126 Corresponding author: [email protected]✽
돌돔 (
Oplegnathus fasciatus
)으로부터 Metallothionein 유전자 Isoform들의 분리 및발현 특징 분석
이상윤・남윤권✽
부경대학교 해양바이오신소재학과
Isolation and mRNA Expression of Metallothionein Isoforms from Rockbream Oplegnathus fasciatus
Sang Yoon Lee and Yoon Kwon Nam*
Department of Marine Bio-Materials and Aquaculture, Pukyong National University, Busan 608-737, Korea
Genetic determinants of two metallothionein isoforms (MT-A and MT-B) were isolated and characterized from the perciform species, rockbream (Oplegnathus fasciatus). Rockbream MT-A and MT-B shared a high degree of homology at amino acid levels with representative orthologs from other perciform species, especially with respect to the conserved cysteine residues. At the genomic level, both MT-A and MT-B genes represent a tripartite structure typical of vertebrate MT genes. However, rockbream MT-B showed unusually large introns (1.2 kb and 0.8 kb for intron I and II, respectively), a phenomenon that has rarely been seen in other vertebrate MT genes. MT-A and MT-B transcripts were ubiquitously detected in a wide array of tissues, wherein brain and eye showed the highest basal expression levels, and the fin exhibited the lowest expression of both isoforms. The basal expression of MT-A in most tissues was significantly higher (ranging from 4- to 10-fold) than that of MT-B. Upon heavy metal exposures to Cd, Cu or Zn at 25 ppb for 48 h, MT-A and MT-B transcripts in the liver were significantly activated by Cd and moderately by Zn. On the other hand, exposure to Cu did not result in alterations of MT-A, nor in the significant suppression of MT-B. Following bacterial challenges with Escherichia coli, Edwardsiella tarda or Streptococcus iniae, MT isoforms in the liver, kidney and spleen were highly modulated and exhibited a pattern that was dependent on the bacterial species, tissues and isoforms. These results suggest that the two MT isoforms could be taken into account as potential indicators of metal toxicity and immune perturbations of this aquaculture-relevant species.
Key words: Gene expression, Genomic structure, Metallothionein, Rockbream (Oplegnathus fasciatus)
서 론
Metallothionein (MT)은 6~7 kDa의 저분자 단백질으로서 60개 내외의 아미노산으로 구성되어 있으며, 특히 시스테인 (cysteine)함량이 높고 시스테인 잔기들의 위치가 대부분 척추 동물에서 잘 보전되어 있는 특징을 가지고 있다 (Coyle et al., 2002; Haq et al., 2003; Cho et al., 2005). MT는 다양한 종류의 중금속들과 결합능력을 보유하고 있으며, 체내 필수 금속 이온들의 저장 및 항상성 유지는 물론 비필수 중금속에 대한 해독 및 생체방어 기능을 담당한다고 알려져 있다 (Wassermann et al., 1992; Ohta et al., 1993). 특히 MT는 체내 유입된 중금속에 의해 신속하게 유도되는 특징을 갖고 있기 때문에 종래 많은 연구들에서 중금속 오염에 대한 생체 분자 표지로서 MT를 활용하고자 한 바 있다 (Tom et al., 2004;
Woo et al., 2006; Knapen et al., 2007; Gao et al., 2009). 뿐만 아니라 최근 여러 연구들에서 MT 유전자가 중금속 해독 기능
외에 다양한 스트레스에도 유의적인 발현 반응을 나타낸다고 보고되고 있으며, 특히 염증, 상처 및 감염 등에 대한 선천성 면역 반응에 밀접히 관여한다고 알려지고 있다 (Jenny et al., 2006; Baerwald et at., 2008; Wang et al., 2009; Chen et al., 2010).
포유류는 일반적으로 4 종류의 MT isoform들을 보유하며 (Palmiter et al., 1992; Coyle et al., 2002; Thirumoorthy et al., 2007), 경골어류는 종에 따라 2~3 종류의 MT isoform을 보유하 는 것으로 보고되어 있다. 포유류에서 밝혀진 MT의 isoform별 다양한 기능들이 어류 MT들에서도 보전되어 있으리라 예상 되지만 아직 어류 MT isoform별 기능의 분화와 자세한 발현 조절 양상에 대해서는 많이 알려져 있지 못한 상태이다. 특히 어류의 MT 유전자 isoform들의 경우 진화 경로가 포유류와는 차이가 있으며, isoform들 간 기능의 분화가 종별로 서로 차이 가 있음이 알려지기 시작하면서 어류 MT를 분자표지로 활용 하기 위해서는 해당 어종별 isoform들의 발현 조절에 대한 이해가 점차 중요시 되고 있다 (Bourdineaud et al., 2006;
Table 1. Oligonucleotide primers used in this study
Primer name Sequence (5'-3') Purpose
RB18S rRNA 1F TAACGGGGAATCAGGGTTCGAT
Real-time RT-PCR of 18S rRNA RB18S rRNA 1R CAAGAATTTCACCTCTAGCGGC
RBMT 1F GACTCAACACACATCACCTG Isolation of MT-A gDNA fragment
RBMT 1R GAAGTAGTTCACACAGGCTC
RBMT-A GW1F CACTGCTTCACCGCTTTGTAACGGTA Genome walking to 3'-downstream region of MT-A genes
RBMT-A GW2F CAGTGTCTTGATTTCAATGTCCTGGA
RBMT-A FW1 CCTATAGCCTGTATTAGCA Confirmation of MT-A gene from the 5'-upstream region to 3'-downstream region
RBMT-A RV1 GAGTCTGAGAGGCAAAATC
RBMT-B GW1F CAAACTGAGCGCATGACTTGTTGATTT Genome walking to 3'-downstream region of MT-B genes
RBMT-B GW2F AGGTCTGAGGGTCTAACTTTCGGCCAA RBMT-B GW3F CATCAACCCTGCAGTTACCTGACTTT RBMT-B GW4F TTCCTTTGTTCCAGTGTCAGCTTGTC
RBMT GW01 TGAGACATGTTAAGCCCACAGTGACA Genome walking to 5'-upstream region of MT-A, B genes
RBMT GW02 AACACTTACTCTTGGAGCAGTCGCAA
RBMT-B FW1 CTGCATCAGGCCCATTCAAA Confirmation of MT-B gene from the 5'-upstream region to 3'-downstream region
RBMT-B RV1 TGAGTCTGAGGGTAGAATCAG
qRBMT-A 1F TCCAAGAGTGGAACCTGCAA Real-time RT-PCR of MT-A transcripts qRBMT-A 1R ACATCATTCCCTATAGACAATTG
qRBMT-B 1F TCCAAGAGTGGAACCTGCAC Real-time RT-PCR of MT-B transcripts / Isolation of MT-B cDNA fragment
qRBMT-B 1R CTTCGTAGACTGCAATTAACTCA
Baerwald et al., 2008). 그럼에도 불구하고 아직 많은 어종들에 서 오직 단일 MT isoform만이 발굴되어 있는 실정이며, 특정 자극 조건하에서 isoform들 간의 조절 양상에 관한 상호 비교 연구는 일부 어종들에서만 이루어진바 있다. 더욱이 현재까지 의 대부분 어류 MT 연구들은 중금속에 대한 반응 조사에 국한되어 있으며 여타 면역자극에 대한 어류 MT 유전자의 조절에 대해서는 그 연구 정보가 매우 부족한 실정이다.
따라서 본 연구는 해산 양식 품종인 돌돔 (Oplegnathus fasciatus)으로부터 MT 유전자 isoform들을 분리하고, 구조적 특징을 분석하며 아울러 실험 자극에 대한 MT 유전자 isoform 들의 발현 반응을 분석하고자 하였다. 이를 위해 돌돔 MT isoform 2종류의 cDNA 및 gDNA를 분리하였으며, 각 isoform의 5'-upstream 영역에 존재하는 전사조절인자들을 조사하였고, 중금속 노출 및 세균 감염에 의한 전사 발현 특징을 분석하였다.
재료 및 방법
실험어 및 cDNA library
본 연구에 사용된 돌돔 개체들은 부산 남천동 활어시장에서 구입하여 유전자원공학 연구실에서 여과해수 (용존산소: 5
± 1 ppm, 수온: 22 ± 1℃)를 이용하여 유지한 개체들을 이용하 였다. MT 유전자 발굴에 이용한 cDNA library는 돌돔의 간 조직으로부터 분리한 mRNA를 이용하여 ZAP-cDNA Synthesis Kit, ZAP-cDNA Gigapack III Gold Cloning Kit
(Stratagene, La Jolla, CA, USA)의 권고 방법에 따라 제작한 cDNA library였다 (Choi, 2005).
돌돔 MT-A isoform 유전자의 분리
돌돔 간 조직 cDNA library로부터 수집한 expressed sequence tag (EST) 단편들을 대상으로 BLAST 검색을 실시하 였으며, 이를 통해 확보한 첫 번째 MT isoform을 MT-A로 명명 하였다. Full-length ORF를 포함하는 genomic 유전자를 확보 하기 위해서 상기 MT-A cDNA의 5'-및 3'-untranslated region (UTR) 영역에서 각각 상보적 결합을 하도록 제작된 RBMT 1F 및 RBMT 1R 프라이머를 이용하여 돌돔 gDNA로부터 MT-A 유전자를 PCR 분리하였다. 본 연구에 사용한 PCR 프라 이머 정보는 Table 1에 나타내었다. Genomic DNA는 지느러 미 조직으로부터 통상의 proteinase K/SDS 방법을 이용하여 추출하였고, 순수 분리한 100 ng을 PCR 반응의 주형으로 사용하였다. PCR은 AccuPower HF PCR premix (Bioneer, Daejeon, Korea)를 이용하여 94℃에서 2분, 94℃에서 45초, 58℃에서 45초, 72℃에서 60초의 30회 순환과정으로 수행하 였고, 증폭된 PCR 산물은 Gel Purification Kit (Bioneer)를 이용하여 제조사의 권고 방법에 따라 정제하였다. 정제 산물 은 pGEM-T Easy Vector System (Promega, Madison, WI, USA) 을 이용하여 제조사의 권고 방법대로 클로닝 한 후 염기서열 을 분석하였다.
MT-A 유전자의 5'-upstream 및 3'-downstream 영역 확보를 목적으로 Universal GenomeWalker Kit (BD Biosciences
Clontech, Mountain View, CA, USA)를 이용 genome walking을 수행하였다. 돌돔 gDNA의 제한 효소 처리, genome walker DNA library의 제작 및 nested PCR 조건은 모두 상기 Kit에 기술된 권고 방법에 따라 수행하였다. 5'-upstream 영역을 확보 하기 위해서 RBMT GW01 및 RBMT GW02 프라이머를 사용 하였고, 반면 3'-downstream 영역은 RBMT-A GW1F 및 RBMT-A GW2F 프라이머를 사용하여 확보하였다. 양방향으 로의 genome walking에 의해서 증폭된 각각의 PCR 산물은 앞서와 마찬가지로 gel purification 및 TA 클로닝 과정을 거쳐 단편들의 염기서열 분석을 실시하였고, 단편 염기서열들의 contig assembly를 통해 연속 단편 염기서열을 확보하였다.
확보한 염기서열을 검증하기 위해서 돌돔 MT-A 유전자의 5'-upstream부터 3'-downstream까지의 영역을 다시 PCR 분리 한 후 대표 염기서열을 확정하였다. PCR 분리는 RBMT-A FW1/RBMT-A RV1 프라이머 쌍을 이용하여 Expand High Fidelity System (Roche Applied Science, Mannheim, Germany) 를 이용하여 실시하였다.
돌돔 MT-B isoform 유전자의 분리
돌돔 MT-B의 genomic 유전자는 앞서 MT-A isoform의 genome walking 과정 중 MT-A와 차등 염기서열을 보이는 클론을 탐색하여, 이로부터 직접 genome walking을 실시하여 확보하였다. MT-A 5'-upstream 영역의 서열과 차이를 나타내 는 위치를 대상으로 3'-downstream 방향으로 2회의 순차적인 genome walking을 수행하였다. 1차 genome walking은 RBMT-B GW1F 및 RBMT-B GW2F 프라이머를 이용하였고 2차 walknig은 RBMT-B GW3F 및 RBMT-B GW4F 프라이머를 이용하였다. 앞서 MT-A 유전자의 클로닝과 마찬가지로 각 증폭단편 서열들의 assembly와 연속 염기서열의 분석을 실시 하였다. 돌돔 MT-B isoform의 cDNA 염기서열을 확보하기 위해서 상기 genomic 서열과 MT-A cDNA서열과의 비교 분석 을 통해서 exon 서열만을 수집하였다. 수집한 MT-B coding region의 서열과 MT-A cDNA 서열간의 비교를 통해서 MT-B cDNA만을 특이적으로 분리할 수 있는 프라이머 (qRBMT-B 1F, qRBMT-B 1R)를 제작하였다. MT-B isoform의 단편 분리를 위해서 상기 두 프라이머를 이용, vectorette PCR를 수행하였 다. Vectorette PCR을 위한 주형 DNA는 돌돔 간 cDNA library 의 excised stock으로부터 제조한 phagemid DNA pool을 이용 하였고, 이때 벡터프라이머는 phagemid 벡터의 multi-cloning site (MCS)에 인식 위치를 갖는 T7 및 SK 프라이머를 사용하였 다. PCR 반응은 qRBMT-B 1F/T7 및 qRBMT-B 1R/SK 프라이 머 쌍을 이용, 독립적으로 실시하였고 두 증폭반응 모두 동일 한 열 순환 조건 (94℃에서 2분, 94℃에서 30초, 58℃에서 30초, 72℃에서 60초의 35회)을 이용하였다. 증폭된 각각의 PCR 산물을 대상으로 앞서와 같이 순수분리, TA클로닝 및 염기서열 분석을 실시하였고, 최종 RBMT-B FW1과 RBMT-B RV1 프라이머를 이용 연속 단편서열을 검증하였다.
생물정보 분석
MT cDNA 염기서열 내 open reading frame (ORF) 영역의
예측 및 추정 아미노산 서열로의 변환은 Open Reading Frame Finder (ORF Finder; http://www.ncbi.nlm.nih.gov/projects/gorf) 를 이용하였다. 돌돔 MT isoform들과 여타 농어목 어류 MT ortholog들과의 아미노산 서열의 비교 분석을 위하여 CLUSTALW (http://align.genome.jp)를 이용, 다중서열배치 (multiple sequence alignment)를 실시하였다. 또한 단백질 분자 량 및 theoretical pI값을 구하기 위하여 ProtParam를 이용하였 다 (Gasteiger et al., 2005). 반면 5'-upstream에 존재하는 전사조 절 인자 부착 위치를 예측하기 위해서 Transcription Element Search System (TESS; www.cbil.upenn.edu/cgi-bin/tess/tess)와 Searching Transcription Factor Binding Sites (ver 1.3) (TFSEARCH; http://www.cbrc.jp/research/db/ TFSEARCH.html) 을 이용한 분석을 실시하였다.
MT isoform들의 mRNA 조직 발현 분석
돌돔 MT의 mRNA 조직 분포를 비교하기 위하여 돌돔 (평균 어체중 = 124 ± 16 g) 18마리를 대상으로 10종류의 장기 조직 (뇌, 눈, 지느러미, 아가미, 심장, 소화관, 신장, 간, 근육, 비장) 을 적출하였다. 적출한 돌돔의 각 조직으로부터 total RNA 분리 및 cDNA 합성을 수행하였다. 10종류의 조직 가운데 아가미, 소화관, 신장, 간, 비장 조직은 RNeasy Mini Kit (Qiagen, Hilden, Germany)와 RNase-Free DNase Set (Qiagen)를 사용하여 제조사의 권고 방법에 따라 total RNA를 추출하였 고, 뇌, 눈, 지느러미, 심장, 근육 조직은 Tripure reagent (Roche) 로 total RNA를 추출한 후 RNeasy Mini Kit (Qiagen)와 RNase-Free DNase Set (Qiagen)를 사용하여 정제과정을 수행 하였다. 총 10종류 조직으로부터 추출한 total RNA는 spectrophotometer (Gene Quant II spectrophotometer; Pharmacia Biotech, Uppsala, Sweden)를 이용한 농도 측정 값과 18S : 28S ribosomal RNA 밴드의 전기영동 상을 고려하여 순도와 양을 평가하였다.
cDNA 합성은 Omniscript RTase Kit (Qiagen)의 권고 방법과 RB18s rRNA 1R 0.1 μM를 첨가하여 역전사 반응을 수행하였 다. 발현 정량 분석을 위하여 수행한 Real-time RT-PCR은 iCycler Real-Time Optic Module (Bio-Rad)과 iQ SYBR Green supermix (Bio-Rad)를 사용하였고, 18S rRNA 증폭에는 RB18S rRNA 1F 및 RB18S rRNA 1R 프라이머를 사용하였으며, MT-A 및 MT-B 증폭에는 qRBMT-A 1F/1R 및 qRBMT-B 1F/1R 프라 이머 쌍을 사용하였다. Real-time RT-PCR은 18S rRNA를 normalization control로 이용하여 조직별 상대 정량을 실시하 였다. Real-time RT-PCR 조건은 94℃에서 2 분간 initial denaturation 과정 후 94℃에서 20초, 58℃에서 20초, 72℃에서 30초의 증폭반응을 18S rRNA 증폭 시 40회 반복하였고, MT isoform 증폭 시 45 회 반복하여 실시간 증폭 양상을 분석한 Ct 값 (threshold cycle)을 산출하였다 (Schmittgen and Livak, 2008).
실험자극 처리 및 발현 정량
중금속 노출에 의한 돌돔 MT isoform들의 발현 양상 분석을 위해서 실험 노출을 수행하였다. 중금속 노출 실험을 위해
12 L 용량의 4개 수조에 8 L (용존산소 = 6.0 ± 1.0; 수온:
22℃)의 여과해수와 돌돔 (어체중 = 2∼3 g)을 8마리씩 각각 수용하였다. 실험그룹에 사용한 중금속 종류는 카드뮴 (Cd), 구리 (Cu), 아연 (Zn)으로 25 ppb의 농도를 48 시간 동안 immersion 처리하였으며 대조군은 여과해수만을 사용하여 실 험군과 동일하게 48 시간 유지한 다음 무작위로 4마리씩 선별 하여 간 조직을 적출하였다.
세균 감염에 대한 간, 신장, 비장 조직에서의 MT-A 및 MT-B 발현 양상 분석을 하였다. 공격실험을 위해 각 50 L의 여과 해수를 담고 있는 60 L 용량의 4개 수조에 돌돔 개체 (평균 무게 = 121 ± 23 g) 6마리씩을 각각 수용하였다. 감염원으로서 Escherichia coli (Gram-; XL1 Blue MRF'), Edwardsiella tarda (Gram-; FSW910410) 및 Streptococcus iniae (Gram+; JSL0108) 를 이용하였다. E. coli와 S. iniae는 Luria-Bertani broth (LB) 배지를 그리고 E. tarda는 tryptic soy broth (TSB + 1% NaCl) 배지를 사용하여 28℃ 또는 37℃에 배양하였고 배양한 세균은 phosphate buffered saline (PBS, pH 7.2)으로 2회 washing한 다음 복강주사를 위해 1 × 105 cell/200 μl의 농도로 PBS에 현탁하였다. 복강 주사는 26 G 주사 바늘을 이용하여 실시하 였고, 이때 대조군은 세균 없이 PBS 200 μl만을 주사하였다.
주사 48 시간 후, 실험군과 대조군에서 각 4마리씩 무작위로 선발하여 간, 신장, 비장을 적출한 후 발현 분석에 이용하였다.
중금속 노출 실험 및 세균 감염 실험 후 확보한 조직들로부 터 total RNA 및 cDNA 합성을 수행하였다. Total RNA 추출 및 정량, cDNA 합성방법은 앞서 조직 발현 분석에서 언급한 방법과 동일하게 수행하였다. 또한 real-time RT-PCR 분석에 사용한 프라이머 및 PCR 조건 역시 조직 발현 분석 실험과 동일하게 수행하였으며 MT isoform들의 mRNA 발현 분석은 18S rRNA로 각 시료를 normalization한 후 대조군과 comparative Ct method를 이용한 fold change 값을 측정 하였다 (Schmittgen and Livak, 2008).
통계 처리
MT 유전자의 발현율 평가를 위하여 Duncan's multiple range test에 의해 ANOVA test를 수행하였다. 중금속 노출 실험 및 세균 공격 실험을 수행한 실험군과 대조군 간의 MT 유전자 발현율 차이를 검증하기 위한 통계처리는 SPSS 프로그램 (ver. 10.1.3)을 이용하였고, P = 0.05 수준에서 평가하였다.
결 과
돌돔 MT isoform들의 cDNA, 아미노산 서열의 특징 돌돔 MT-A의 cDNA 총 길이는 376 bp로서 34 bp의 5'-UTR, 180 bp의 ORF 영역 (종결코돈 taa 제외) 그리고 poly (A) tail을 포함한 162 bp의 3'-UTR을 포함하고 있었다 (GenBank accession no. JF419527). 반면 MT-B의 총 cDNA 길이는 373 bp로 5'-UTR는 40 bp, ORF 영역은 180 bp, 3'-UTR은 153 bp로 구성되어 있었다 (GenBank accession no. JF419528). MT-A와 MT-B는 모두 poly (A)+ tail로부터 17 bp 앞쪽에 전형적인 polyadenylation signal (aataaa)이 관찰되었다 (data not shown).
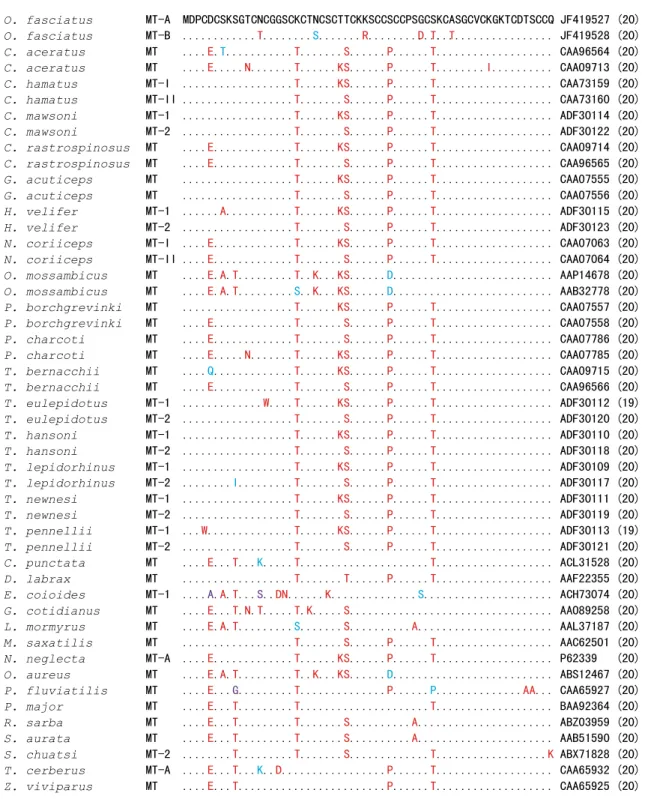
MT-A 및 MT-B는 공통적으로 60 개의 아미노산 중 20개의 cysteine 잔기 (33.3%)를 포함하고 있었고, 서열 내 cysteine들 의 존재 양상과 위치는 돌돔 MT isoform들 뿐만 아니라 분석한 농어목 대부분 어류들의 MT에서도 모두 공통적으로 잘 보전 되어 있었다. 돌돔의 MT-A와 MT-B간의 아미노산 서열은 6 영역 (아미노산 변환: Asn→Thr13, Asn→Thr22, Lys→Arg30, Gly
→Asp39, Ser→Thr41, Ala→Thr44)에서 차이가 관찰되었다 (Fig.
1). 농어목 어류의 MT 아미노산 서열 가운데 Cys 갯수가 다른 두 종 (Trematomus eulepidotus 및 T. pennellii)의 경우 공통적으 로 Cys14 또는 Cys4이 Trp으로 치환된 형태였다. 농어목 MT 어류의 가운데 돌돔 MT isoform들과 가장 높은 상동성을 나타 내는 ortholog는 Dicentrarchus labrax (AAF22355) MT였고 (MT-A 및 MT-B와 각각 95% 및 88% 상동성), 여타 농어목 어류 MT isoform들과 아미노산 수준에서 78∼93%의 상동성 을 나타내었다 (data not shown).
돌돔 MT-A 유전자 및 프로모터의 구조 분석
PCR 분리 및 genome walking을 통해서 확보된 돌돔 MT-A 유전자의 길이는 총 2198 bp로서 935 bp의 5'-upstream 영역 (ATG 개시코돈 기준), 654 bp의 구조유전자 영역 (개시코돈- 종결코돈), 그리고 609 bp의 3'-downstream flanking영역으로 구성되어 있었다. 돌돔 MT-A 구조 유전자는 3개의 exon으로 구성되어 있었으며 (tripartite 구조), 각 exon의 길이는 25, 66 및 92 bp였고 intron-I 및 intron-II의 길이는 118 및 348 bp로 확인되었으며 exon/intron junction 위치에서 전형적인 gt/ag 염기서열이 관찰되었다 (Fig. 2). MT-A의 5'-upstream 영역에는 개시코돈을 기준으로 80 bp 앞 쪽에 TATA box가 위치하고 있었고, metal regulatory transcription factor 1 (MTF-1)의 binding위치는 모두 5개로 확인되었으며 그 가운데 정방향의 인식 motif는 개시코돈 앞쪽으로 541 bp 및 109 bp에, 그리고 역방향의 인식 motif는 개시코돈 앞쪽으로 566 bp, 171 bp 및 133 bp에 위치하고 있었다. Aryl hydrocarbon receptor (AhR) 인식 위치는 모두 역방향의 인식 motif로서 개시코돈 앞쪽으 로 134 bp 및 127 bp에 2개가 확인되었다. 그리고 면역, 스트레 스 관련 전사조절인자 인식 motif들로서 CCAAT/enhancer- binding protein (C/EBP), activator protein 1 (AP-1), hepatic nuclear factor (HNF-3) 및 nuclear factor for activated T-cells (NF-AT) site들이 각 1개씩 예측되었다 (Fig. 2).
돌돔 MT-B 유전자 및 프로모터 구조 분석
MT-B 유전자 역시 MT-A 유전자와 동일하게 3개의 exon으 로 구성되어 있었으며 각 exon 길이는 MT-A와 동일하였고 exon-intron splicing rule 역시 gt/ag 규칙으로 잘 보전되어 있었 다. 그러나 돌돔 MT-B intron들의 길이는 각각 1199 bp 및 807 bp로서 MT-A에 비하여 각각 1081 bp 및 469 bp 긴 것으로 확인되었다 (Fig. 3). MT-B의 5'-upstream 영역의 TATA box의 위치는 MT-A와 마찬가지로 개시코돈 앞쪽으로 80 bp에 위치 하고 있었다. 그리고 정방향의 MTF-1 인식 위치는 개시코돈 에서 528 bp 및 109 bp 앞쪽에서 관찰되었고, 역방향의 MTF-1 인식 위치는 개시코돈에서 561 bp, 550 bp, 171 bp 및 133
bp 앞쪽에 위치하고 있어 총 6개가 존재하는 것으로 확인되었 다. AhR motif는 MT-A와 동일하게 역방향으로 2개가 존재하 였고, 위치 또한 동일하였다. MT-A와 유사하게 AP-1 motif는 개시코돈 앞쪽으로 609 bp위치에, HNF-3과 NF-AT motif들은
각각 835 bp 및 317 bp 앞쪽에서 예측되었다. 반면, MT-A의 5'-upstream 영역에서는 관찰되지 않은 upstream stimulatory factor (USF) motif가 개시코돈 앞쪽으로 1593 bp 및 1475 bp의 위치에 정방향으로 2개 존재하였다 (Fig. 3).
Fig. 1. Multiple sequence alignment of Oplegnathus fasciatus MT isoforms along with representative orthologs from perciform species. Dots indicate the identical amino acid residues with the O. fasciatus MT-A. The accession code for each sequence is provided at the end of the sequence, and the number of cysteine residues is indicated in the parenthesis following the each accession code.
Fig. 2. Gene and promoter structure of Oplegnathus fasciatus MT-A isoform. Coding regions are indicated by bold uppercase letters while non-coding regions by lowercase letters. In the 5'-flanking region, putative transcription factor binding sites are noticed by arrows wherein the orientation of each sequence is indicated by direction of the arrow. The site for the beginning of polyadenylation is noted by a broken arrow. Figures at the left are distance from the ATG translation initiation codon (+1).
Fig. 3. Gene and promoter structure of Oplegnathus fasciatus MT-B isoform. Coding regions are indicated by bold uppercase letters while non-coding regions by lowercase letters. In the 5'-flanking region, putative transcription factor binding motifs sites are noticed by arrows wherein the orientation of each sequence is indicated by direction of the arrow. The examined site for the beginning of polyadenylation is noted by a broken arrow. Figures at the left are distance from the ATG translation initiation codon (+1).
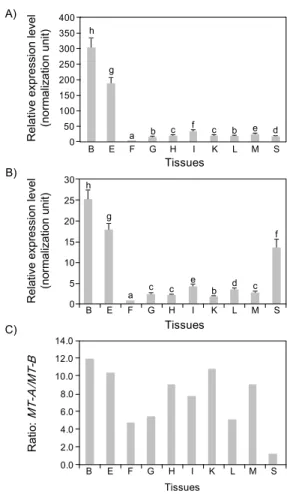
Fig. 4. Tissues distribution patterns and basal expression levels of Oplegnathus fasciatus MT-A (A) and MT-B (B) transcripts, as judged by real-time RT-PCR assay. Expression level of MT-A relative to MT-B in each tissue is also provided in (C). Mean±SDs with different letters are significantly different based an ANOVA followed by Duncan's multiple range test (P<0.05).
돌돔 MT-A 및 MT-B transcripts의 조직 분포 및 발현 수준
돌돔 조직들을 대상으로 MT-A 및 MT-B isoform들의 기초 발현 양상을 조사한 결과, 분석한 모든 조직에서 두 isoform의 transcripts가 검출되었으나 isoform 및 조직별 mRNA 발현 수 준은 다양하게 관찰되었다 (Fig. 4). MT-A transcripts의 경우 분석한 10 종류의 조직들 가운데 뇌 (brain)에서 가장 높은 발현율을 나타내었고 (P<0.05), 그 다음으로 눈 (eye)에서 높은 발현율을 나타내었다. 그리고 뇌와 눈 조직들을 제외한 나머지 조직들 경우 상대적으로 낮은 수준의 발현율을 나타내 었다. 뇌와 눈 조직을 제외한 나머지 조직들 간의 비교 시 소화관 (intestine), 근육 (skeletal muscle), 심장 (heart)/신장 (kideny), 간 (liver), 비장 (spleen), 아가미 (gill) 순으로 높은 발현율을 나타내었고, 지느러미 (fin)에서 가장 낮은 발현율을 나타내었다 (P<0.05). MT-B의 transcripts 역시 MT-A에서와 유사한 경향을 나타냄으로써, 뇌 및 눈 조직에서 상대적으로 높은 발현율을 나타내었고 (P<0.05), 여타 조직들에서는 상대 적으로 낮은 발현율을 나타내었다. 그러나 MT-A와 비교 시 MT-B의 경우 비장 (spleen)에서 유의적으로 높은 상대 발현이 관찰되었으며 역시 MT-A와 마찬가지로 지느러미에서 가장 낮은 발현을 나타내었다 (P<0.05).
각 조직별 두 MT isoform들의 발현 수준을 비교한 결과, 조사한 대부분 조직에서 MT-A는 MT-B보다 유의적으로 높은 mRNA 발현 수준을 나타내었으며 (P<0.05), 그 발현 수준의 차이는 조직별로 다양하게 관찰되었다. 뇌, 눈 및 신장의 경우 10배 이상의 발현 수준 차이가 관찰되었고, 소화관, 심장 및 근육의 경우 7∼9배, 그리고 지느러미, 아가미 및 간에서는 4∼6배의 발현 수준의 차이가 관찰되었다. 반면 비장의 경우 두 isoform들 간 기초 발현 수준이 유사한 것으로 관찰되었다 (Fig. 4).
Fig. 5. Transcriptional modulation of Oplegnathus fasciatus MT isoforms in the liver during acute exposure to 25 ppb of copper (Cu), cadmium (Cd) or zinc (Zn). For 48 h, as judged by real time RT-PCR assays. Mean±SDs with different letters are significantly different based an ANOVA followed by Duncan's multiple range test (P<0.05).
Fig. 6. Altered mRNA expression of Oplegnathus fasciatus MT-A and MT-B in the liver (A), kidney (B) and spleen (C), resulting from experimented bacterial challenges with Escherichia coli (EC), Edwardsiella tarda (ET) or Streptococcus iniae (SI). Mean±SDs with different letters are significantly different based an ANOVA followed by Duncan's multiple range test (P<0.05).
중금속 노출에 의한 간 조직 내 MT isoform들의 발현 변화
구리 및 카드뮴, 아연 노출 실험 결과, MT-A와 MT-B 공통적 으로 모두 카드뮴에서 가장 높은 유도 발현이 관찰되어 MT-A 의 경우 대조군 발현 수준에 비해 38배, 그리고 MT-B의 경우 77배의 증가된 transcripts 수준을 나타내었다 (P<0.05). 반면 아연 노출 그룹에서는 MT-A 및 MT-B 모두 약 4배 증가된 mRNA 수준이 관찰되었다. 그러나 구리 노출에서 MT-A의 경 우 유의적인 발현의 변화가 없었던 반면, MT-B의 경우 오히려 3배 이상 감소하는 경향을 나타내었다 (P<0.05) (Fig. 5).
세균 공격 실험에 의한 MT-A 및 MT-B의 전사 발현
특징
돌돔 MT isoform들은 모두 세균 공격 실험에 의해 유의적인 전사 발현 변화가 야기될 수 있는 것으로 나타났다 (Fig. 6).
간 조직 내에서의 MT-A 발현율은 E. coli (EC), E. tarda (ET) 및 S. iniae (SI) 처리에 대하여 모두 감소하는 것으로 확인된 반면, MT-B는 동일 세균 감염 처리에 대하여 2∼4배 증가된 mRNA 발현을 나타내었다 (Fig. 6A). 신장 조직 내에서는 MT-A 및 MT-B 모두 ET 처리에 의한 높은 유도 발현이 관찰되 었고 (P<0.05), MT-A는 SI 및 EC 순으로 발현이 높았던 반면, MT-B는 EC 및 SI 공격 순으로 발현이 높았다 (Fig. 6B). 비장 조직 내에서의 MT-A 및 MT-B 발현 역시 모두 ET 처리에 의한 유도 발현이 가장 높게 나타났다 (Fig. 6C).
고 찰
본 연구를 통해서 돌돔 MT isoform들의 유전자 및 프로모터 영역에 대한 구조적 특징 분석과 중금속 노출 및 박테리아 공격실험에 의한 발현 조절 양상을 조사하였다. 본 연구에서 발굴된 돌돔 MT isoform들의 아미노산 서열은 농어목 어류들 을 포함한 어류 MT 서열들과 매우 높은 상동성을 나타내었으 며, 특히, 대부분 종들에서 공통적으로 시스테인 (cysteine) 잔기들은 그 위치와 존재 형태가 특징적으로 잘 보전되어 있었다 (Coyle et al., 2002). 돌돔에서 분리된 2개의 isoform들 은 6개 영역에서 아미노산 차이가 관찰되어 여타 종내 MT paralog들 간의 차이 정도와 비교할 때 상대적으로 큰 차이를 나타내었지만, 두 isoform들 간의 분자량 (MT-A: 5.99 kDa 및 MT-B: 6.09 kDa) 및 등전점 (isoelectric point; pI; MT-A:
8.24 및 MT-B: 8.05)에서는 유의적인 차이가 없었다. Genomic DNA 수준에서도 역시 돌돔 MT isoform들은 모두 현재까지 알려진 척추동물 MT 유전자들과 마찬가지로 3개의 exon들로 구성된 tripartite 구조를 나타내어 이전의 연구 보고들과 유사 하였다 (Yan and Chan, 2004; Cho et al., 2005, 2008). 그러나 본 연구에서 분리된 돌돔 MT-B 유전자의 경우 여타 어류 MT 유전자들과는 달리 매우 긴 intron 길이를 갖고 있는 것으로 나타나 흥미로웠다. 돌돔 MT-B의 경우 intron-I 및 intron-II에 서 각각 1199 bp (돌돔 MT-A: 118 bp) 및 807 bp (돌돔 MT-A:
353 bp)의 길이를 나타냄으로써, intron-I의 경우 MT-A에 비해 10배 이상으로 길었다. 또한 많은 선행 연구들에서 어류 MT
유전자들은 adenine 및 thymine 빈도 (A+T%)가 높은 intron을 갖는다고 알려져 있으며 (Cho et al., 2008, 2009a), 본 연구에서 발굴된 돌돔 MT-A의 경우 앞서 연구 결과들과 유사한
“A+T-rich” intron 들을 나타내었지만 (intron-I: 63% 및 intron-II: 60%), 돌돔 MT-B 유전자의 경우 상대적으로 낮은 A+T% (intron-I 및 intron-II에서 각각 59% 및 57%)를 보였다.
아직 돌돔 MT-B 유전자가 전형적인 어류 MT 유전자들의 길이 와 확연히 다른 유전자 크기를 갖는 정확한 이유는 알 수 없으며 추가 연구가 필요한 상태이다. 일반적으로 척추동물 유전자의 크기는, 특히 non-coding region의 경우, 해당 종의 genome size와 비례한다고 알려져 있으나, 돌돔의 경우 전형적 인 농어목 돔류 어종들의 genome size를 갖기 때문에 돌돔의 genome size (C-value: 0.93; http://www.genomesize.com/)에 비 례하여 MT 유전자 길이가 증가되었다고 판단하기는 어렵다 (see also Cho et al., 2009c). 어류의 경우 분류군에 따라 진화 과정 중 유전체 복제 (genome duplication)를 통해서 다양한 isoform 유전자들을 획득한 바 있음이 알려져 있으며, 특히 면역 및 스트레스에 관련한 유전자들의 경우 빈번한 유전자 복제 (gene duplication)가 일어난다고 보고된 바 있다 (Cho et al., 2009a, 2009b). 아직 본 연구의 MT-B의 자세한 진화 경로는 알 수 없으나 MT-A가 전형적인 척추동물 MT 유전자의 구조를 나타내는 반면 MT-B는 tripartite 구조를 가지지만 아직 경골어류에서는 관찰되지 않는 유의적으로 긴 intron길이를 보유한다는 점에서 본 MT-B는 돌돔 내에서 최근의 분화 또는 복제에 의한 산물일 가능성이 높으며, 따라서 MT-B와 유사한 isoform ortholog들이 근연종들에서도 관찰되는지에 대한 추 가 탐색이 필요하다고 판단된다. 또한 향후 본 연구에서 관찰 된 두 종류의 MT isoform들에 대한 genomic 유전 좌위 (chromosomal locus) 및 분자계통 분석이 이루어진다면 상기 가설에 대한 보다 선명한 근거를 제시할 수 있으리라 기대된 다. 어류 MT isoform 유전자들의 경우 분류 군에 따라 서로 다른 진화 또는 복제 경로를 거친다고 알려져 있으며, 미꾸라 지 (Misgurnus mizolepis; Cypriniformes)에서 종 특이적인 MT isoform들의 분화와 isoform들이 tandem genomic organization 형태로 존재한다고 보고된 바 있다 (Cho et al., 2009a). 경골어 류들은 일반적으로 2개의 MT isoform을 갖는다고 알려져 있 으나 아직 발굴된 어류 MT isoform들의 정확한 분류 기준 및 명명체계는 정립되어 있지 못한 상태이며, 뿐만 아니라 일부 어종에서는 3개 이상의 MT isoform들이 존재함이 보고 되어 있다 (Knapen et al., 2005). 따라서 본 연구에서 발굴된 2종류의 MT isoform외에 돌돔 genome내 추가적인 MT isoform 들의 존재 가능성에 대한 연구와 함께 본 연구에서 명명한 MT-A 및 MT-B 명칭 역시 추후 어류 MT isoform들의 진화 및 분화 경로에 관한 추가 분석을 바탕으로 신중한 재검토가 역시 필요할 수 있다고 예상된다.
Non-coding 영역에서의 큰 차이점에도 불구하고 돌돔 MT-A와 MT-B 유전자들은 5'-upstream 조절 영역에서 유사한 전사조절인자 결합 위치 (transcription factor binding sites;
TFBS) 양상을 나타내었음은 물론, 본 연구에서 예측된 대부분
의 TFBS들은 이미 여타 척추동물 MT 프로모터들에서 관찰된 것으로서, 따라서 돌돔 MT isoform들 역시 척추동물 MT 유전 자의 일반적인 전사 조절 기작을 잘 보전하고 있으리라 예상 되었다. 특히 MT 등 중금속 관련 유전자들의 발현 조절에 중추적인 역할을 담당한다고 알려져 있는 metal transcription factor 1 (MTF-1)이 부착하는 위치 (metal responsive element;
MRE; consensus sequence = TGCRCNC)가 MT-A 및 MT-B에서 각각 5개 및 6개로 관찰되었고, 이 중 3개의 MRE copies (-171 bp, -133 bp 및 -109 bp)는 MT-A와 MT-B 모두 동일한 위치에서 예측되었다. 따라서 본 연구에서 관찰된 돌돔 MRE copies의 분포 양상은 경골 어류 MT 프로모터에 존재하는 MRE motif들 은 일반적으로 여러 copies가 군집 (cluster) 형태로 존재하며, proximal 프로모터에 존재하는 MRE cluster는 1 kb 이내에 존재한다는 특징과 잘 일치하였다 (Olsson et al., 1995; Samson et al., 2001; Yan and Chan, 2004; He et al., 2007; Cho et al., 2009a). 한편 돌돔 MT 프로모터에서 예측된 xenobiotic responsive element (XRE; consensus sequence = CACGCW)는 aryl-hydrocarbon reductase (AhR)가 부착하는 위치로서, AhR 은 다양한 독성 오염원의 해독에 관여하는 유전자의 전사 조절에 관여하는 것으로 알려져 있고 (Tanguay et al., 1999;
Tijet et al., 2006), 또한 일부 어류 MT 프로모터에서도 동정된 바 있으며, 어류 MT가 중금속 외에 여타 독성 오염원 반응 경로에 관여함을 잘 보여주고 있다 (Cho et al., 2008, 2009a).
그 외 돌돔 MT 프로모터에서 예측된 TFBS들로서 AP-1, USF 및 C/EBP site들은 염증 및 면역 반응에 관여하는 유전자들에 서 쉽게 관찰되는 TFBS들로서 항균 펩타이드 (antimicrobial peptide) 유전자 (Cho et al., 2009b), 항산화 효소 (antioxidant enzyme) 유전자 (Cho et al., 2009c) 및 어류 MT 유전자들 (He et al., 2007)에서 보고된 바 있다. 따라서 MT 단백질이 어류에 서도 역시 선천성 면역 반응 및 항산화 방어 등에 관여할 수 있음을 간접적으로 시사한다.
돌돔 MT-A와 MT-B 모두 분석한 10 종류의 조직에서 항시 발현하는 것으로 관찰되었으며, 이러한 광범위한 MT transcripts의 조직 분포 양상은 현재까지 보고된 대부분의 MT 연구 결과들과 잘 일치한다. 하지만 본 연구뿐만 아니라 종래 의 모든 선행 연구들에서 MT transcripts의 발현은 비록 거의 모든 조직에서 검출되지만 그 발현 수준은 조직 별로 다르며, 조직 별 발현 양상은 어종 마다 서로 차이가 있다고 보고되어 있다 (Hermesz et al., 2001; Kim et al., 2008; Cho et al., 2009a).
본 연구에서 두 종류 돌돔 MT isoform들은 모두 뇌 (brain) 및 눈 (eye)에서 가장 높은 수준의 mRNA 발현을 보였는데 이러한 양상은 암수동체 점박이송사리 (Kryptolebias marmoratus; Cyprinodontiformes)와 유사하였다 (Rhee et al., 2009). 또한 돌돔 MT isoform들의 경우 MT-A가 MT-B에 비하 여 대부분 조직에서 훨씬 높은 기초 발현 수준을 나타내었는 데 이는 미꾸라지에서 관찰된 바 있는 MT-IA 및 MT-IB의 결과와 유사하였다 (Cho et al., 2009a). 이처럼 두 MT isoform들 간에 기초 발현에 큰 차이가 있다는 점은 두 isoform 사이에 항시구성적 (ubiquitously constitutive) 항상성 조절
(homeostatic regulation)과 해독 작용 (detoxification function) 간에 관한 잠재적 기능 분화 등이 이루어졌을 가능성을 시사 하지만, 아직 이를 입증하기 위해서는 추후 더 많은 실험 자극 과 발현 분석 실험이 뒤따라야 할 것이다 (Lee et al., 2010).
구리, 카드뮴 및 아연을 이용한 중금속 노출 실험 결과를 통해서 돌돔의 두 isoform들 모두 카드뮴 처리에 의해서 가장 높은 유도 발현이 관찰되었고, 아연에 의해서 비교적 낮은 유도 발현을 나타내었으며, 반면 구리 처리 군에서는 두 isoform들의 조절 방향이 서로 다르게 나타났다. 본 연구 결과 는 카드뮴이 MT 발현을 신속하게 유도한다는 종래 많은 연구 보고들과도 잘 일치하고 있다 (Bourdineaud et al., 2006; Rhee et al., 2009). 한편 동일 카드뮴 처리 조건에서 MT-B의 유도 발현 양이 MT-A에 비해 2배 이상 높게 나타났는데 이는 이들 프로모터에 존재하는 MRE copies의 개수 차이에 기인하는 것으로 판단되나 (Chan and Chan, 2008), 향후 본 연구에서 생물정보학 분석으로 동정된 각 MRE copies가 모두 실제 중금 속 반응 기능을 갖는지 또는 MRE에 따라 차등 반응 특징을 갖는지에 관한 후속 연구가 뒷받침 되어야 할 것이다. 한편 앞서 언급한 바와 같이 돌돔 MT-A와 MT-B isoform들은 구리 노출에 의해서 큰 차이를 나타내었는데, 본 연구 결과는 중금 속 별로 유도 발현뿐만 아니라 MT의 억제가 함께 일어날 수 있음을 보고한 종래의 연구결과들과 유사하였다 (Hermesz et al., 2001; Woo et al., 2006; Cho et al., 2009a). 또한 동일 중금속 처리라 할지라도 어종 별로 확연히 다른 MT 발현 양상을 나타내며, 중금속에 대한 내성 역시 어종 별로 차이가 있음이 보고되어 있으므로 (Egg et al., 2000), 본 연구 결과를 바탕으로 돌돔 MT를 중금속 표지 바이오마커로 이용하기 위해서는 다양한 중금속에 대하여 다양한 농도와 처리 조건을 사전에 검증해야 할 것이다 (Serafim and Bebianno, 2001;
Vasconcelos et al., 2002)
중금속 처리뿐만 아니라 돌돔 MT isoform들은 비병원성 및 병원성 세균을 이용한 공격 실험에서도 유의적인 발현율의 변화를 나타내었다. 특히 두 isoform들 모두 신장과 비장에서 는 사용한 세균의 종류에 따라 다양한 발현 증가 폭을 나타내 었고, 특히 E. tarda 처리에 의해서 가장 많은 발현 증가가 관찰되었다. 이러한 면역 자극에 의한 MT 발현의 증가는 MT 가 선천성 면역에 관한 다양한 세포 활동에 밀접히 관계함을 시사한다 (Thirumoorthy et al., 2007). 아직 본 연구에서와 같이 어류 MT를 대상으로 면역 자극 실험을 수행한 선행 연구 결과는 많지 않지만 대서양 대구 (Gadus morhua)의 경우 Vibrio anguillarum과 Aeromonas salmonicida감염으로 인한 아 가미 조직에서 감염 3 시간 후 급속한 MT 발현의 증가가 확인되었고, 무지개송어의 경우 Myxobolus cerebralis 감염 처 리 2 시간 후 피부조직에서 MT-B 발현율이 5배 증가함이 보고된 바 있어 본 연구결과와 유사하였다 (Baerwald et al., 2008; Caipang et al., 2011). 한편 비장 및 신장 조직과는 달리 간 조직의 경우 박테리아 감염에 의한 MT 조절의 방향이 반대로 나타나 흥미로웠는데, 세균의 종류와 관계없이 MT-A 의 경우 간 조직에서 모두 발현이 억제된 반면, 반대로 MT-B는
모두 유의적인 증가 양상을 나타내었다. 이는 MT의 면역 반응 이 조직 별로 차이가 있음을 시사하며, 간 조직에서 두 isoform 들 간의 면역 반응 담당 역할에 차이가 있을 수 있음을 나타낸 다. 따라서 MT의 면역 관계 기능을 세분화하여 추적하기 위해 서는 조직 별 각 MT isoform들의 발현과 함께 염증 반응 지표 단백질들의 발현 (예컨대, 항균 단백질 및 cytokines류 등)을 동시에 추적할 필요성이 있다고 판단된다.
본 연구를 통해 돌돔으로부터 서로 다른 두 MT isoform 유전자들을 발굴하였고, 각 isoform 별 유전자 및 프로모터의 구조 해석과 함께 중금속 및 세균 감염에 대한 발현 반응을 평가하였다. 본 연구 결과, 두 isoform들은 척추동물 MT와 기본적으로 공통적인 유전자 구조를 갖지만 isoform별로 조직 발현 및 자극에 의한 유도 발현에 있어 공통적인 발현 반응 양상과 함께 isoform별 차등 발현 반응의 특징을 함께 나타내 었다. 따라서 본 연구를 바탕으로 각 isoform의 발현 조절에 영향을 미치는 다양한 생물학적 또는 무생물학적 요인들이 조사된다면, 유용 양식 품종인 돌돔의 스트레스 및 생체 방어 연구에 기여할 수 있다고 기대된다.
사 사
본 연구는 한국연구재단 한-우크라이나 국제협력기반 조성 사업 (국제공동연구 #2010-00091) 지원에 의해 수행되었음.
참고문헌
Baerwald MR, Welsh AB, Hedrick RP and May B. 2008.
Discovery of genes implicated in whirling disease infection and resistance in rainbow trout using genome-wide expression profiling. BMC Genom 9, 37.
Bourdineaud J-P, Baudrimont M, Gonzalez P and Moreau J-L. 2006. Challenging the model for induction of metallothionein gene expression. Biochimie 88, 1787-1792.
Caipang CMA, Lazado CC, Brinchmann MF, Rombout JHWM and Kiron V. 2011. Differential expression of immune and stress genes in the skin of Atlantic cod (Gadus morhua). Comp Biochem Physiol Genom Proteonomics, in press
Chan WWL and Chan KM. 2008. Cloning and characterization of a tilapia (Oreochromis aureus) metallothionein gene promoter in Hepa-T1 cells following the administration of various heavy metal ions. Aquat Toxicol 86, 59-75.
Chen FY, Liu HP, Bo J, Ren HL and Wang KJ. 2010.
Identification of genes differentially expressed in hemocytes of Scylla paramamosain in response to lipopolysaccharide. Fish Shellfish Immunol 28, 167-177.
Cho YS, Choi BN, Ha EM, Kim KH, Kim SK, Kim DS and Nam YK. 2005. Shark (Scyliorhinus torazame) metallothionein: cDNA cloning, genomic sequence, and expression analysis. Mar Biotechnol 7, 350-362.
Cho YS, Lee SY, Kim K-Y, Bang IC, Kim DS and Nam YK. 2008. Gene structure and expression of metallothionein during metal exposures in Hemibarbus mylodon. Ecotoxicol Environ Saf 71, 125-137.
Cho YS, Lee SY, Kim K-Y and Nam YK. 2009a. Two metallothionein genes from mud loach Misgurnus mizolepis (Teleostei; Cypriniformes): gene structure, genomic organization, and mRNA expression analysis. Comp Biochem Physiol B Biochem Mol Biol 153, 317-326.
Cho YS, Lee SY, Kim KH, Kim SK, Kim DS and Nam YK. 2009b. Gene structure and differential modulation of multiple rockbream (Oplegnathus fasciatus) hepcidin isoforms resulting from different biological stimulations. Dev Comp Immunol 33, 46-58.
Cho YS, Lee SY, Bang IC, Kim DS and Nam YK. 2009c.
Genomic organization and mRNA expression of manganese superoxide dismutase (Mn-SOD) from Hemibarbus mylodon (Teleostei, Cypriniformes).
Fish Shellfish Immunol 27, 571-576.
Choi BN, 2005. Molecular characterization and expression of antioxidant genes in rockbream (Oplegnathus fasciatus). MS. thesis, Pukyong National University, Busan, Korea.
Coyle P, Philcox JC, Carey LC and Rofe AM. 2002.
Metallothionein: The multipurpose protein. Cell Mol Life Sci 59, 627-647.
Egg M, Sturmbauer C and Dallinger R. 2000.
Metallothionein-A cDNA in Alpine Arctic char (Salvelinus alpinus): notes on metal tolerance and relationships to other salmonids. Neth J Zool 50, 1-14.
Gao D, Wang GT, Chen XT and Nie P. 2009.
Metallothionein-2 gene from the mandarin fish Siniperca chuatsi: cDNA cloning, tissue expression, and immunohistochemical localization. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol 149, 18-25.
Gasteiger E, Hoogland C, Gattiker A, Duvaud S and Wilkins MR. 2005. Protein identification and analysis tools on the ExPASy server. In: John M. Walker (ed): the proteomics protocols handbook. humana press, Totowa, N.J.U.S.A., 571-607.
Haq F, Mahoney M and Koropatnick J. 2003. Signaling events for metallothionein induction. Mutat Res 533, 211-226.
He P, Xu M and Ren H. 2007. Cloning and functional characterization of 5'-upstream region of metallothionein-I gene from crucian carp (Carassius cuvieri). Int J Biochem Cell Biol 39, 832-841.
Hermesz E, Abrahám M and Nemcsók J. 2001.
Tissue-specific expression of two metallothionein genes in common carp during cadmium exposure and temperature shock. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol 128, 457-465.
Jenny MJ, Warr GW, Ringwood AH, Baltzegar DA and Chapman RW. 2006. Regulation of metallothionein genes in the American oyster (Crassostrea virginica):
ontogeny and differential expression in response to different stressors. Gene 379, 156-165.
Kim JH, Wang SY, Kim IC, Ki JS, Raisuddin S, Lee JS and Han KN. 2008. Cloning of a river pufferfish (Takifugu obscurus) metallothionein cDNA and study of its induction profile in cadmium-exposed fish.
Chemosphere 71, 1251-1259.
Knapen D, Redeker ES, Inacio I, Coen WD, Verhyen E, Blust R. 2005. New metallothionein mRNAs in Gobio gobio reaveal at least three gene duplication events in cyprinid metallothionein evolution. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol 140, 347-355.
Knapen D, Reynders H, Bervoets L, Verheyen E and Blust R. 2007. Metallothionein gene and protein expression as a biomarker for metal pollution in natural gudgeon populations. Aquat Toxicol 82, 163-172.
Lee SY, Stoliar O and Nam YK. 2010. Transcriptional alteration of two metallothionein isoforms in mud loach (Misgurnus mizolepis) fry during acute heavy metal exposure. Fish Aqua Sci 13, 112-117.
Ohta H, Seki Y and Imamiya S. 1993. Possible role of metallothionein on the gastrointestinal absorption and distribution of cadmium. Kitasato Arch Exp Med 65, 137-145.
Olsson PE, Kling P, Erkell LJ and Kille P. 1995. Structural and functional analysis of the rainbow trout (Oncorhyncus mykiss) metallothionein-A gene. Eur J Biochem 230, 344-349.
Palmiter RD, Findley SD, Whitmore TE and Durnam DM.
1992. MT-III, a brain-specific member of the metallothionein gene family. Proc Natl Acad Sci USA 89, 6333-6337.
Rhee JS, Raisuddin S, Hwang DS, Lee KW, Kim IC and
Lee JS. 2009. Differential expression of metallothionein (MT) gene by trace metals and endocrine-disrupting chemicals in the hermaphroditic mangrove killifish, Kryptolebias marmoratus.
Ecotoxicol Environ Saf 72, 206-212.
Samson SL-A, Paramchuk WJ and Gedamu L. 2001. The rainbow trout metallothionein-B gene promoter:
contributions of distal promoter elements to metal and oxidant regulation. Biochim Biophys Acta 1517, 202-211.
Schmittgen TD and Livak KJ. 2008. Analyzing real-time PCR data by the comparative CT method. Nat Protoc 3, 1101-1108.
Serafim MA and Bebianno MJ. 2001. Variation of metallothionein and metal concentrations in the digestive gland of the clam Ruditapes decussatus:
sex and seasonal effects. Environ Toxicol Chem 20, 544-552.
Tanguay RT, Abnet CC, Heideman W and Peterson RE.
1999. Cloning and characterization of the zebrafish (Danio rerio) aryl hydrocarbon receptor. Biochim Biophys Acta Gene Struct Expr 1444, 35-48.
Thirumoorthy N, Manisenthil Kumar KT, Shyam Sundar A, Panayappan L and Chatterjee M. 2007.
Metallothionein: an overview. World J Gastroenterol 13, 993-996.
Tijet N, Boutros PC, Moffat ID, Okey AB, Tuomisto J and Pohjanvirta R. 2006. Aryl Hydrocarbon Receptor Regulates Distinct Dioxin-Dependent and Dioxin-Independent Gene Batteries. Mol Pharmacol 69, 140-153.
Tom M, Chen N, Segev M, Herut B and Rinkevich B.
2004. Quantifying fish metallothionein transcript by real time PCR for its utilization as an environmental biomarker. Mar Pollut Bull 48, 705-710.
Wang L, Song L, Ni D, Zhang H and Liu W. 2009.
Alteration of metallothionein mRNA in bay scallop Argopecten irradians under cadmium exposure and bacteria challenge. Comp Biochem Physiol C Toxicol Pharmacol 149, 50-57.
Wassermann K, Pirsel M and Bohr VA. 1992.
Overexpression of metallothionein in Chinese hamster ovary cells and its effect on nitrogen mustard-induced cytotoxicity: role of gene-specific damage and repair. Cancer Res 52, 6853-6859.
Woo S, Yun S, Jung JH, Shim WJ, Lee CH and Lee TK. 2006. Heavy metal-induced differential gene expression of metallothionein in Javanese Medaka, Oryzias javanicus. Mar Biotechnol 8, 654-662.
Vasconcelos MH, Tam SC, Hesketh JE, Reid M and Beattie JH. 2002. Metal- and tissue-dependent relationship between metallothionein mRNA and protein. Toxicol Appl Pharmacol 182, 91-97.
Yan CHM and Chan KM. 2004. Cloning of zebrafish metallothionein gene and characterization of its gene promoter region in HepG2 cell line. Biochim Biophys Acta 1679, 47-58.
2011년 2월 22일 접수 2011년 4월 8일 수정 2011년 4월 8일 수리