한수지 53(1), 90-97, 2020
90
Copyright © 2020 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
Korean J Fish Aquat Sci 53(1),90-97,2020
Original Article
서 론
해양동물의생물학적최소형
,
성비,
생식세포의분화,
생식소 발달,
성성숙및생식주기등에관한연구는종의생식생물학적 이해와생물자원의보존관리및양식기술개발에필요한정보 를제공하게된다.
생식세포의분화과정에관한정보는이러한 연구에필요한기초자료를제공하는매우중요한과정의하나 이다.
배우자형성과정은생식세포의분화,
발달과정에관한것 이지만,
생식소발달과정은생식소내에우점하는생식세포들 을기준으로구분하는것으로배우자형성과정과생식소발달과 정은서로다른개념이다.
전복류를비롯한패류의난모세포는 제1
감수분열말기에체외로산란된다.
따라서난자형성과정은 감수분열단계에따른구분이아니라난원세포단계부터제1
난모세포의형태및조직학적구분에의한것이다
(Sobhon et al., 1999; Gosling, 2004; Gurney and Mundy, 2004).
전복과의난 자형성에관한연구로는Haliotis tuberculata coccinea (Bilbao et al., 2010)
와H. midae (Roux et al., 2013)
의난모세포발달을 비롯한연구가있으나난자형성과정동안난모세포의미세구조 적변화를보고한연구는그수가부족하다.
전복과
(Haliotidae)
는 복족강(Gastropoda),
원시복족목(Ve- tigastropoda)
에 속하며,
현재62
종24
아종이 보고되어 있다(WoRMS, 2018).
국내에분포하는전복과는둥근전복속(Hali- otis)
의2
종2
아종과오분자기속(Sulculus)
의2
아종이보고되어 있다.
이가운데북방전복H. discus hannai
은국내연안의수심5-10 m
조하대에서서식하는각장10 cm
의중형전복으로한 국,
중국및일본등지에서양식및자원조성등으로이용되는북방전복(Haliotis discus hannai) 난모세포의 미세구조적 분화
김혜진·김현진
1·김영숙
2·이정식
1*
국립수산과학원 남동해수산연구소, 1전남대학교 수산생명의학과, 2국립수산과학원 수산방역과
Microstructural Differentiation of the Oocyte in the Abalone Haliotis discus hannai
Hyejin Kim, Hyeon Jin Kim
1
, Young Sook Kim2
and Jung Sick Lee1
*Southeast Sea Fisheries Research Institute, National Institute of Fisheries Science, Tongyeong 53017, Korea
1Department of Aqualife Medicine, Chonnam National University, Yeosu 59626, Korea
2Pathology Division, National Institute of Fisheries Science, Busan 46083, Korea
The purpose of this study was to provide basic information on sexual maturity and reproductive biology for the management of biological resources in abalone Haliotis discus hannai . The nucleus of the oogonium occupied about 42% of the cytoplasm, and had a distinctive basophilic chromatin. The cytoplasm of previtellogenic oocytes was homogeneous and the size of nuclear pores increased. Fine granular and vacuolar yolk granules were observed in the cytoplasm of the initial vitellogenic oocyte. In this stage, the egg stalk and jelly membrane began to develop.
The nucleus of the active vitellogenic oocyte was located near the animal pole. Yolk granules were strongly acido- philic. Lampbrush chromosomes were observed in the nucleus and rough endoplasmic reticulum. Annulate lamellae developed in the cytoplasm. The shape of the ripe oocyte was rounded polygonal. The size of ripe oocytes was 202.9±21.40×142.1±18.82 μm and the thickness of the jelly membrane was 10.1±1.52 μm. These results show that yolk accumulation in H. discus hannai is based on two methods: exogenous accumulation, through the egg stalk, and endogenous accumulation, through intracellular organelles. Management of biological resources will be necessary when oocytes predominate after the active vitellogenic stage.
Keywords: Haliotis discus hannai , Oocyte, Microstructural differentiation
*Corresponding author: Tel: +82. 61. 659. 7172 Fax: +82. 61. 659. 7179 E-mail address: [email protected]
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received 15 January 2020; Revised 24 January 2020; Accepted 10 February 2020 저자 직위: 김혜진(연구사), 김현진(대학원생), 김영숙(연구원), 이정식(교수) https://doi.org/10.5657/KFAS.2020.0090
Korean J Fish Aquat Sci 53(1), 90-97, February 2020
북방전복 난모세포의 미세구조적 변화
91
중요한생물자원가운데하나이다
(Lee et al., 2015).
본연구는 북방전복난자형성과정중난모세포의미세구조적변화를기재 하고이를통해생물자원관리에필요한성성숙및생식생물학 적기초정보를제공하고하고자하였다.
재료 및 방법
재료
본연구에서는생물학적최소형이상
(
각장69.0±12.80 mm)
의북방전복52
개체를분석하였다.
광학현미경 표본 제작
광학현미경조직표본제작은생식소가포함된간췌장의일부 를절취하여
10%
중성포르말린에24
시간동안고정하였다.
고 정된시료는흐르는물에48
시간동안수세한후ethanol
로단계 별탈수를진행하여paraplast (Leica, Wetzlar, Germany)
에포 매하였다.
포매된시료는microtome (RM2235, Leica, Wetzlar, Germany)
을이용하여4-6 μm
두께로연속절편한후유리슬라 이드에부착하였다.
제작된표본은Mayer’s hematoxylin-eosin (H-E)
염색, Masson
삼중염색그리고alcian blue-periodic acid and Schiff’s solution (AB-PAS, pH 2.5)
반응후광학현미경으 로분석하였다.
투과전자현미경 표본 제작
투과전자현미경분석용시료제작은해부단계에서생식소의 일부를절취하여
0.1 M phosphate butter (pH 7.4)
로완충된2.5% glutaraldehyde
용액에2-3
시간고정하였다.
고정된시료 는0.1 M phosphate butter (pH 7.4)
에20
분씩3
회수세한후, 2% osmium tetroxide
로2
시간동안4°C
암조건에서후고정하 였다.
후고정된시료를0.1 M phosphate butter (pH 7.4)
로20
분씩3
회수세한후, ethanol
단계별탈수과정을진행하였다.
그리고30
분간격으로propylene oxide
의치환과정을2
회거 쳐epoxy resin
으로60°C
오븐에서36-48
시간동안중합하였 다.
완성된epoxy block
은0.7-1 μm
두께로semithin
절편후ultramicrotome (EM-UC 7, Leica, Wetzlar, Germany)
을이용하여
600-900 nm
두께로절편한후, uranyl acetate-lead citrate
의이중염색과정을거쳐투과전자현미경(LIBRA
Ⓡ120, Carl Zeiss NTS GmbH, Oberkochen, Germany)
으로관찰하였다. 현미경 화상분석
조직학적특징의정량화를위해현미경화상장치
(KY-F1030,
JVCKENWOOD, Yokohama, Japan) (Moticam 10, Motic Electric Group Co., Ltd, Xiamen, China)
로광학현미경조직표본을
JPEG
파일화하였다.
난모세포의조직학적특징을각항목별로 화상분석프로그램
(i-solution, IMT i-Solution Inc., NY, USA)
을이용하여분석하였다(Fig. 1).
Proportion of nucleus to cytoplasm (%)=
Nucleus area (μm
2) Ooplasm area (μm
2) ×100
조직화학적반응후염색친화도판정은광학현미경조직표본
JPEG
파일을Adobe Photoshop CS6 (Adobe, San Jose, CA, USA)
를통해RGB
방식의색을확인하였다.
이를Code Beau- tify (www.codebeautify.org)
에서Pantone
ⓇFormula Guide/
solid coated (Pantone Inc., Carlstadt, NJ, USA)
의유사색상과 대조하여그고유번호를괄호안에표시하였다.
결 과
북방전복의난소는생식소외막과간췌장사이의생식소내강 에형성된격벽에의해다수의내강으로구분되어있었으며
,
내 강의형태는소엽형으로난자형성소낭을따라난모세포들이발 달되어있었다.
난모세포의분화가진행되면서난모세포,
핵및 인의크기가증가하였고(Fig. 2)
세포질의염색성은호염기성에 서호산성으로전환되는경향을보였다.
난모세포분화과정은 생식세포의크기,
염색성및세포소기관의광학및투과전자현 Fig. 1. Microscopic image analysis of the oocyte in abalone Hali-otis discus hannai. A, oocyte diameter; B, thickness of outer jelly
membrane. Fig. 2. Changes of the diameter and thickness of outer jelly mem-
brane of the oocyte during oogenesis in abalone Haliotis discus hannai. Avo, active vitellogenic oocyte; Ivo, initial vitellogenic oo- cyte; Og, oogonium; Pvo, previtellogenic oocyte; Ro, ripe oocyte.
Vertical error bars indicate SD.
김혜진
ㆍ
김현진ㆍ
김영숙ㆍ
이정식92
미경적특징에따라난원세포
(oogonium),
난황형성전기난모세포
(previtellogenic oocyte),
난황형성개시기난모세포(initial vitellogenic oocyte),
난황형성활성기난모세포(active vitello- genic oocyte)
및완숙기난모세포(ripe oocyte)
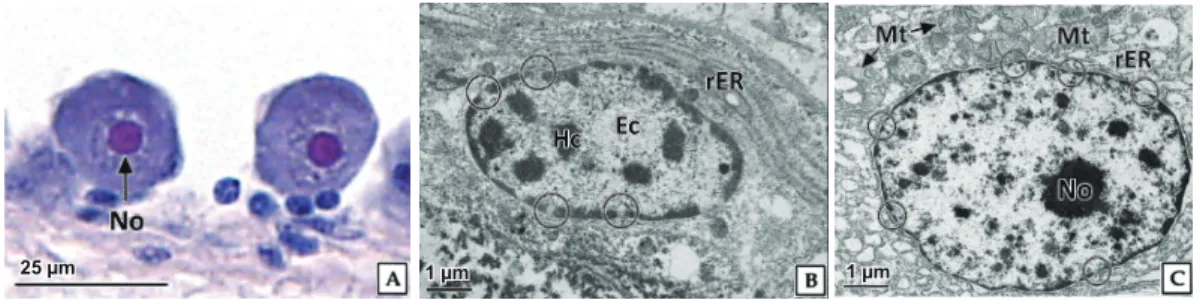
로구분되었다. 난원세포기
난원세포의 형태는 원형으로 직경
10.7±1.90×8.0±1.21 μm
이었다(Fig. 2).
핵은원형으로직경5.8±0.79 μm
로세포질 의약42%
를차지하였다.
난원세포는H-E
염색, Masson
삼중 염색및AB-PAS (pH 2.5)
반응에서강한호염기성을나타내는염색사가핵질에분포함을확인할수있었다
.
세포질은H-E
염색에서는매우약한호염기성을나타냈으며
, Masson
삼중염색 에서옅은붉은색(7430C)
을나타냈으나AB-PAS (pH 2.5)
반 응에서는옅은푸른색(7445C)
으로반응하였다(Fig. 3A).
투과 전자현미경관찰결과,
난원세포의핵질은이형염색질과진정 염색질이뚜렷하게구분되었다(Fig. 3B).
핵막의외막과내막 사이에핵주변공간(perinuclear space)
의거리는약50 nm
이 었으며,
핵공이뚜렷하게구분되었다.
핵의외막을따라리보솜 이부착된조면소포체가발달되어있었다.
세포질에서는크리스테가발달된직경약
300-500 nm
의미토콘드리아를관찰할수있었다
(Fig. 3C).
난황형성전기
난황형성전기의 난모세포는 원형 또는 장방형으로 크기는
15.8±2.21×15.1±2.13 μm
로난원세포에비해약1.5
배증가 하였다(Fig. 2).
핵의크기는8.9±1.32 μm
로난원세포에비해 약65%
증가하였으나세포질에대한핵의비율은27.1±1.70%
로난원세포단계에비해약
40%
감소하였다. H-E
염색결과,
난황형성전기난모세포의핵질에서는강한호염기성을나타내 는원형의인이관찰되었으며,
핵질에는호염기성의염색사들 이산재하고있었다.
세포질은균질한상태였고난원세포단계 에비해H-E
염색과AB-PAS (pH 2.5)
반응에서강한호염기 성으로나타났으며, Masson
삼중염색에서는적자색(682C)
을나타냈다
(Fig. 4A).
핵질에는전자밀도가높은과립상의이형염색질이분포하였으며
,
난원세포단계에비해핵공의크기가 증가하였다.
세포질에는크리스테가발달한다수의미토콘드 리아와시스터네가잘발달된조면소포체가산재하고있었다(Fig. 4B, 4C).
난황형성개시기
난황형성개시기 난모세포는 다각형 또는 장방형으로 난황 형성전기의난모세포에비해다소신장된형태였다
.
난모세포 의크기는61.6±7.10×38.0±6.84 μm
이었으며,
핵의크기는25.8±5.66 μm
로난황형성전기난모세포에비해약3
배증가하였다
(Fig. 2).
초기난황형성개시기난모세포의핵질에는강한호염기성을나타내는원형의인과염색사들이산재하고있 었다
.
세포질의염색성은난황형성전기난모세포에비해호염 기성이다소감소하였으며,
세포질에서는미세과립형태의난 황과립들과 소형의공포들이관찰되었다(Fig. 5A).
세포질에 분포하는난황과립은H-E
염색에서는약한호산성과공포상으 로나타나는2
종류였으며,
난자형성과정이진행되면서난자형 성소낭과난모세포를연결하는난병구조가발달하기시작하였다
(Fig. 5B).
난병부위의세포질에서는소형의공포와호산성의난황과립들이축적되어있었으며
, AB-PAS (pH 2.5)
반응에 서진한푸른색으로반응하는젤리층이확인되었다(Fig. 5C).
핵질에서는전자밀도가높은이형염색질의응집이뚜렷하였으 며
,
세포질에는조면소포체가발달해있었다(Fig. 5D).
세포질 에는관상의미토콘드리아와원형의시스터네구조가잘발달 된골지체다발이분포하고있었다.
난황과립은전자밀도가높 은단백질성과립과투명한지질성과립의두종류가구분되었 는데,
단백질성과립은가장자리와중앙의전자밀도의차이를 보였다(Fig. 5E).
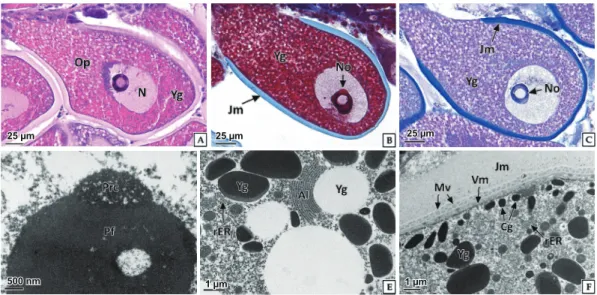
난황형성활성기
난황형성활성기의 난모세포는 난자형성소낭과 난모세포를 연결하는 난병의 구조가명확해지면서난황형성개시기의 난 모세포에비해더욱신장된형태였다
.
난모세포의평균크기는197.9±28.31×80.1±18.82 μm
로측정되었다(Fig. 2).
세포질 Fig. 3. Light and transmission electron microscopy on the oogonia of abalone Haliotis discus hannai. A, note the basophilic chromatin, H-E stain; B, note the euchromatin (Ec) and granular heterochromatin (Hc) in karyoplasm; C, Note the numerous mitochondria (Mt) and rough endoplasmic reticulum (rER) in the cytoplasm; N, nucleus; circle, nuclear pore.25 µm 1 µm 500 nm
25 µm 1 µm 500 nm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
북방전복 난모세포의 미세구조적 변화
93
에대한핵의비율은
15.2±1.96%
이었으며,
핵질을차지하는 인의비율은난황형성개시기난모세포에비해감소하였다.
핵 은동물극부근에위치하며,
핵질에는강한호염기성을나타내 는원형의인이뚜렷하였다.
염색사는이전단계에비해다소감소한양상을보였다
. H-E
염색에서세포질염색성은난황형성개시기에비해약한호염기성에서호산성으로전환되었다
.
세포질에는난황과립과소형의공포들이산재하고 있었으며, H-E
염색에서대부분의난황과립은강한호산성으로나타났 다(Fig 6A).
난황과립들은Masson
삼중염색에서는진한붉은 색(7641C)
으로반응하였으며, AB-PAS (pH 2.5)
반응에서공포상형태와
PAS
에양성으로반응하는난황과립(509C)
의두종류로구분되었다
.
난모세포젤리층외막의두께는4.1±0.92 μm
이었으며,
염색성은H-E
염색에서연한붉은색(2365C)
을 나타냈다.
이들은Masson
삼중염색에서푸른색(271C)
으로반 응하였고(Fig 6B) AB-PAS (pH 2.5)
반응에서alcian blue
에양성으로 강하게 반응하여 진한 푸른색
(2136C)
을 나타냈다(Fig. 6C).
투과전자현미경표본에서난모세포의 핵질에서는솔염색체
(lampbrush chromosome)
가 관찰되었으며,
인의내 부에서는전자밀도가낮은pars fibrosa
가관찰되며,
인의주변 에는망상구조의perinucleolar chromatin
이위치하고있었다 Fig. 4. Light and transmission electron microscopy on the previtellogenic oocytes of abalone Haliotis discus hannai. A, note the basophilic cytoplasm and eosinophilic nucleolus (No), H-E stain; B, note the well developed rough endoplasmic reticulum (rER) in the cytoplasm;C, not the well developed nuclear pore (circle) and electron dense nucleolus in karyoplasm; Ec, euchromatin; Hc, heterochromatin; Mt, mitochondria.
25 µm 1 µm 500 nm
25 µm 1 µm 500 nm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
Fig. 5. Light and transmission electron microscopy on the initial vitellogenic oocyte of abalone Haliotis discus hannai. A, Note the baso- philic cytoplasm and small vacuolar yolk granules, H-E stain; B and C, Note the eosinophilic and PAS positive yolk granules near the egg stalk (Es). AB-PAS (pH 2.5) reaction; D, note the well developed mitochondria (Mt) and rough endoplasmic reticulum (rER); E, two types of yolk granules (Yg) and Golgi complex (Gc) with well developed cisternae; Ec, euchromatin; Hc, heterochromatin; Jm, outer jelly mem- brane; N, nucleus; No, nucleolus; circle, nuclear pore.
25 µm 1 µm 500 nm
25 µm 1 µm 500 nm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
김혜진
ㆍ
김현진ㆍ
김영숙ㆍ
이정식94
Fig. 6. Light and transmission electron microscopy on the active vitellogenic oocyte of abalone Haliotis discus hannai. A-C, note the nucleus (N) in the animal pole; A, H-E stain; B, Masson’s trichrome stain; C, AB-PAS (pH 2.5) reaction; D, network structure of perinucleolar chromatin (Prc) and electron lucent pars fibrosa (Pf); E, two types of yolk granules (Yg) and well developed annulate lamella (Al) in the ooplasm (Op); F, cortical granules (Cg) under the vitellin membrane (Vm) and developed outer jelly membrane (Jm); Mv, microvilli; rER, rough endoplasmic reticulum.
25 µm 1 µm 500 nm
25 µm 1 µm 500 nm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
Fig. 7. Light and transmission electron microscopy on the ripe oocyte of abalone Haliotis discus hannai. A-C, note the well developed outer jelly membrane (Jm); A, H-E stain; B, Masson’s trichrome stain; C, AB-PAS (pH 2.5) reaction; D, annulate lamella (Al) with electron lu- cent yolk granule in the ooplasm (Op); E, Electron lucent karyoplasm (Kp) and numerous nuclear pore (circle); F, Microvilli (Mv) between vitellin membrane (Vm) and outer jelly membrane (Jm); Cg, cortical granule; N, nucleus; Ne, nuclear envelope; No, nucleolus; rER, rough endoplasmic reticulum; Yg, yolk granules.
25 µm 1 µm 500 nm
25 µm 1 µm 500 nm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
25 µm 1 µm 1 µm
25 µm 25 µm 25 µm
500 nm 1 µm 1 µm
(Fig. 6D).
난황과립사이의세포질에서는조면소포체와환층판
(annulate lamellae)
이발달되어있었으며,
환층판은지질성 난황과립을둘러싸고있었다(Fig. 6E).
또한젤리층외막의내부에는미세융모가발달되어있었으며
,
난황막아래의세포질 에는전자밀도가높은피질과립(cortical granule)
들이산재하 고있었다(Fig. 6F).
북방전복 난모세포의 미세구조적 변화
95
완숙기
완숙기 난모세포의 형태는 둥근 다각형이며
,
평균 크기는202.9±21.40×142.1±18.82 μm
로 난황형성활성기 난모세 포에비해단경이약56%
증가하였다(Fig. 2).
염색사는난황 형성활성기에비해감소하였으며,
세포질에는난황과립과소 형의공포가세포질전체에분포하고있었다.
난황과립은H-E
염색에서 강한 호산성의 붉은색
(224C)
으로 나타났으며(Fig.
7A), Masson
삼중염색에서붉은색(200C) (Fig.7B), AB-PAS (pH 2.5)
반응에서는푸른색으로반응하였다(Fig. 7C).
성숙기 난모세포의젤리층외막의두께는10.1±1.52 μm
로난황형성 활성기난모세포에비해약2
배이상증가하였다.
투과전자현 미경표본에서완숙기난모세포의핵질에는솔염색체가여전 히발달되어있었고세포질은전자밀도가높은단백질성의난 황과립과투명하게관찰되는지질성분의난황과립들로채워져 있었으며,
원형의환층판이발달되어있었다(Fig. 7D).
완숙기 난모세포에서핵공의크기는난황형성활성기에비해감소되었 고젤리층외막에서는발달된미세융모들을관찰할수있었으 며(Fig. 7E),
난황막아래의세포질에서는직경약250 nm
의피 질과립들이분포하고있었다(Fig. 7F).
고 찰
전복과를포함한대부분의복족류의성은자웅이체로난소는 다수의난자형성소낭으로 이루어져있으며
,
난자형성소낭내 에서난모세포의분화및발달이진행된다.
난모세포들의발달 은각소낭내에서비동시적으로진행되며,
계절에따른변화양 상을나타낸다(Voltzow, 1994; Jung et al., 2007).
북방전복의 난소또한다수의난자형성소낭으로이루어져있었으며,
각소 낭을따라난모세포들이발달하고있었다.
난자형성소낭은결 합조직으로이루어진격벽이생식소외막의안쪽에서부터간 췌장까지발달되어있는소엽형구조를가지고있었다.
이러한 난소의구조는H. rufescens (Martin et al., 1983), H. iris
와H.
australis (Wilson and Schiel, 1995)
를비롯한다른전복류와동 일하였다(Sobhon et al., 1999; Najmudeen and Victor, 2004;
Bilbao et al., 2010; Visser-Roux, 2011).
패류에서난모세포의분화
,
발달과정동안뚜렷한특징중하 나는난모세포의크기증가로이는세포질내난황물질의축 적에따른결과이다.
둥근전복속(Haliotis)
의완숙난모세포크 기는H. rufescens 250 μm (Giorgi and DeMartini 1977), H.
asinina 125 μm (Capinpin et al., 1998), H. asinina 80-140 μm (Sobhon et al., 1999), H. varis 180±20 μm (Najmudeen and Victor, 2004), H. midae 250-300 μm (Visser-Roux, 2011; Wu et al., 2018)
으로종에따라다르게보고되고있다.
본연구에서 북방전복의완숙난모세포의크기는장경약202.9±21.4 μm
였는데,
이와같은완숙난모세포의크기차이는난황축적정도 에따른것으로판단되지만,
이러한차이가종특이성인지생태학적특성인지에대해서는추후의연구가필요하다
.
난모세포의난황축적방법은난황전구물질의기원부위에따 라외재적난황축적과내재적난황축적으로구분된다
.
난모세 포발달과정에서난황축적과관련있는구조가운데난병은난 모세포의외재적난황축적과밀접한관계가있으며,
인을포함 한세포소기관들은난모세포의내재적난황축적과밀접한관계 가있다(Beams and Sekhon, 1966; Wallace and Selmen, 1981;
Eckelbarger and Young, 1997).
복족류와이매패류에서난병은미세소관들이밀집된구조로 난소형성소낭과난모세포의부착및지지기능과동시에난황 전구물질의통로기능을담당한다
(Beams and Sekhon, 1966;
Park et al., 2003; Jung et al., 2007).
북방전복에서도난병은난 황형성개시기의난모세포시기부터구분되기시작하여난모세 포가발달됨에따라더욱뚜렷해졌다.
또한동일한시기부터난 병부위의난세포질에서H-E
염색에서호산성으로반응하고AB-PAS (pH 2.5)
반응에서는PAS
에양성으로반응하는난황 과립들이확인되었다.
내재적난황축적은기본적으로세포내물질합성기작에의한 것으로핵내의유전자의활성으로부터시작되어인과조면소 포체및골지체등의세포내소기관에의해주로이루어진다
(Wallace and Selman, 1981; Visser-Roux, 2011).
원시복족목 패류에서난모세포의난황축적은비대성여포세포와관계가있으며
, 1)
조면소포체와골지체의결합작용을통한세포질내의자가합성 형태와
2)
명확하지는않지만미토콘드리아와같은 다양한세포소기관에의한방법, 3)
음세포작용등을통한타가 합성의세가지난황축적방법이보고되었다(Eckelbarger and Young, 1997).
뿔소라과의
Bolinus brandaris
의경우에도난황축적방법은 여포세포에의한외재적방법과조면소포체등에의한내재적 방법에의한다(Amor et al., 2004).
소라(Batillus cornutus)
의 난황축적과정에서도난병의발달과인과세포소기관의활성이 관찰되어소라의난황축적도외재적방법과내재적난황축적 의두가지방법에의한것으로보고되었다(Jung et al., 2007).
본연구에서도북방전복의난모세포발달과정이진행됨에따 라핵질에서솔염색체의활성화와세포질내에서조면소포체
,
환층판및골지체등의세포소기관의발달이확인되었다.
이러 한결과는북방전복의난황축적은난병을통한외재적축적과 세포내소기관을통한내재적축적의두가지방법에의한것임 을보여주는것이다.
패류에서난모세포의분화
,
발달과정동안핵의이동,
피질과 립및젤리층의발달은수정과산란후환경적응을위한구조 적분화이다.
성숙기 난모세포의 조직학적 특징인 핵의 동물극
(animal
pole)
으로의 이동은 수정 시 웅성전핵과의 결합을 용이하게하기 위한것이며
,
피질과립의발달은수정막형성에관여한 다(Wallace and Selmen, 1981).
이와같은현상은H. midae
김혜진
ㆍ
김현진ㆍ
김영숙ㆍ
이정식96
(Wood and Buxton, 1996)
와H. asinina (Jebreen et al., 2000)
에서보고되었으며,
본연구의북방전복에서도확인되었다.
체외수정을위해수중에산란을하게되는무척추동물의경우 다양한물리화학적외부환경요인으로부터난을보호하기위 해젤리층과같은다양한난막구조를발달시킨다
(Bolton et al., 2000; Beninger and Chérel, 2019).
복족류인Tegula funebralis
에서성숙난모세포는두께250-260 μm
의젤리층으로둘러싸 여있으며(Moran, 1997),
소라의완숙난모세포에서젤리층의 두께는약5.3 μm
이며(Jung et al., 2007), H. asinina
의완숙난 모세포에서젤리층외막의발달이보고되었다(Wetakan et al., 2001).
북방전복의난모세포에서젤리층외막은난황형성활성기단 계부터급격히발달하여완숙기난모세포에서는두께약
10 μm
로분석되었으며,
조직화학적으로젤리층은산성의당단백질 로구성된것으로확인되었다.
한국에서북방전복의주산란기 는9
월전후로보고되었지만(Kim et al., 2016),
전복류의주산 란기는수온을비롯한환경조건에따른성성숙속도및생식 소발달단계의차이에의해변화가가능하다(Uki and Kikuchi, 1984; Grubert and Ritar. 2004; Najmudeen and Victor, 2004).
따라서북방전복은세포학적측면에서난황형성활성기이후의 난모세포들이우점하는시기에남획을최소화하는금어기등과 같은생물자원의관리가필요할것으로판단된다
.
사 사
본연구는해양수산부농림수산식품기술기획평가원
(Golden Seed
프로젝트, 213008-05-4-SB720)
의지원에의해수행되었 습니다.
References
Amor MJ, Ramo M and Durfort M. 2004. Ultrastructural stud- ies of oogenesis in Bolinus brandaris (Gastropoda: Mu- ricidae). Sci Mar 68, 343-353. https://doi.org/10.3989/
scimar.2004.68n3343.
Beams HW and Sekhon SS. 1966. Electron microscope studies on the oocyte of the fresh-water mussel (Anodonta), with special reference to the stalk and mechanism of yolk de- position. J Morphol 119, 477-501. https://doi.org/10.1002/
jmor.1051190407.
Beninger PG and Chérel D. 2019. Cloaked bivalve oocytes: les- sons in evolution, ecology, and scientific awareness. Ecol- ogy https://doi.org/10.1002/ecy.2818.
Bilbao A, Tuset V, Viera M, De Vicose GC, Fernández-Palacios H, Haroun R and Izquierdo M. 2010. Reproduction, fecun- dity, and growth of abalone (Haliotis tuberculata coccinea, Reeve 1846) in the Canary Islands. J Shellfish Res 29, 959- 967. https://doi.org/10.2983/035.029.0430.
Bolton TF, Thomas FIM and Leonard CN. 2000. Maternal en-
ergy investment in eggs and jelly coats surrounding eggs of the echinoid Arbacia punctulata. Biol Bull 199, 1-5. https://
doi.org/10.2307/1542700.
Capinpin Jr EC, Encena II VC and Bayona NC. 1998. Studies on the reproductive biology of the Donkey's ear abalone,
Haliotis asinina Linne. Aquaculture 166, 141-150. https://
doi.org/10.1016/S0044-8486(98)00275-0.
Ecklebarger KJ and Young CM. 1997. Ultrastructure of the ovary and oogenesis in the methane-seep mollusc
Bathynerita naticoidea (Gastropoda: Neritidae) from the
Louisiana slope. Invertebr Biol 116, 299-312. https://doi.org/10.2307/3226862.
Giorgi AE and DeMartini JD. 1977. A study of the reproductive biology of the red abalone, Haliotis rufescens Swainson, near Mendocino, California. Calif Fish Game 63, 80-94.
Gosling E. 2004. Bivalve molluscs: Biology, ecology and cul- ture. Blackwell science, Oxford, U.K., 1-443.
Grubert MA and Ritar AJ. 2004. Temperature effects on the dy- namics of gonad and oocyte development in captive wild- caught blacklip (Haliotis rubra) and greenlip (H. laevigata) abalone. Invertebr Reprod Dev 45, 185-196. https://doi.org/
10.1080/07924259.2004.9652590.
Gurney LJ and Mundy CN. 2004. Reproductive biology: iden- tifying spawning cycles in the genus Haliotis. In: Technical Report Series. Tasmanian Aquaculture Fisheries Institute 23, 1-27.
Jebreen E, Counihan T, Fieldder D and Degnan B. 2000. Syn- chronous oogenesis during the semilunar spawning cycle of the tropical abalone Haliotis asinina. J Shellfish Res 19, 845-851.
Jung GK, Park JJ, Ju SM, Jin YG and Lee JS. 2007. Ovarian structure and oogenesis of the spiny top shell, Batillus cor-
nutus (Lightfoot, 1786) (Gastropoda: Turbinidae). Korean J
Malacol 23, 209-216.Kim H, Kim BH, Son MH, Jeon MA, Lee YG and Lee JS. 2016.
Gonadal development and reproductive cycle of aquaculture abalone, Haliotis discus hannai in Korea (Gastropoda: Hali- otidae): Implications for seed production. J Shellfish Res 35, 653-659. https://doi.org/10.2983/035.035.0311.
Lee JS, Son MH, Lee JS, Lim HK, Won SH, Lee JH, Kang JC, Kim JW and Kim SY. 2015. Korean abalone (I): morphol- ogy, taxonomy and ecology. Maple Design, Busan, Korea, 1-68.
Martin GG, Romero K and Miller-Walker C. 1983. Fine struc- ture of the ovary in the red abalone Haliotis rufescens (Mol- lusca: Gastropoda). Zoomorphology 103, 89-102. https://
doi.org/10.1007/BF00312240.
Moran AL. 1997. Spawning and larval development of the black turban snail Tegula funebralis (Prosobranchia: Tro- chidae). J Mar Biol 128, 107-114. https://doi.org/10.1007/
s002270050074.
Najmudeen TM and Victor ACC. 2004. Reproductive biology
북방전복 난모세포의 미세구조적 변화
97
of the tropical abalone Haliotis varia from Gulf of Mannar.
J Mar Biol Ass India 46, 154-161.
Park JJ, Lee JY, Lee JS and Chang YJ. 2003. Gonadal de- velopment and gametogenic cycle of the equilateral ve- nus, Gomphina veneriformis (Bivalvia: Veneridae). Ko- rean J Fish Aqua Sci 36, 352-357. https://doi.org/10.5657/
kfas.2003.36.4.352.
Roux A, Lambrechts H and Roodt-Wilding R. 2013. Reproduc- tive histology of cultured Haliotis midae (Linnaeus, 1758) and preliminary evaluation of maturation. J Shellfish Res 32, 143-153.
Sobhon P, Wetakan SA, Chanpoo M, Wanichanon C, Linthong V, Thongkukiatkul A, Jarayabhand P, Kruatrachue M, Upatham SE and Poomthong T. 1999. Classification of germ cells, reproductive cycle and maturation of gonads in
Haliotis asinina Linnaeus. Sci Asia 25, 3-21.
Uki N and Kikuchi S. 1984. Regulation of maturation and spawning of an abalone, Haliotis (Gastropoda) by external environmental factors. Aquaculture 39, 247-261. https://doi.
org/10.1016/0044-8486(84)90270-9.
Visser-Roux A. 2011. Reproduction of the South African aba- lone, Haliotis midae. Ph.D. Dissertation, University of Stel- lenbosch, Stellenbosch, Republic of South Africa.
Voltzow J. 1994. Gastropoda: Prosobranchia. In: Microscopic anatomy of invertebrates vol. 5. Mollusca I. Harrison FW and Kohn AJ, eds. Wiley-Liss, New York, NY, U.S.A., 111- Wallace RA and Selmen K. 1981. Cellular and dynamic aspects 252.
of oocyte growth in teleosts. Am Zool 21, 325-343. https://
doi.org/10.1093/icb/21.2.325.
Wetakan SA, Linthong V, Wanichanon C, Panasophonkul S, Meepool A, Kruatrachue M, Upatham ES, Pumthong T and Sobhon P. 2001. Ultrastructure of female germ cells in
Haliotis asinina Linnaeus. Invertbr Reprod Dev 39, 67-79.
https://doi.org/10.1080/07924259.2001.9652468.
Wilson NHF and Schiel DR. 1995. Reproduction in two spe- cies of abalone (Haliotis iris and H. australis) in southern New Zealand. Mar Freshwater Res 46, 629-637. https://doi.
org/10.1071/MF9950629.
Wood AD and Buxton CD. 1996. Aspects of the biology of the abalone Haliotis midae (Linne, 1758) on the east coast of South Africa. 2. Reproduction. S Afr J Mar Sci 17, 69-78.
https://doi.org/10.2989/025776196784158356.
WoRMS (World Register of Marine Species). 2018. Taxonomic status of family haliotidae. Available at: http://www.marine- species.org/aphia.php?p= taxdetails&id=1727 on Oct 7, 2019.
Wu Y, Kaiser H and Jones CLW. 2018. A first study on the ef- fect of dietary soya levels and crystalline isoflavones on growth, gonad development and gonad histology of farmed abalone, Haliotis midae. Aquac Int 27, 167-193. https://doi.
org/10.1007/s10499-018-0315-6.