic fibrosis.
· Human gene therapy: Human gene therapy involves adding a normal copy of a gene (transgene) to the genome of a person carrying defective copies of the gene. The potential for treatments for the 5,000 named genetic diseases is huge and transgenic animals could play a role.
3. Industrial Applications
In 2001, two scientists at Nexia Biotechnologies in Canada spliced spider genes into the cells of lactating
goats. The goats began to manufacture silk along with their milk and secrete tiny silk strands from their body.
By extracting polymer strands from the milk and weaving them into thread, the scientists can create a light, tough, flexible material that could be used in such applications as military uniforms, medical micro-sutures, and tennis racket strings.
Conclusion
Stem cell therapy is certainly a promising area for research. Stem cells have the ability to give rise to many specialized cells in an organism. The broadest potential application is the generation of cells and tissues that could be used to repair or replace damaged organs. If scientists can learn how to control stem cell conversion into new, functionally mature cells, doctors might be able to cure many diseases for which therapy is currently inadequate.
References
1. Assady, S., Maor, G., Amit, M., Itskovitz-Elder, J., Skorecki, K. L. and Tzukerman M. 2001. Insulin production by human embryonic stem cells. Diabetes.
50:1691–97.
2. Avilion, A. A., Nicoles, S. K., Pevny, L. H., Peuz, L., Vivian, N. and Lovell-badge, R. 2003. Multipotent cell lineages in early mouse development depend on SOX-2 function. Genes Dev. 17:126-140.
3. Bar, T. 1983. Patterns of vascularization in the developing cerebral cortex. In Development of the vascular system, pp. 20-36. Ciba Foundation Symp. 100. London: Pitman Books.
4. Behfar, A., Perez-Terzic, C., Faustino, R. S. 2007.
Cardiopoietic programming of embryonic stem cells for tumor-free heart repair. J. Exp. Med. 204:405–20.
5. Blyszczuk, P., Czyz, J., Kania, G., Wagnes, M., Roll, U., St Onge, L, Wobus, A. M. 2003. Expression of Pax-4 in embryonic stem cells promotes differentiation of
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM
www.earticle.net
nestin-positive progenitor and insulin producing cells.
Pron Natl Acad. Sci. USA. 100:998-1003.
6. Bremel, R. D., Yom, H.C. and Bleck, G.T. 1989.
Alteration of milk composition using molecular genetics.
J. Dairy Sci. 72:2826–2833.
7. Brustle, O., Jones, K. N., Learish, R. D., Karram, K., Choudhary, K., Wiestler, O. D., Duncan, I. D. and McKay, R. D. 1999. Embryonic stem cells-derived glial precursor: a source of myelinating transplants. Sci.
285:754-756.
8. Brustle, O., Spiro, A. C., Karram, K., Choudhary, K., Okabe, S., McKay, R. D. 1997. In vitro generated neural precursors participate in mammalian brain development.
Proc. Natl. Acad. Sci. USA. 94:14809-14814.
9. Cawpecchi, M R. 1989. Altering the genome by homologous recombination. Sci. 244:1288-1292.
10. Chapman, G., Remiszewski, J. L., Webb, G. C., Schulz, T.
C., Bottema, C. D. and Rathjen, P. D. 1997. The mouse homeobox gene, Gbx 2: genomic organization and expression in pluripotent cells in vitro and in vivo. Genomics. 46:223-233.
11. Coffin, J. D. and Poole, T. J. 1988. Embryonic vasculature development: immunohistochemical identification of the origin and subsequent morphogenesis of the major vessel primordia in quail embryos. Development 102:735-748.
12. Cowan, C.A., Klimanskaya, I. and McMahon, J. 2004.
Derivation of embryonic stem-cell lines from human blastocysts. N. Engl. J. Med. 350:1353–56.
13. Evans, M. I. and Kaufman, M. H. 1981. Establishment in culture of pluripotent cells from mouse embryos.
Nat. 1292:154-156.
14. Fan, Y., Melhem, M. F. and Chaillet, J. R. 1991. Forced expression of the homeobox-containing gene Pem blocks differentiation of embryonic stem cells. Dev.
Biol. 210:481-496.
15. Fijnvandraat, A. C., vanGinnkken, A. C., Schumacher, C. A., Boheler, K. R., Lelanne Dupez, R. H., Christoffels, V. M. and Moorman, A. F. 2003.
Cardiomyocytes purified from differentiated embryonic stem cells exhibit characteristics of early chamber myocardium. J. Mol. Cell Cardiol. 35:
1461-1472.
16. Foley, A. and Mercola, M. 2004. Heart induction:
embryology to cardiomyocyte regeneration. Trends Cardiovasc. Med. 14:121–5.
17. Gassmann, M., Donoho, G. and Berg, P. 1995.
Maintenance of an extra-chromosomal plasmid vector in mouse embryonic stem cells. Proc. Natl. Acad. Sci.
USA. 92:1291-1296.
18. Hochedlinger, K. and Jaenisch, R. 1999. Nuclear transplantation, embryonic stem cells and the potential for cell therapy. N. Eng. J. Med. 349:275-286.
19. Hori, Y., Rulifson, I. C., Tsai, B. C, Heit, J. J., Cahoy, J. D. and Kim, S. K. 2002. Growth inhibitors promote differentiation of insulin-producing tissue from embryonic stem cells. Proc. Natl. Acad. Sci. 99:16105–
10.
20. Hosler, B. A., Rogers,, H. B., Kozak, C. A., Gudas, L.
J. 1993. An octamer motif contributes to the expression of the retinoic acid regulated zinc finger gene Rex-1 (Zfp-42) in F9 teratocarcinoma. Mol. Cell Biol. 13:2919-2928.
21. Houser, J. W., Ackerman, G. A. and Knouff, R. A.
1961. Vasculogenesis and erythropoiesis in the living yolk sac of the chick embryo. Anat. Rec. 140:29-43.
22. Hubner, K., Fuhrmann, G., Christenson, L. K., Kehler, J., Reinbold, R., De La, F. R., Wood, J., Strauss, III. J.
F., Boiani, M. and Scholer, H. R. 2003. Derivation of oocytes from mouse embryonic stem cells. Sci.
300:1251-1256.
23. Kahan, B. W., Jacobson, L. M. and Hullett, D. A. 2003.
Pancreatic precursors and differentiated islet cell types from murine embryonic stem cells. Diabetes. 52:2016–
24.
24. Kania, G., Blyszczuk, P., Czyz, J., Navarrete-Santos, A.
and Wobus, A. M. 2003. Differentiation of mouse embryonic stem cells into pancreatic and hepatic cells.
Meth. Enzymol. 365:287–303.
25. Kehat, I., Kenyagin-Karsenti, D. and Snir, M. 2001.
Human embryonic stem cells can differentiate into myocytes with structural and functional properties of cardiomyocytes. J. Clin. Invest. 108:407–14.
26. Klug, M. G., Soonpaa, M. H., Koh, G. Y. and Field, L.
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM
www.earticle.net
J. 1996. Genetically selected cardiomyocytes from differentiating embryonic stem cells from stable intracardiac grafts. J. Clin. Invest. 98:21-224.
27. Klug, M. G., Soonpaa, M. H., Koh, G. Y., Field, L. J.
1996. Genetically selected cardiomyocytes from differentiating embryonic stem cells from stable intracardiac grafts. J. Clin. Invest. 98:21-224.
28. Leon-Queent, T., Jones, J., Skoudy, A., Burcin, M. and Soria, B. 2004. In vitro directed differentiation of mouse embryonic stem cells into insulin producing cells. Diabetologia. 47:1442-1451.
29. Leor, J., Gerecht, S. and Cohen, S. 2007. Human embryonic stem cell transplantation to repair the infarcted myocardium. Heart. 93:1278–84.
30. Lewis, A. J., Speer, V. C. and Haught, D. G. 1978.
Relationship between yield and composition of sows milk and weight gains of nursing pigs. J. Anim. Sci.
47:634–638.
31. Lumelsky, N., Blondel, O., Laeng, P., Vlesco, I., Ravion, R. and McKay, R. 2001. Differentiaion of embryonic stem cells to insulin-secreting structures similar to pancreatic islets. Sci. 292:1398–94.
32. Martin, G R. 1981. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma cells. Proc. Natl. Acad.
Sci. USA. 78:7634-7638.
33. Min, J. Y., Yang, Y., Converso, K. L., Liu, L., Huang, Q., Morgan, J. P. and Xiao, Y. F. 2002. Tranplantation of embryonic stem cells improved cardiac function in post infracted rats. J. Appl. Physiol. 92:288-296.
34. Miyazaki, S., Yamato, E. and Miyazaki, J. 2004 Regulated expression of pdx-1 promotes in vitro differentiation of insulin-producing cells from embryonic stem cells. Diabetes. 53:1030–37.
35. Moritoh, Y., Yamato, E., Yasui, Y., Miyazaki, S. and Miyazaki, J. 2003. Analysis of insulin-producing cells during in vitro differentiation from feeder-free embryonic stem cells. Diabetes. 52:1163–68.
36. Mummery, C., Ward-van Oostwaard, D. and Doevendans, P. 2003. Differentiation of human embryonic stem cells to cardiomyocytes: role of coculture with visceral endoderm-like cells. Circulation. 107:2733–40.
37. Noden, D. M. 1991. Development of craniofacial blood vessels. In the Development of the Vascular System, (ed. R. N. Feinberg, G. K. Sherer, and R. Auerbach), pp. 1-24.
38. Nussbaum, J., Minami, E. and Laflamme, M.A. 2007.
Transplantation of undifferentiated murine embryonic stem cells in the heart: teratoma formation and immune response. FASEB J. 21:1345–57.
40. Pardanaud, L., Yassine, F. and Dieterien-Lievre, F. 1989.
Relationship between vasculogenesis, angiogenesis, and haemopoiesis during avian ontogeny. Development.
105:473-485.
41. Pursel, V. G. and Rexroad, C. E. 1993. Status of research with transgenic farm animals. J. Anim. Sci.
71(Suppl. 3):10–19.
42. Putnam, A. J. and Mooney, D. J. 1996. Tissue engineering using synthetic extracellular matrices. Nat.
Med. 2:824-826.
43. Rajagopal, J., Anderson, W. J., Kume, S., Martinez, O.
I. and Melton, D. A. 2003. Insulin staining of ES cell progeny from insulin uptake. Sci. 299:363.
44. Risau, W. and Lemmon, V. 1988. Changes in the vascular extracellular matrix during embryonic vasculogenesis and angiogenesis. Dev. Biol. 125:441-450.
45. Rodda S, Sharma S, Scherer M, Chapman G, Rathjen P. CRTR-1, a develpomentally regulated transcriptional repressor related to the CP2 family of transcription factors. 2001. J. Biol. Chem. 276:3324-3332.
46. Rogers, M. B., Hosler, B. A. and Gudas, L. J. 1991.
Specific expression of a retinoic caid-regulated, zinc-finger gene, Rex-1 in preimplantation embryos, trophoblast and spermatocytes. Development. 113:815-824.
47. Sabin, F. R. 1920. Studies on the origin of the blood vessels and of red blood corpuscles as seen in the riving blastoderm of chick during the second day of incubation. Contrib. Embryol. Carnegie Inst. 9:215-262.
48. Segev, H., Fishman, B., Ziskind, A., Shulman, M. and Itskovitz-Eldor, J. 2004. Differentiation of human embryonic stem cells into insulin-producing clusters.
Stem Cells. 22:265–74.
49. Sipione, S., Eshpeter, A., Lyon, J. G., Korbutt, G. S.
and Bleackley, R. C. 2004. Insulin expressing cells from
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM
www.earticle.net
differentiated embryonic stem cells are not beta cells.
Diabetologia. 47:499–507.
50. Smithies, O. 1985. Insertion of DNA sequences into the human chromosomal beta-globin locus by homologous recombination. Nat. 317:230-234.
51. Soria, B. and Skoudy, A. and Martin F. 2001. From stem cells to beta cells. Diabetologia. 44:407–15.
52. Stewart, P. A. and Wiley, M. J. 1981. Developing nervous tissue induces formation of blood-brain barrier characteristics in invading endothelial cells: a study using quail-chick transplantation chimeras. Dev. Biol.
84:183-192.
53. Stoffel, M., Vallier, L. and Pedersen, R. A. 2004.
Navigating the pathway from embryonic stem cells to beta cells. Semin Cell Dev. Biol. 15:327–36.
54. Sutton, J., Costa R., Klug, M., Field. L., Xu, D., Largaes pada, D. A., Fletcher, C. F., Jentkins, N. A., Copeland, N. G., Klemz, M., Hromas, R. 1996.
Genesis, a winged helix transcriptional repressor with expression restricted to embryonic stem cells. J. Biol.
Chem. 271:23126-23133.
55. Thomas, K. R. and Capecchi, M. R. 1987. Site directed mutragenesis by gene targeting in mouse embryo-derived stem cells. Cell. 51:503-512.
56. Thomas, K. R. and Capecchi, M. R. 1990. Targeted disruption of the murine int-1 proto-oncogene resulting in severe abnormalities in midbrain and cerebellar development. Nat. 346:847-850.
57. Thomson, J. A., Itskovitz-Eldor. J., Shapiro, S. S., Waknitz, M. A., Swiergiel, J. J., Marshall, V. S. and Jones, J. M. 1998. Embryonic stem cell lines derived from human blastocysts. Sci. 282:1145-1147.
58. Toyooka, V., Tsunekawa, N., Akasu, R. and Nou, T.
2003. Embryonic stem cells can form germ cells in vitro. Proc. Natl. Acad. Sci. USA. 100:11457-11462.
59. Wagner, R. C. 1980. Endothelial cell embryology and growth. Adv. Microcirc. 9:45-75.
60. Wheeler, M. B. and Choi, S. J. 1997. Embryonic stem cells and transgenics: Recent advances. Arch. Fac. Vet.
Universidade Federal, Rio Grande do Sul, Brazil. 25:64–
83.
61. Wheeler, M. B. and Walters, E. M. 2001. Transgenic Technology and applications in Swine. Theriogenology.
56:1345–1370.
62. Wilmut, I., Schnieke, A. E., Mc Whir, J., Kind, A. J.
and Campbell, K. H. 1997. Viable offspring derived from fetal and adult mammalian cells. Nat. 385:810 -813.
63. Yanagimachi, R. 2002. Cloning: experience from the mouse and other animals. Mol. Cell Endocrinol.
187:241-248.
64. Zandstra, P. W., Bauwens, C. and Yin, T. 2003.
Scalable production of embryonic stem cell-derived cardiomyocytes. Tissue Eng. 9:767–78.
(투고일: 2011.03.02. 수정일: 2011.05.24. 판정일: 2011.05.25.)
[Provider:earticle] Download by IP 118.70.52.165 at Monday, December 20, 2021 8:06 PM