Introduction
About 29% of freshwater fish are native species that habitat only in Korea and totally hundreds of these species have been reported to belonging to the order Cypriniformes by now (Kim and Park, 2002). Some species are threatened or endan- gered, so cytogenetic and molecular studies should be done to provide necessary information for conservation of these species. Among the endangered species, Microphysogobio longidors-
alis M
ORI, 1935 belonging to Gobioninae, a sub- family of Cyprinidae, is a native species of Korea and usually distributes in the midstream and upstream of rivers. However, due to the environ- mental pollution and diminishment of habitat, the number of this species has decreased sharply and population ecology as well as taxonomy has been studied recently for the purpose of conser- vation of this species (Kim, 1984; Song and Son, 2003).
The chromosome numbers and karyotypes of species of genus Cobitis, genus Moroco and genus Iksookimia belonging to Cypriniformes have been
─
─ 189 ──
Cytogenetic and Molecular Studies of Endangered Freshwater Species from Korea I. Microphysogobio longidorsalis Mori
(Cyprinidae: Gobioninae)
Jae Hyun Im, Wan-Ok Lee
1, Lei Peng, Jae Koo Noh
2, Yoon Kwon Nam and Dong Soo Kim*
Department of Aquaculture, Pukyong National University, Busan 608-737, Korea
1Inland Fisheries Ecological Research Institute, National Fisheries Research and Development Institute, Chongpyong 477-810, Korea
2Fish Genetics and Breeding Research Center, National Fisheries Research and Development Institute, Geoje 656-842, Korea
The population of Microphysogobio longidorsalis has decreased sharply in Korea due to environmental pollution and diminishment of habitat. For conservation and stock recruitment of this species, cytogenetic and molecular studies should be carried out to provide basic genetic information. In total, 122 metaphase plates were observed, and the results revealed that the chromosome number was 50, with 9 metacentric pairs, 13 submetacentric pairs and 3 acrocentric pairs. Ag-- stained nucleolar organizer regions (Ag-- NORs) localized on one pair of submetacentric chromosomes were found. DNA content was 2.6 pg/cell. The molecular structure of the full sequence of cytochrome b (cyto b) was typical of those observed in several fish taxa. A 623--bp segment of 12S ribosomal RNA gene has been sequenced. These sequences were compared with those of common carp, crucian carp and Atlantic salmon. Combination of the results of sequence analysis in this study indicated that a close relationship existed among Microphysogobio longidorsalis, common carp and crucian carp. The results of the present study would be useful for genetic improve- ment and conservation of this species.
Key words : Microphysogobio longidorsalis, Ag-- NORs, DNA content, cyto b, 12S rRNA
*Corresponding author: [email protected]
studied (Lee et al., 1983; Lee, 1984; Lee et al., 1986; Kim et al., 1995; Kim et al., 2003). How- ever, no study has been done on chromosome number and karyotype of Microphysogobio longi- dorsalis. So, cytogenetic analysis to provide basic information for conservation and stock recruit- ment of this species should be conducted. The diploid numbers of chromosomes have been repo- rted varying from 2 in some nematodes to over 200 in some fish species and is a characteristic for a given species in most cases (Kim et al., 1982, Kim et al, 1988; Park and Lee, 1996; Kim et al., 2001). Also, the number of chromosome arms (NF value) is important for studies on chro- mosomes evolution or fish breeding since this defines the genetic content of a chromosome complement. In addition, karyotype analysis has been used as a tool for genome mapping which is useful for genetic improvement in fish culture.
Physical and genetic characterization of chromo- somes is also a useful tool in studying of both normal and pathologic molecular physiology of a given species.
Though chromosome and karyotype of a given species provide basic and necessary information for cytogenetic research, other modern methods that are commonly applied including Ag-stain- ing of nucleolar organizer regions (Ag-NORs) and fluorescence in situ hybridization (FISH) can give more deep insight into the specificity of a species, especially when used for phylogenetic study. Nucleolar organizer regions (NORs) are loops of DNA coding for ribosomal RNA (rRNA).
They are associated with non-histone nucleao- protein and hence can be identified by silver staining. The Ag-NORs has been a useful chro- mosome marker, revealing not only interspecific difference, but also interchromosomal differ- ences. Numbers and location of Ag-NORs were studied in the present study.
Recently, with the quick development of tech- nologies of molecular biology, different molecular markers have been developed and considered to be remarkably versatile molecular tools. Nucl- eotide sequencing data are considered to be one of the most appropriate tools to detect genetic variation and DNA sequencing of the cytochrome region of the mtDNA is very specific since it allows the differentiation at the individual level.
Mitochondrial DNA cytochrome b (cyt b) gene is a well-known protein-coding marker and it has been one of the most frequently used segments of mtDNA because of the availability of universal
amplification primers (Kocher et al., 1989). Esp- ecially, it would be useful where a definite answ- er is not possible with other methods (Partis and Wells, 1996; Apostolidis et al., 2001). A full seq- uence of cyt b was determined in the present study.
In addition, other portions of the mtDNA such as the rRNA genes have been shown to be useful for determining relationships at different taxo- nomic levels and it is believed that accumulated sequence data on the mtDNA would be a valu- able tool for elucidating various higher order relationships (Apostolidis et al., 2001). Comm- only, cytochrome or noncoding control regions are proved to be useful in resolving relationships among closely related taxa, whereas the 12S or 16S rRNA gene sequences have been used to examine the phylogenetic relationships among higher taxa such as genus, family, or order (Sumida et al., 2000). Also, 12S rRNA has been proved to be promising tools for tracing the his- tory of more recent evolutionary events (Hills and Dixon, 1991). A segment of 12S rRNA gene was sequenced and compared with those of related species.
The aim of present study is to provide basic information on the karyotype of Microphysogobio longidorsalis. In addition, the number and local- ization of Ag-NORs, cell and nuclear size and DNA content were studied. The genetic charac- teristics were further studied by comparison of sequences of mitochondrial cyt b and 12S riboso- mal RNA regions with those of related species. A 1,141-bp region of the mt cyt b gene and a 623 bp segment of 12S ribosomal RNA gene were sequenced and the genetic variation was asse- ssed based on these sequence data. The results of the present study would be useful for conser- vation and genetic improvement of Microphy- sogobio longidorsalis.
Materials and Methods
1. Specimens and chromosome preparation
Seven adult specimens (3 female and 4 male)
which were caught from Imjin River, Kyonggi-
do, Korea were obtained and maintained in a
simple recirculating aquaculture system until
sacrificed. Chromosome slides were prepared by
the standard kidney procedure with some mod-
ification (Kim et al., 1995). Brief ly, f ish received an intraperitoneal injection of colchicine (Sigma, USA) at a concentration of 0.1 mg/ml (0.1 ml/10 g fish body weight) and after several hours, fish were sacrificed and kidney cell were collected, teased and suspended in hypotonic solution, then supernatants were collected and to which Carnoy’s fixative was added. The cell suspen- sions were treated repeatedly 3 times with fresh Carnoy’s fixative, stored in 4� C refrigerator and centrifugation at 1,500 g for 5 min. Finally, the samples were spread onto well-prepared slide glass and stained for 20 min with 5% Giemsa in Sörenson’s phosphate buffer (pH 6.8). Well spre- ad metaphases were selected and photographed.
The chromosomes were classified following Levan’s nomenclature (Levan et al., 1964).
2. Cell and nuclear size of erythrocytes
The blood from each sample was extracted in 1 ml syringe and smeared on glass slide, then fixed with 95% ethanol and stained with Giemsa staining solution. Totally more than 100 cells of each specimen were checked and recorded. The major axis (a) and minor axis (b) of cell and nuc- lear were measured with eyepiece micrometer.
Surface area (s) and volume (v) were calculated as (s) = ab
π/4 and (v) = 4(a/2)(b/2)
2π/3, respec- tively, according to Sezaki and Kobayashi (1978).
3. Ag-
-NOR and f low cytometric analysis One-step silver nitrate method was used to identify the Ag-NORs (Howell and Black, 1980).
To measure the DNA content, f low cytometry was performed using a PA II f low cytometer (Partec, Germany) according to the protocol pro- vided by the manufacturer. Briefly, 100~150
µl blood samples were centrifuged at 200 g for 10 min and the concentrated erythrocytes were res- uspended in FPS. The cell solution was stained with DAPI reagent and applied for f low cytome- try analysis. Blood cells of common carp, Cypri- nus carpio, and rock bream, Oplegnathus fascia- tus, were used as controls for estimation of rela- tive DNA contents of Microphysogobio longidor- salis.
4. DNA extraction, amplification and sequencing
Total DNA was extracted from liver tissue from each individual by using conventional SDS/
proteinase K method as described by Nam et al.
(1998). Briefly, samples were digested in a solu- tion of 7 M urea, 10 mM Tris, 100 mM EDTA, pH 8.0, 125 mM NaCl, 1% SDS, 100
µg/ml proteinase K at 55� C overnight, and DNA was extracted using TE (10 mM Tris, 1 mM EDTA, pH 8.0)- saturated phenol, phenol/chloroform (1 : 1), and chloroform/isoamyl alcohol (24 : 1) extractions.
DNA was precipitated by adding an equal vol- ume of isopropanol followed by centrifugation.
The DNA pellet was rinsed with 70% ethanol, air-dried, and resuspended in TE buffer.
The full sequences of the cyt b genes were amp- lified via PCR from each individual. The primers used were specifically constructed as: forward primer, 5
′-AGCCTACGGAAAAC-3′and reverse primer, 5
′-CACTGAAATGAGCTT-3′. PCR mix- tures were prepared in 25
µl with a final concen- tration of 100 nanograms of template DNA and 50
µM primers, using PCR PriMix according to the manufacturer’s specification (Bioneer, Korea).
The amplification process was conducted as fol- lows: an initial cycling at 94� C for 2 min followed by 30 cycles of 94� C, 1 min; 55� C, 1 min; 72� C, 1 min, and a final extension of 72� C for 5 min.
The primers used in amplification of 12S ribo- somal RNA were universal primers R1091 (5
′- AAAAAGCTTCAAACTGGGATTAGATACCCCA CTAT-3′) and R1478 (5′-TGACTGCAGAGGGT GACGGGCGGTTGTGT-3′ ). One hundred nano- grams of template gDNA and 50
µM above men- tioned primers were placed in PCR PriMix accord- ing to the manufacturer’s specification (Bioneer, Korea). PCR were performed under the following conditions: a 2-min denaturation at 94� C, fol- lowed by 35 cycles of 50 seconds at 94� C; 50 seconds at 58� C; 1 min at 68� C, and finally, an extra extension step of 7 min at 68� C.
Amplified DNA was fractionated by electropho- resis through 0.6% agarose gels, recovered from the gels and purified with Gel Purification Kit (Bioneer, Korea) according to the supplier’s pro- tocol. Purified products were applied to sequenc- ing reaction with BioDye terminator sequencing Kit (Applied Biosystem Co., USA). Effort was made to avoid mislabeling and contamination.
Amino acid sequences were deduced from the nucleotide sequences of the cyt b gene using BLASTX program (http://www.ncbi.nlm.nih/
BLAST/). Sequences were aligned with those of
common carp, crucian carp and Atlantic salmon
using CLUSTAL W program (Thompson et al.,
1994) and verified visually.
Results
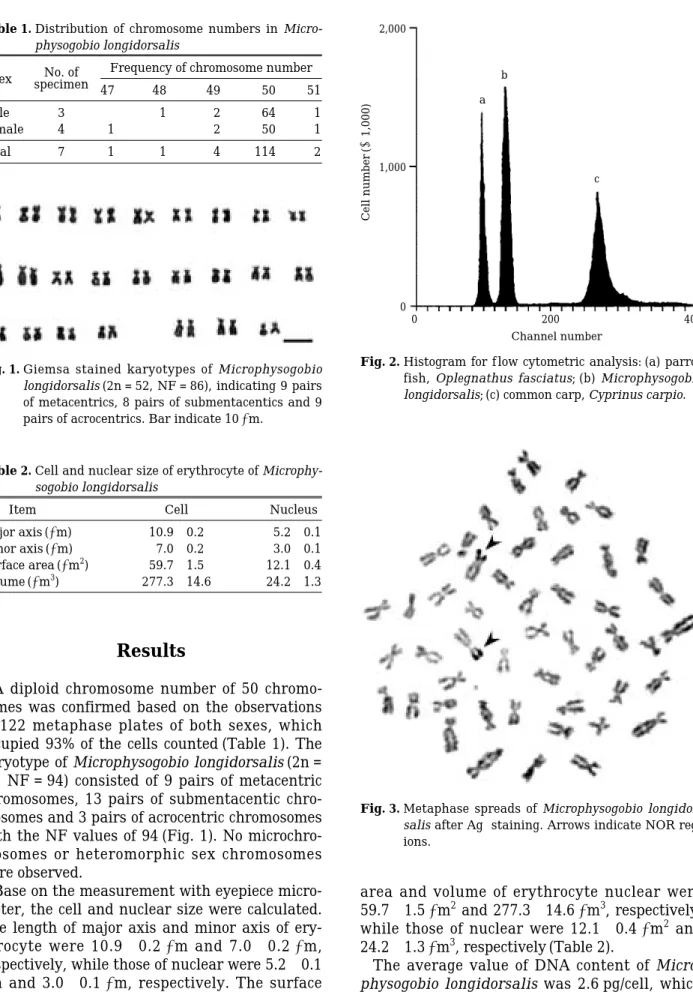
A diploid chromosome number of 50 chromo- somes was confirmed based on the observations of 122 metaphase plates of both sexes, which occupied 93% of the cells counted (Table 1). The karyotype of Microphysogobio longidorsalis (2n = 50, NF = 94) consisted of 9 pairs of metacentric chromosomes, 13 pairs of submentacentic chro- mosomes and 3 pairs of acrocentric chromosomes with the NF values of 94 (Fig. 1). No microchro- mosomes or heteromorphic sex chromosomes were observed.
Base on the measurement with eyepiece micro- meter, the cell and nuclear size were calculated.
The length of major axis and minor axis of ery- throcyte were 10.9±0.2
µm and 7.0±0.2
µm, respectively, while those of nuclear were 5.2±0.1
µm and 3.0±0.1
µm, respectively. The surface
area and volume of erythrocyte nuclear were 59.7±1.5
µm
2and 277.3±14.6
µm
3, respectively, while those of nuclear were 12.1±0.4
µm
2and 24.2±1.3
µm
3, respectively (Table 2).
The average value of DNA content of Micro- physogobio longidorsalis was 2.6 pg/cell, which
Table 1. Distribution of chromosome numbers in Micro- physogobio longidorsalis
Sex No. of Frequency of chromosome number
specimen 47 48 49 50 51
Male 3 1 2 64 1
Female 4 1 2 50 1
Total 7 1 1 4 114 2
Fig. 1. Giemsa stained karyotypes of Microphysogobio longidorsalis (2n = 52, NF = 86), indicating 9 pairs of metacentrics, 8 pairs of submentacentics and 9 pairs of acrocentrics. Bar indicate 10µm.
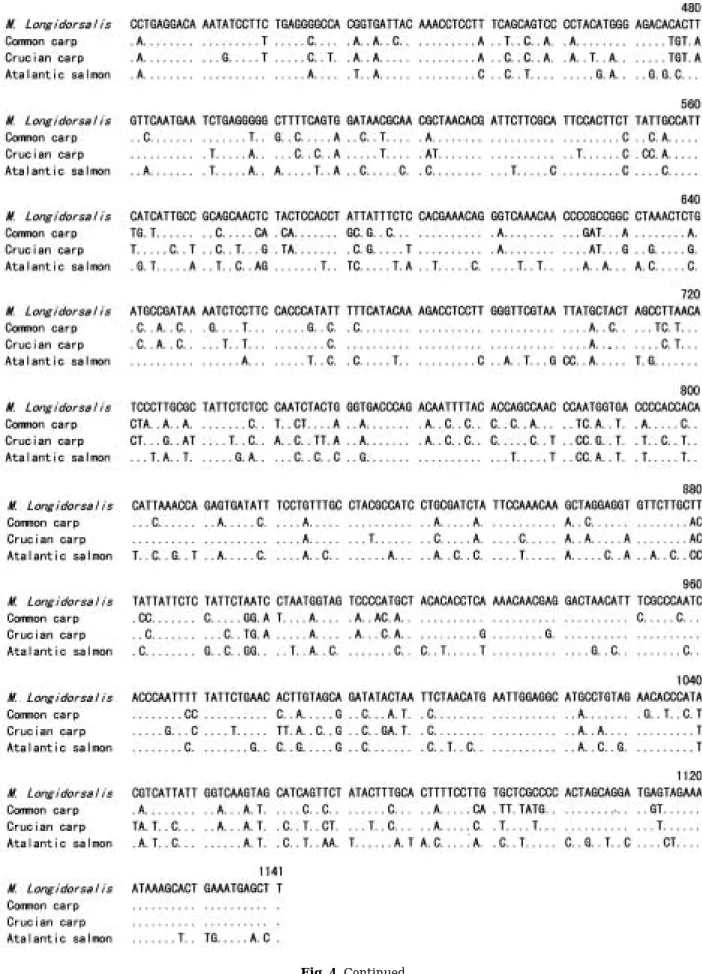
Fig. 2. Histogram for f low cytometric analysis: (a) parrot fish, Oplegnathus fasciatus; (b) Microphysogobio longidorsalis; (c) common carp, Cyprinus carpio.
Table 2. Cell and nuclear size of erythrocyte of Microphy- sogobio longidorsalis
Item Cell Nucleus
Major axis (µm) 10.9±0.2 5.2±0.1
Minor axis (µm) 7.0±0.2 3.0±0.1
Surface area (µm2) 59.7±1.5 12.1±0.4
Volume (µm3) 277.3±14.6 24.2±1.3
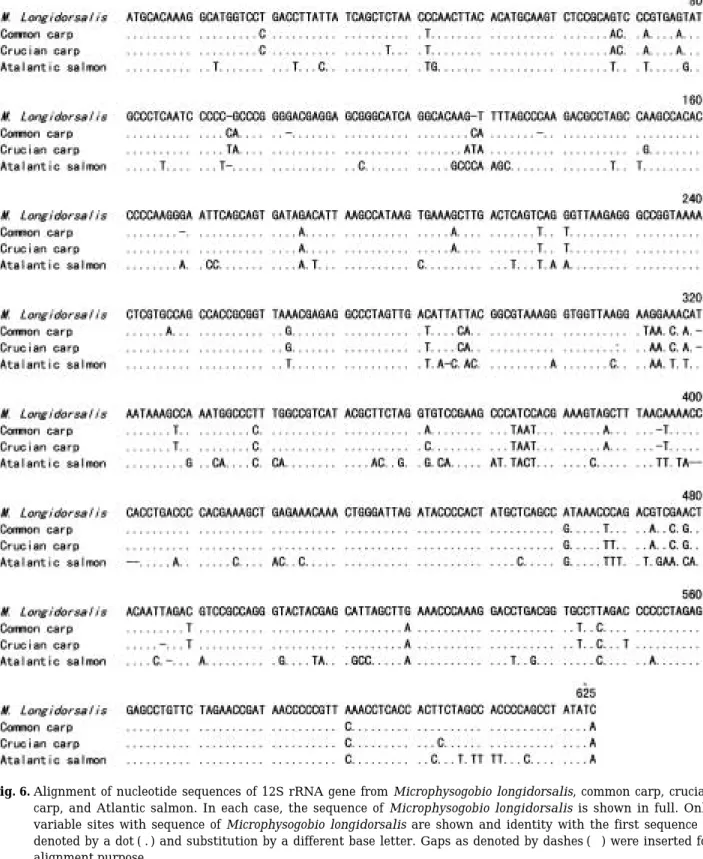
Fig. 3. Metaphase spreads of Microphysogobio longidor- salis after Ag-staining. Arrows indicate NOR reg- ions.
2,000
1,000
0
Cell number(×1,000)
a b
c
0 200 400
Channel number
was calculated based on the known DNA con- tents of rock bream (1.8 pg/cell) and common carp (3.6 pg/cell) (Fig. 2). This value was about 72% of that of common carp. Ag-NORs were analyzed in 27 metaphases and only one pair of Ag-NORs located on a pair of submetacentric chromosomes was observed (Fig. 3).
A total of 1141 bp of the cyt b was determined, including no insertions or deletions (Fig. 4). Base composition in this sequence was as follows: low G content (15.2%) and almost equal A, C, and T contents (25.7%, 30.2%, and 28.9%, respectively).
Nucleotide sequence identities of Microphyso- gobio longidorsalis with those of common carp, crucian carp and Atlantic salmon were 83%, 82%
and 79%, respectively (Table 3). However, the deduced amino acid sequence identities were much higher, with the corresponding values of 93%, 94% and 91%, respectively (Table 3). Align- ment of the amino acid sequence of Microphy- sogobio longidorsali with those of common carp,
crucian carp and Atlantic salmon was shown in Fig. 5.
Nucleotide sequence of the 12S rRNA was deter- mined in the 623-bp segment of the mtDNA gene. Among 7 samples analyzed, this 623-bp segment provided no variable site (Fig. 6). When compared with common carp and crucian carp,
Table 3. Similarity on nucleotides and amino acid resi- dues among common carp, crucian carp, Atlantic salmon, and Microphysogobio longidorsalis.
Based on cytochrome b gene sequences
Cytochrome b M. longidorsalis Crucian Common Atlantic
(nt/aa) carp carp salmon
(1135/380) (1141/380) (1141/380) (1141/380) M. longidorsalis -
Crucian carp 0.82/0.94* -
Common carp 0.83/0.93 0.88/0.98 -
Atlantic salmon 0.79/0.91 0.76/0.91 0.77/0.90 -
*This indicates the % similarity of nucleotide/amino acid residues of different species.
Fig. 4. Alignment of nucleotide sequences of mitochondrial cytochrome b gene from Microphysogobio longidorsalis, com- mon carp, crucian carp, and Atlantic salmon. In each case, the sequence of Microphysogobio longidorsalis is shown in full. Only variable sites with sequence of Microphysogobio longidorsalis are shown and identity with the first sequence is denoted by a dot ( . ) and substitution by a different base letter.
Fig. 4. Continued.
two gaps were found in this 12S rRNA sequence.
The A, G, C and T contents were 29.7%, 22.0%, 18.6% and 29.7%, respectively. A slightly A-T nucleotide bias was presented in this gene, about 59.4%. Nucleotide sequence identities of Micro- physogobio longidorsalis with those of common carp, crucian carp and Atlantic salmon were 92
%, 91% and 82%, respectively (Table 4).
Discussion
The chromosome number was found to be 50 in the present study. In the studies of the kary- otypes of 7 species of the Gobioninae fishes and three Cyprinid species of genus Moroco, diploid chromosomes numbers were found to be 50 for all the species examed (Lee, 1984; Lee et al., 1987). The result of the present study was in agreement with them. Though these species studied in their research and Microphysogobio longidorsalis studied here have the same chro- mosome number, their karyotypes were different from each other, as Pseudogobio esocinus, Sarco- cheilichthys nigripinnis morii and Pungtungia herzi having 8 metacentric chromosomes; Gna- thopogon strigatus, Squalidus gracilis majimae and Hemibarbus longirostris having 7 metacen- tric chromosomes; and Coreoleucisus splendidus
Fig. 5. Comparisons of deduced amino acid sequences of mitochondrial cytochrome b gene from Microphysogobio longi- dorsalis, common carp, crucian carp and Atlantic salmon. In each case, the sequence of Microphysogobio longidor- salis is shown in full. Only variable sites with sequence of Microphysogobio longidorsalis are shown. Identical amino acid residues with the first sequence are denoted by a dot (.) and substitution by a different base letter.
Table 4. Similarity on nucleotides among common carp, crucian carp, Atlantic salmon, and Microphy- sogobio longidorsalis. Based on 12S ribosomal RNA gene sequences
12S rRNA M. longidorsalis Crucian Common
Atlantic
carp carp salmon
M. longidorsalis -
Crucian carp 0.92 -
Common carp 0.91 0.98 -
Atlantic salmon 0.82 0.83 0.82 -
having only 6 metacentric chromosomes. The acrocentric and telocentric chromosomes num- bers of each species studied were either 3 or 4.
Also, the NF values were either 92 or 94, indi- cating no great difference among these species.
Arai (1982) concluded that the diploid chromo-
Fig. 6. Alignment of nucleotide sequences of 12S rRNA gene from Microphysogobio longidorsalis, common carp, crucian carp, and Atlantic salmon. In each case, the sequence of Microphysogobio longidorsalis is shown in full. Only variable sites with sequence of Microphysogobio longidorsalis are shown and identity with the first sequence is denoted by a dot ( . ) and substitution by a different base letter. Gaps as denoted by dashes (-) were inserted for alignment purpose.
some number of 50 was the most primitive after comparison of the karyotypes of Cyprinid species and proposed that new species originated from these primitives via chromosome fusion or fis- sion, a process including a few rearrangements such as conversion of metacentric to submenta- centric or vice versa (Uribe-Alcocer et al., 2002).
Also, Cypriniformes are believed to be primitive because they have more subtelocentric to telo- centric chromosomes (Campos et al., 1997).
Dipnoan, Protopterus sp., has been reported to show tendency to change, from ancient to more recent genera, towards a reduction in chromo- some number (Morescalchi et al., 2002). A thro- ugh comparison of the karyotypes of as many species as possible would be required to avoid making arbitrary conclusions on the revolutio- nary of these species.
Cyprinid species from Korea previously studied have been reported to be absence of heteromor- phic sex chromosomes (Lee, 1984; Lee et al., 1986; Lee et al., 1987). In fish, approximately 80 species, which corresponds to 5% of the fish who- se karyotype had been analyzed to date, were found to have heteromorphic sex chromosomes (Ueno et al., 2001). However, evidence for and against sex chromosome differentiation exists (Ozouf-Costaz et al., 1990; Pandey and Larka, 1997).
The DNA content of Microphysogobio longidor- salis obtained here was 2.6 pg/cell. Higher DNA content of 80 pg/cell has been reported for lung fish (Morescalchi et al., 2002). Using flow cyto- metry to analyze DNA content of a given species is one of the most effective methods in terms of protecting the experimental animals from injury.
Also, it is a convenient way to identify ploidy and to analyze hybrids cytogenetically. The cell size and nuclear size of erythocytes were measured in the present study. This method is considered to be the most convenient for determination of ploidy level and success or failure of hybridiza- tion (Sim et al., 2002). The results of the present experiment can be used for these purposes.
The most frequent localization of Ag-NORs in fish is telomeric (Gold, 1984) and Ag-NORs sites have generally been detected on a single chro- mosomal site in numerous species of bony fish (Rocco et al., 2002). In cyprinid species, Ag- NORs located on a single pair of chromosomes have also been reported (Lee et al., 1986; Lee et al., 1987). However, these reported Ag-NORs together with that detected in the present study
are all located on submetacentric chromosomes.
Multiple Ag-NORs sites have been found in thr- ee species of genus Salvelinus and other cyp- rinid species (Zhuo et al., 1995; Li et al., 1997).
The Ag-NORs have been a useful chromosome marker. Their number, location and size are often species specific and Ag-NORs have been reported to be highly polymorphic in fish (Castro et al., 1996). Unfortunately, the Ag-NOR pat- terns in Korean cyprinid species have not been thoroughly studied. In order to obtain the pat- tern of NORs evolution, more species from differ- ent families of this order should be analyzed to get accurate picture of numbers, sizes and loca- tions of Ag-NORs.
Though Ag-NORs are very useful for cytoge- netic study, they do not reveal all the sites on chromosomes containing ribosomal DNA (rDNA) genes (Li and Havenhand, 1997). Ag-NORs are commonly regarded as only capable of detecting active rDNA sites and represent rDNA transcrip- tion activity or transcription potential. Fluores- cence staining using FISH method which can allow to precisely identify the NOR-bearing chromosomes should be employed to further define the repeated DNA families in regardless of their activation status.
The cyt b gene was 1141 bp long in all the spec- imens sequenced and its molecular structure was typical of those observed in several fish taxa (Chang et al., 1994; Zardoya et al., 1995; Rocha- Olivares et al., 1999). Though the identities of nucleotide sequences were low, the identities of deduced amino acid sequence were much higher, indicating high conservation for amino acid sequence. This result was consistent with pre- vious studies that inferred an exceptionally low rate of amino acid replacement in the fish lin- eage, only about a fifth as rapid as in mammals or birds (Adachi et al., 1993; Cantatore et al., 1994; Briolay et al., 1998; Apostolidis et al., 2001).
A slightly bias towards A±T in the sequence of
12S rRNA gene was noted. The mitochondrial
genomes of arthropod and scleractinian corals
have been reported to be highly A±T biased
(Zehethofer and Sturmbauer, 1998; Chen et al.,
2002). The mitochondrial 12S rRNA is believed
to evolve at a relatively slow evolutionary rate
(Simons and Mayden, 1998). This gene may thus
be especially suitable for resolving relationships
between more distantly related taxa. The low
sequence variation of 12S rRNA gene confirmed
that there was no great genetic differentiation
between cyprinid species analyzed herein. Other than being used as a reliable tool for genetic study, 12S rRNA gene has been used for study of gene evolution (Chen et al., 2002). The results of the present experiment also can be used for this purpose with the accumulation of mtDNA seque- nces data for cyprinid species. Mitochondrial genomes are maternally inherited and difference between the distributions of the haplotypes between male and female are not expected (van der Veer et al., 2003). The results of the present experiment agreed with this, as no variation existed between male and female samples analy- zed.
Combination of the results obtained through comparisons of cyt b and 12S rRNA indicated that Microphysogobio longidorsalis was closely related to common carp and crucian carp, where- as a great genetic distance of Atlantic salmon to cyprinid species was noted.
In conclusion, a diploid chromosome number of 50 was confirmed with the karyotype as: 9 pairs of metacentrics; 13 pairs of submentacentrics;
and 3 pairs of acrocentrics. Only one pair of Ag- NOR was found to be located on a pair of sub- metacentrics. Full sequence of cyt b and a 623- bp segment of 12S rRNA gene have been sequ- enced and compared with common carp, crucian carp and Atlantic salmon. The molecular struc- ture of cyt b was typical of those observed in several fish taxa. Combination of results of sequ- ence analysis indicated that a close relationship existed among Microphysogobio longidorsalis, common carp and crucian carp. The results of the present study can serve as database for gen- etic improvement and conservation of Microphy- sogobio longidorsalis.
References
Adachi, J., C. Ying and M. Hasegawa. 1993. Tempo and mode of mitochondrial DNA in vertebrates at the amino acid sequence level: rapid evolution in warm-blooded vertebrates. J. Mol. Evol., 36:
270~281.
Apostolidis, A.P., Z. Amuris and C. Triantaphy- llidis. 2001. Phylogenetic relationships among four species of Mullidae (Perciformes) inferred from DNA sequences of mitochondrial cytochro- me b and 16S rRNA genes. Biochem. Syst. Ecol., 29: 901~909.
Arai, R. 1982. A chromosome study on two cyprinid fishes, Acrossocheilus labiatus and Psedorasbora
pumila pumila, with notes on Eurasian cyprin- ids and their karyotypes. Bull. Nat. Sci. Mus., Tokyo, Ser. A, 8: 131~152.
Briolay, J., N. Galtier, R.M. Brito and Y. Bouvet.
1998. Molecular phylogeny of cyprinidae infer- red from cytochrome b DNA sequences. Mol.
Phylogenet. Evol., 9: 100~108.
Campos, H., G. Arratia and C. Cuevas. 1997. Kary- otypes of the most primitive catfishes (Teleostei:
Siluriformes: Diplomystidae). J. Zool. Sys. Evol.
Res., 35: 113~119.
Cantatore, P., M. Roberti, G. Pesole, A. Ludovico, F.
Milella, M.N. Gadaleta and C. Saccone. 1994.
Evolutionary analysis of cytochrome b sequen- ces in some perciformes: Evidence for a slower rate of evolution than in mammals. J. Mol.
Evol., 39: 589~597.
Castro, J., A. Arias, S. Rodriguez, L. Sanchez and P.
Martinez. 1996. Population analysis of a nucle- olar organizer region number polymorphism in brown trout (Salmo trutta). Chromosome Res., 3:
45~46.
Chang, Y.-S., F.-L. Huang and T.-B. Lo. 1994.
The complete nucleotide sequence and gene org- anization of carp (Cyprinus carpio) mitochon- drial genome. J. Mol. Evol., 38: 138~155.
Chen, C.A., C.C. Wallace and J. Wolstenholme.
2002. Analysis of the mitochondrial 12S RNA gene supports a two-clade hypothesis of the evo- lutionary history of Scleractinian corals. Mol.
Phylogenet. Evol., 23: 137~149.
Gold, J.R. 1984. Silver-staining and heteromor- phism of chromosomal nucleolous organizer regions in North American cyprinid fishes.
Copeia, 1: 133~139.
Hills, D.M. and M.T. Dixon. 1991. Ribosomal DNA:
molecular evolution and phylogenetic inference.
Rev. Biol., 66: 411~453.
Howell, W.M. and D.A. Black. 1980. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: 1-step met- hod. Experientia, 36: 1041~1045.
Kim, D.S., E.H. Park and J.S. Kim. 1982. Karyo- types of nine species of the Korean catfish (Tele- ostomi: Siluriformes). Korean J. Genet., 4: 57~ 68 (in Korean).
Kim, D.S., I.B. Kim, H.T. Huh and I.S. Park. 1988.
Cytogenetic analysis of catfish, Silurus asotus (Teleostomi: Siluriformes). Ocean Res., 10: 33~
37 (in Korean).
Kim, D.S., Y.K. Nam and I.S. Park. 1995. Survival and karyological analysis of reciprocal diploid and triploid hybrids between mud loach (Misgur- nus mizolepis) and cyprinid loach (Misgurnus anguillicaudatus). Aquaculture, 135: 257~265.
Kim, D.S., H.J. Cho, I.S. Park, J.C. Choi and Y.K.
Nam. 2001. Cytogenetic traits and gonad devel- opment of induced triploid far eastern catfish, Silurus asotus. Korean J. Genet., 23: 55~62.
Kim, I.S. 1984. The taxonomic study of subgenus of the subfamily Gobioninae (Cyprinidae) in Korea.
Bull. Korean Fish. Soc., 17: 436~448 (in Korean).
Kim, I.S. and J.S. Park. 2002. Freshwater fisheries of Korea. Kyo-hak Publishing. 2002. pp. 94~
156 (in Korean).
Kim, I.S., E.K. Choi, H. Yang and M.H. Koh. 2003.
Karyotype analysis of Southern spined loach, Iksookimia hugowolfeldi (Pisces, Cobitidae).
Korean J. Ichthyol., 15: 127~129 (in Korean).
Kocher, T.D. W.K. Thomas, A. Meyer, S.V. Edwa- rds, S. Paabo, F.X. Villablanca and A.C. Wilson.
1989. Dynamics of mitochondrial DNA evolution in animals: amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA., 86: 6196~6200.
Lee, H.Y., C.H. Yu, S. Jeon and H.S. Lee. 1983. The karyotype analysis on 29 species of fresh-water fish in Korea. Bull. Inst. Basic Sci., Inha Univ., 4: 79~86.
Lee, G.Y. 1984. Karyotypes of seven species of the Gobioninae fishes (Cyprinidae) in Korea. Korean J. Ichthyol., 17: 24~34. (in Korean)
Lee, H.Y., H.S. Lee and C.S. Chang. 1986. Karyo- type analysis and geographical polymorphism in Korean Cobitis. Korean J. Genet., 8: 65~74 (in Korean).
Lee, H.Y., H.S. Lee, J.W. Cho and S.R. Jeong. 1987.
Karyotype analysis of Korean fresh water fish, Moroco semotilus (Pisces; Cyprinidae). Korean J.
Genet. 9: 176~180 (in Korean).
Levan, A., K. Fredga and A.A. Sandberg. 1964.
Nomenclature of centromeric position on chro- mosomes. Hereditas, 52: 201~220.
Li, X.X. and J.N. Havenhand. 1997. Karyotype, nucleolus organizer regions and constitutive hete- rochromatin in Ostrea angasi (Molluscae: Biva- lvia): evidence of taxonomic relationships within the Ostreidae. Marine Biol., 127: 443~448.
Morescalchi, M.A., L. Rocco and V. Stingo. 2002.
Cytogenetic and molecular studies in a lungfish, Protopterus annectens (Osteichthyes, Dipnoi).
Gene, 295: 279~287.
Nam, Y.K., C.H. Jeong and D.S. Kim. 1998. A simple clearing method for improved visual- ization of LacZ expression in fish larvae. Trans- genic Res., 135: 257~265.
Ozouf-Costaz, C., G.G. Teugels and M. Legendrc.
1990. Karyological analysis of three strains of the African catfish Clarias gariepinus (Clari- idae) used in aquaculture. Aquaculture, 87: 271
~277.
Pandey, N. and W.S. Larka. 1997. Evidence of fem- ale heterogamety, B-chromosome and natural tetraploidy in the Asian catfish, Clarias batra- chus, used in aquaculture. Aquaculture, 149:
31~37.
Park, I.S. and C.L. Lee. 1996. Cytogenetic analysis
of Bagarid catfish Pseudobagrus fulvidraco (Teleostomi: Siluriformes). Korean J. Ichthyol., 8: 10~15.
Partis, L. and R.J. Wells. 1996. Identification of fish species using random amplified polymorphic DNA (RAPD). Mol. Cell. Probes., 10: 435~441.
Rocco, L., M.A. Morescalchi, D. Costagliola and V.
Stingo. 2002. Karyotype and genome chara- cterization in four cartilaginous fishes. Gene, 295: 289~298.
Rocha-Olivares, A., R.H. Rosenblatt and R.D.
Vetter. 1999. Molecular evolution, systematics, and zoogeography of the rockfish subgenus Sebastomus (Sebastes, Scorpaenidae) based on mitochondrial cytochrome b and control region sequences. Mol. Phylogenet. Evol., 11: 411~458.
Sezaki, K. and H. Kobayashi. 1978. Comparison of erythrocytic size between diploid and tetrapolid in spinous loach, Cobitis biwae. Bull. Jap. Soc.
Sci. Fish., 41: 851~854.
Sim, M.A., J.K. Noh, Y.K. Nam and D.S. Kim. 2002.
Cytogenetic analysis of spotty belly greenling (Hexagrammos agrammus) and greenling (H.
otakii). J. Korean Fish. Sci., 35: 682~685.
Simons, A.M. and R.L. Mayden. 1998. Phylogenetic relationships of the western North American Phoxinins (Actinopterygii: Cyprinidae) as infer- red from mitochondrial 12S and 16S ribosomal RNA sequences. Mol. Phylogenet. Evol., 9: 308~ 329.
Song, H.B. and Y.M. Son. 2003. Population ecology of Microphysogobio longidorsalis (Cyprinidae) from Korea. Korean J. Ichthyol., 15: 303~310.
Sumida, M., M. Ogata and M. Nishioka. 2000. Mole- cular phylogenetic relationships of pond frogs distributed in the Palearctic regions inferred from DNA sequences of mitochondrial 12S riboso- mal RNA and cytochrome b genes. Mol. Pylog- enet. Evol., 16: 278~285.
Thompson, G.D., D.G. Higgins and T.J. Gibson.
1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment thro- ugh sequence waiting position specific gap pen- alty’s and weight matrix choice. Nucleic Acids Res., 22: 4673~4680.
Ueno, K., K. Ota and T. Kobayashi. 2001. Hetero- morphic sex chromosomes of lizard fish (Synodo- ntidae): focus on the ZZ-ZW1W2 system I Trac- hinocephalus myops. Genetica, 111: 133~142.
Uribe-Alcocer, M., H. Olvera-García and P. Díaz- Jaimes. 2002. Karyotype analysis in three spe- cies of the genus Chirostoma (Atheriniformes:
Atherinidae). Ichthyological Res., 49: 85~88.
Van der Veer, M., K. Kanobana, H.W. Ploeger and E. de Vries. 2003. Cytochrome oxidase c subunit I polymorphisms show significant differences in distribution between a laboratory maintained population and a field isolates of Cooperia onco- phora. Vet. Parasitol., 116: 213~238.
Zardoya, R., A. Garrido-Pertierra and J.M. Bauti- sta. 1995. The completer nucleotide sequence of the mitochondrial DNA genome of the rainbow trout, Oncorhynchus mykiss. J. Mol. Evol., 41:
942~951.
Zehethofer, K. and C. Sturmbauer. 1998. Phylo- genetic relationships of central European wolf
spinders (Araneae: Lycosidae) inferred from 12S ribosomal DNA sequences. Mol. Phylogenet.
Evol., 10: 391~398.
Zhuo L., K.M. Reed and R.B. Phillips. 1995. Hyper- variability of ribosomal DNA at multiple chro- mosomal sites in lake trout (Salvelinus namay- cush). Genome, 38: 487~496.
한국산 멸종위기 담수어의 세포유전학 및 분자생물학적 연구
I.
배가사리, Microphysogobio long
idorsalis Mori (Cyprinidae: Gobioninae)임재현∙이완옥1∙Lei Peng∙노재구2∙남윤권∙김동수*
부경대학교 양식학과, 1국립수산과학원 내수면생태연구소,
2국립수산과학원 어류육종연구센터
배가사리, Microphysogobio longidorsalis는 현재 서식지의 감소와 환경 오염으로 인하여 멸종 위기에 처해있는 한국 고유의 담수어종이다. 본 연구에서는 종 보존과 자원증대 차원에서 세포유 전학적 및 분자생물학적인 연구를 통해 배가사리의 유전학적인 정보를 밝히고자 하였다. 총 122 개의 염색체 중기상을 관찰 결과, 배가사리의 염색체 수는 2n = 50개 이었으며9쌍의 중부염색체 와 13쌍의 차중부염색체 및 3쌍의 말단염색체가 확인되었다. NORs banding을 실시하여 활성을 갖는rDNA coding 부위를 조사한 결과 한쌍의 차중부염색체에서 명확히 확인이 가능한NORs 부 위가 확인되었다. 또한, f low cytometry를 사용하여DNA 함량을 조사한 결과2.6 pg/cell로 확인되 었다. Cytochrome b전체 염기서열과 623 염기쌍의 12S ribosomal RNA 유전자 단편에 대한 서열 을 분석하여 이를 잉어, 붕어 및 대서양 연어의 염기서열과 비교 분석한 결과, 배가사리와 잉어 및 붕어간 유연관계를 확인할 수 있었다.
Received: May 31, 2004 Accepted: August 29, 2004