Classification and Expression Analysis of Cytochrome P450 Genes from Soybean
Bong-Gyu Kim, Jae Hyeong Ko, Hor-Gil Hur1 and Joong-Hoon Ahn*
Bio/Molecular Informatics Center, Konkuk Univeristy, Seoul 143-701, Korea
1Department of Environmental Science and Engineering, Gwangju Institute of Science and Technology, Gwangju 500-712, Korea
Received October 26, 2004; Accepted November 18, 2004
Cytochrome P450 monooxygenases in plants are involved in various metabolic pathways. Synthetic pathways for phenylpropanoid, fatty acid, terpenoid and various plant hormones are only parts in which P450s are involved. With vast information available from various plant genome projects, many P450 sequences from several plants can be accessed. However, only few of their functions are known.
Based on P450 classification system, eight-two soybean P450 genes were classified and their expression patterns in different tissues and upon UV-B irradiation analyzed by RT-PCR. In most soybean P450 genes, its expression pattern is correlated with P450 clan to which individual P450 belongs. It would be a first step to understand biological functions of individual P450s from soybean.
Key words: Cytochrome P450, Glycine max, Flavonoids, Phenylpropanoids, Second Metabolism
All organisms perform oxidative metabolism toward a vast number of endogenous and exogenous chemicals. Among the several enzymes involved in these reactions, cytochrome P450 monooxygenase activates molecular oxygen through the insertion of one of its atoms into the substrate and reduction of the other to form water. Most of P450s use NADPH as an electron donor but interact with a flavonoprotein known as a P450 reductase that transfers electrons.1) P450s are present in microorganisms, plants, and animals2) with extensive attention paid to those that function in the detoxification of xenobiotics from animals and bacteria.
Plants have a large multigene superfamily of P450 because plant P450s participate in many biochemical processes such as biosynthesis of phenylpropanoid, alkaoid, terpenoid, fatty acid, plant growth regulator, and cyanogenic glycosides.1) In facts, Arabidopsis thaliana, whose genome sequencing was completed, is estimated to have 273 P450s,3,4) and rice also has more than 300 different P450s.5) It is represented about 1% of total genes in Arabidopsis and rice. Other organisms such as human, mouse, Drosophila melanogaster, and Caenorhabditis elegans have between 54 and 105 P450s, representing approximately 0.1 to 0.5% of total genes.6) The number of P450 in plants is much higher than in other organisms, because plants produce more chemicals and detoxify xenobiotics. The classification of plant P450 is based on its amino acid homology and plant P450s are assigned names from CYP71A1 to CYP99XY, then from CYP701A1 and above. If the amino acid sequence of a new P450 shows more
than 40% homology, it is said to be a member of the same clan. Name of cytochrome P450 denotes the following order;
family (clan), subfamily, and isoform. For example, CYP71A1 means it belongs to clan 71, subfamily A and isoform 1. Unlike bacterial P450s, most plant P450s are bound to membranes through a short hydrophobic segment of their N-terminus;7) thus, analysis of plant P450 sequence reveals potential signal peptide.
Ultraviolet radiation causes damages to DNA and other macromolecules, which lead to the cellular metabolism changes and eventually death. Plants develop means to cope with the exposure to UV. Phenolic compounds, which are synthesized via the phenylpropanoid pathway, draw attention in the protection of plants against UV radiation. Among many phenolic compounds, flavonoids including antocyanin have been demonstrated to be involved in UV-protection.8,9) The Arabidopsis mutant tolerant to the UV-B showed elevated levels of flavonoids and other phenolics.10) Many P450s are involved in the phenylprophanoid pathway that leads to the biosynthesis of lignin, flavonoids, and antocyanin. The expression of P450s involved in flavonoid biosynthesis such as flavanone 3- hydroxylase (F3H) and leucoanthocyanidin dioxygenase (LDOX) was enhanced by UV in carrot.11) However, in spite of the vast amount of sequences from several plant genomes available, the biological functions of most genes are unknown.
Thus, development of effective analysis methods is necessary to elucidate the biological functions.
In this study to help understand the biological functions of soybean P450s, soybean P450s were classified based on the P450 classification system, and the expression pattern of soybean p450s in the response to UV-B irradiation and different tissues were analyzed.
*Corresponding author
Phone: +82-2-450-3764, Fax: +82-2-456-7183 E-mail: [email protected]
Materials and Methods
Plant materials and UV-irradiation. Soybean (Glycine max) cultivar Sinpaldal was grown for 3 weeks. After 21 days, soybean plants were transferred to the growth chamber containing three UV-B lamps (Philips TL 20W/12 UV) and subjected to UV-B treatment. The plants were located to 20-25 cm below the UV-B lamps.
Classification of P450. Partial or full soybean P450 sequences in The Institute For Genomic Research (TIGR, www.tigr.org) were used as query sequences for the BLAST search in NCBI (http://www.ncbi.nlm.nih.gov/BLAST/) and Arabidopsis cytochrome P450 database (http://132.192.64.52/
p450.html.).
Reverse transcription PCR. Total RNA was isolated from the sample harvested at 1, 2, 4, 8, 12, 16, 20, and 24 h after UV-B irradiation. cDNA was synthesized in 20-µl reaction mixtures containing 2µg of total RNA, Ominiscript transcriptase (Qiagen, Gaitherburg, MD, USA), 15 pmol of oligo (dT)15, and 20 U of Rnasin (Promega, Madison, WI, USA). Polymerase chain reaction was performed with Hotstart DNA polymerase (Qiagen, Gaitherburg, MD) under the following conditions: 28 cycles of 1 min at 94oC, 1 min at 55oC, and 1 min at 72oC. Primers were designed based on the published P450 sequences in TIGR.
Results and Discussion
Genome sequencing project of several plants are in progress. Because soybean is one of the crop plant with a great economical impact, EST(Expressed Sequence Tag) project is in progress.
Plant P450s were classified based on their amino acid homology. Soybean P450 sequences from TIGR were used to classified based on a plant P450 classification system. The individual P450 sequence from TIGR was used as a query for BLAST. The database searched was either NCBI BLAST search or Arabidopsis P450 BLAST search. Even though some soybean P450s examined in this study have partial sequences, we were enable to find the best matched P450 homologues based on the P450 classification system.
However, an accurate and complete sequencing could change the clan of the present classification into a new one. Eighty- two soybean P450 genes used belonged to a particular subgroup of P450 (Table 1). Among 30 plant P450 clans, P450 genes used cover 22 clans. Because soybean is expected to have more P450 genes than Arabidopsis and rice, soybean that produces more secondary metabolites could have at least one gene in each clan in contrast to Arabidopsis and rice, both of which lack 12 clans.5)
P450s are involved in myriad biochemical pathways.4,6) The expression pattern of each P450s upon stimulation or in different tissue is different. For example, members of CYP82 are highly stress-induced12) and some of CYP93 are induced by methyl jasmonate in soybeans.13) Stimuli that regulate the
expression of P450 biosynthesis are various including UV-B, which could be one of stimuli that could either enhance or decrease the expression of P450. Furthermore, UV-B radiation has many direct and indirect effects on plants including damages to DNA, proteins, and membranes, alterations in transpiration and photosynthesis, and changes in growth, development and morphology.14) In addition, UV-B induces accumulation of a range of secondary metabolites including flavonoids, anthocyanins, alkaloids, waxes, and polyamines8) and up-regulates genes of the phenylpropanoids pathway.15,16) Many of the responses that are regulated by UV- B are mediated by P450s. Thus, we irradiated soybeans with UV-B and investigated the changes in the expression pattern of soybean P450 genes. To determine the UV-B irradiation time, we first examined the expression of chalcone synthase (chs) upon UV-B irradiation. CHS is located at the entry point of the flavonoid biosynthetic pathway and thus would act as an indicator gene because UV-B irradiation causes the accumulation of flavonoids. In fact, expression of chs in A.
thaliana was enhanced up to 6h and its expression in parsley cell was increased by UV.8, 15) The expression of chs in soybean was examined. The soybean plant was harvested at 0, 4, 8, 12, 16, 20 and 24 h after UV-B irradiation. The expression of chs at 0 h was very low, then increased reaching maximum at 12 h and decreased (Fig. 1). Therefore, samples harvested at 8 and 12 h after UV-B irradiation were used for further experiments.
Eight-two soybean P450 primer sets were designed based on the published soybean P450 sequences to study the expression profile of each member of the soybean P450s in response to UV-B irradiation, and RT-PCR was performed.
The specificity of each primer was checked by sequencing the PCR product. Twenty-nine P450 genes exhibited induction compared to the UV-untreated sample, and 13 decreased (Fig 2). Some, such as SP51 and SP61 showing induction were only expressed with UV-B irradiation while others were not expressed upon induction. Expression pattern of individual P450 was not always related to its clan. For example, even though SP30 and SP61 belonged in same family and subfamily (CYP78A), SP61 increased while SP30 decreased.
CYP98 family is known to contain P450s involved in phenylpropanoid pathway whose products are accumulated upon UV-B irradiation.17-19) SP58 that is classified as CYP98 increases its expression upon UV-B irradiation, suggesting the classification based on the amino acid sequence homology would be helpful to understand the actual function of a gene.
Flavonoid is one of the indicators of UV-B irradiation. Thus, CYP75 containing genes for the flavonoid hydroxylation20) is expected to increase upon UV-irradiation. SP10 and SP71 belong to CYP75. Expression of SP71 was enhanced, while that of SP10 did not change. CYP 86 is involved in fatty acid metabolism21,22) and SP54 was classified as CYP86 and up- regulated. These results suggest that P450 classification and examination of gene expression pattern upon various stimuli could give a clue about the biological function of individual
genes.
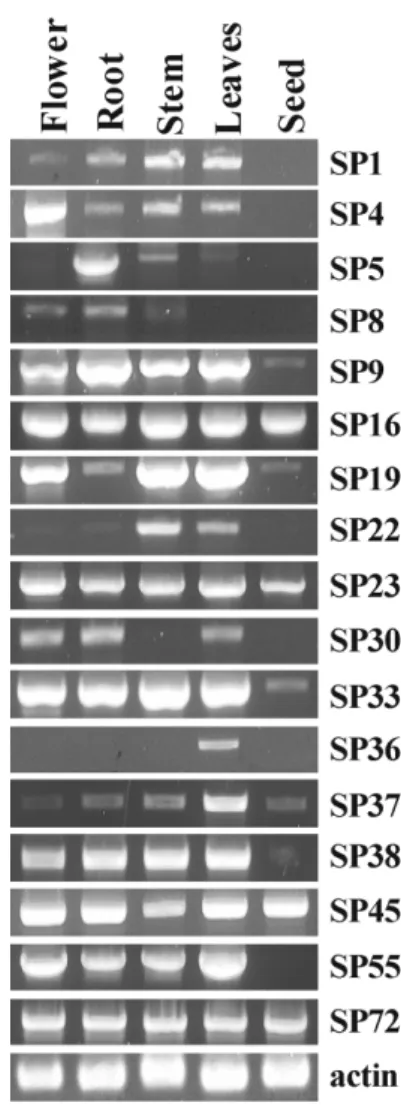
We examine the expression of 17 soybean P450s in different tissues (Fig. 3). Total RNA was isolated from flowers, roots, stems, leaves and seeds of soybean plant and RT-PCR was performed. Overall expression pattern was quite unique.
Relative expression level showed difference with the different tissues. For example, overall expression levels of SP5, SP8, SP22 and SP36 were relatively low. SP5 was expressed mainly in the root whereas SP36 was expressed only in the leaves. SP16, SP45, and SP72 were expressed in all the tissues
investigated, even though their expression patterns in response to UV were different. In the case of isoflavone synthase known to catalyze naringenin into genistein, it is expressed mainly in seeds, roots, and nodules because the content of isoflavone in soybean seed is approximately 1.8 mg/g seed23) and also isoflavone has a role in attracting nitrogen-fixation bacteria from rhizosphere. In other hand, chalcone synthetase (chs) is expressed mainly in leaves and stems since it serves as a precursor for the biosynthesis of other flavonoids(data not shown).
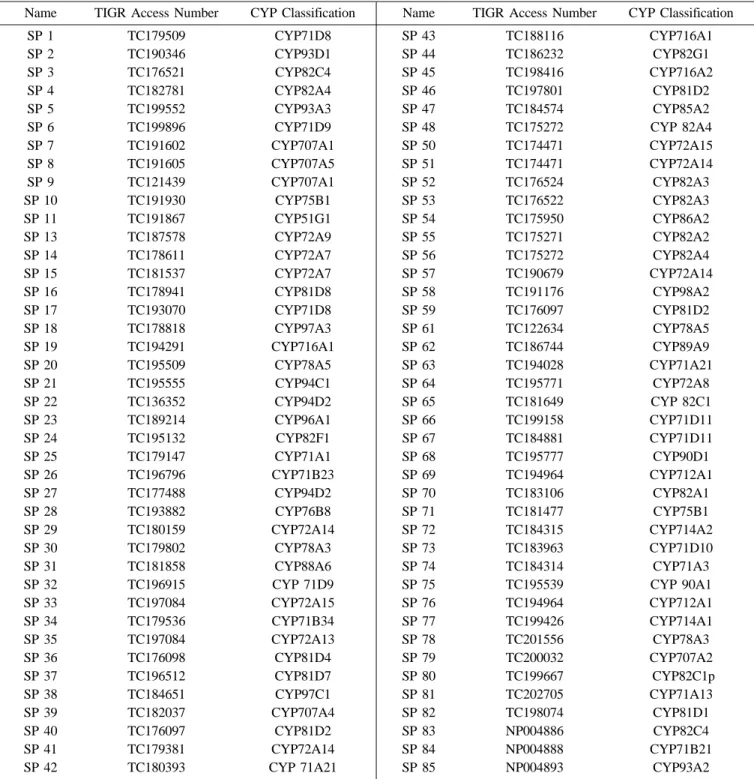
Table 1. Classification of soybean P450s
Name TIGR Access Number CYP Classification Name TIGR Access Number CYP Classification
SP 1 TC179509 CYP71D8 SP 43 TC188116 CYP716A1
SP 2 TC190346 CYP93D1 SP 44 TC186232 CYP82G1
SP 3 TC176521 CYP82C4 SP 45 TC198416 CYP716A2
SP 4 TC182781 CYP82A4 SP 46 TC197801 CYP81D2
SP 5 TC199552 CYP93A3 SP 47 TC184574 CYP85A2
SP 6 TC199896 CYP71D9 SP 48 TC175272 CYP 82A4
SP 7 TC191602 CYP707A1 SP 50 TC174471 CYP72A15
SP 8 TC191605 CYP707A5 SP 51 TC174471 CYP72A14
SP 9 TC121439 CYP707A1 SP 52 TC176524 CYP82A3
SP 10 TC191930 CYP75B1 SP 53 TC176522 CYP82A3
SP 11 TC191867 CYP51G1 SP 54 TC175950 CYP86A2
SP 13 TC187578 CYP72A9 SP 55 TC175271 CYP82A2
SP 14 TC178611 CYP72A7 SP 56 TC175272 CYP82A4
SP 15 TC181537 CYP72A7 SP 57 TC190679 CYP72A14
SP 16 TC178941 CYP81D8 SP 58 TC191176 CYP98A2
SP 17 TC193070 CYP71D8 SP 59 TC176097 CYP81D2
SP 18 TC178818 CYP97A3 SP 61 TC122634 CYP78A5
SP 19 TC194291 CYP716A1 SP 62 TC186744 CYP89A9
SP 20 TC195509 CYP78A5 SP 63 TC194028 CYP71A21
SP 21 TC195555 CYP94C1 SP 64 TC195771 CYP72A8
SP 22 TC136352 CYP94D2 SP 65 TC181649 CYP 82C1
SP 23 TC189214 CYP96A1 SP 66 TC199158 CYP71D11
SP 24 TC195132 CYP82F1 SP 67 TC184881 CYP71D11
SP 25 TC179147 CYP71A1 SP 68 TC195777 CYP90D1
SP 26 TC196796 CYP71B23 SP 69 TC194964 CYP712A1
SP 27 TC177488 CYP94D2 SP 70 TC183106 CYP82A1
SP 28 TC193882 CYP76B8 SP 71 TC181477 CYP75B1
SP 29 TC180159 CYP72A14 SP 72 TC184315 CYP714A2
SP 30 TC179802 CYP78A3 SP 73 TC183963 CYP71D10
SP 31 TC181858 CYP88A6 SP 74 TC184314 CYP71A3
SP 32 TC196915 CYP 71D9 SP 75 TC195539 CYP 90A1
SP 33 TC197084 CYP72A15 SP 76 TC194964 CYP712A1
SP 34 TC179536 CYP71B34 SP 77 TC199426 CYP714A1
SP 35 TC197084 CYP72A13 SP 78 TC201556 CYP78A3
SP 36 TC176098 CYP81D4 SP 79 TC200032 CYP707A2
SP 37 TC196512 CYP81D7 SP 80 TC199667 CYP82C1p
SP 38 TC184651 CYP97C1 SP 81 TC202705 CYP71A13
SP 39 TC182037 CYP707A4 SP 82 TC198074 CYP81D1
SP 40 TC176097 CYP81D2 SP 83 NP004886 CYP82C4
SP 41 TC179381 CYP72A14 SP 84 NP004888 CYP71B21
SP 42 TC180393 CYP 71A21 SP 85 NP004893 CYP93A2
aSP 12, SP49, and SP60 are absent due to the overlap with other SPs.
Currently, 26 full length soybean P450 genes are available in the GenBank. Upon comparison of these genes with those in TIGR, 9 complete soybean P450 genes, SP1, SP2, SP3, SP4, SP5, SP6, SP55, SP84 and SP85, existed as partial clones in TIGR. Even though we used a partial sequence for the classification, they fell into the same clan.
Only a few of full length soybean P450 genes such as isoflavone synthase24), flavone 6-hydroxylase25), and phenylurea herbicides metabolizing P45026) were characterized for their biological functions. Biological function of most of soybean P450, and other P450s from other plants are still unknown.
Biological functions of most P450s from plants including soybean are still unknown. This study might add new insight to current understanding of soybean P450s. However, other methods of analysis should be developed to understand the overall structures and the individual functions of soybean P450s. One of the best methods to accomplish this would be to express the individual P450 cDNAs in heterologuous systems able to provide the interactive P450 reductase as the electron transfer for catalytic function.
Acknowledgments
This work was supported by a grant of Biogreen 21 Program, Rural Development Administration, Republic of Korea and KRF2004-F00019 (KRF).
References
1. Chapple, C. (1998) Molecular-genetic analysis of plant cyto- chrome P450-dependent monooxygenase. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 311-343.
2. Lewis, D. F. V., and Sheridan, G. (2001) Cytochrome P450, oxygen, and evolution. Scientific World. 1, 151-167.
3. Werck-Reichhart, D., Bak, S., and Paquette S. (2002) Cyto- chrome P450. In The Arabidopsis Book, eds. Somerville, C.
R., Meyerowitz, E. M., American Society of Plant Biolo- gists, Rockville, MD.
4. Schuler, M.A., and Werck-Reichhart, D. (2003) Functional Fig. 2. Changes in soybean P450 gene expression upon UV-
irradiation. Expression levels were examined by RT-PCR.
Act is a soybean actin gene used to confirm that the same amount of RNA was used for each reaction. -, Total RNA was isolated at 0 h (just before UV-irradiation), +, Total RNA was isolated at 8 h or 12 h after UV-irradiation. Genes with dif- ferent expression upon UV-irradiation were shown. Soybean P450s that were enhanced upon UV-irradiation. (A) Soybean P450s that increased upon UV-irradiation. (B) Soybean P450s that decreased upon UV-irradiation.
Fig. 1. Expression of soybean chalcone synthase (chs) gene upon UV-irradiation.
Fig. 3. Tissue specific expression of soybean P450 genes.
Expression of 17 P450 genes in different tissues was investi- gated by RT-PCR. Actin gene was used to confirm that the same amount of RNA was used for each reaction.
genomics of P450s. Annu. Rev. Plant. Biol. 54, 629667.
5. Nelson, D. R., Schuler, M. A., Paquette, S. A., Werck- Reichhart, D., and Bak, S. (2004a) Comparative genomics of rice and Arabidopsis; analysis of 727 cytochrome P450 genes and pseudogenes from a monocot and a dicot. Plant Physiol. 135, 756-772.
6. Nelson, D. R., Zeldin, D. C., Hoffman, S. M. G., Maltais, L. J., Wain, H.M., and Nebert, D.W. (2004b) Comparison of cytochrome P450 (CYP) genes from the mouse and human genomes, including nomenclature recommendations for genes, pseudogenes, and alternative-splice variants.
Pharmacogenetics 14, 1-18.
7. Williams, .P A., Cosme, J., Sridar, V., Johnson, E., and McRee, D. E. (2002) Mammalian microsomal cytochrome P450 monooxygenase: structural adaptions for memebrane binding and functional diversity. Mol. Cell 5, 121-131.
8. Li, J., Ou-Lee T.-M., Raba, R., Amundson, R. G., and Last, R. L. (1993) Arabidopsis flavonoid mutants are hypersensi- tive to UV-B irradiation. Plant Cell 5, 171-179.
9. Harbone, J. R. B., and Williams, C. A. (2000) Advances in flavonoid research since 1992. Phytochem. 55, 481-504.
10. Bieza, K., and Lois, R. (2001) An Arabidopsis mutant toler- ant to lethal ultraviolet-B levels shows constitutively ele- vated accumultation of flavonoids and other phenolics.
Plant Physi. 126, 1105-1115.
11. Hirner, A. A., Stefan, V., and Seitz, H. U. (2001) Regula- tion of anthocyanin biosynthesis in UV-A-irradiated cell cul- tures of carrot and in organs of intact carrot plants. Plant Sci. 161, 315-322.
12. Schopfer, C. R., and Ebel, J. (1998) Identification of elici- tor-induced cytochrome P450s of soybean (Glycine max L.) using differential display of mRNA. Mol. Gen. Genet. 258, 315-322.
13. Suzuki, G., Ohta, H., Kato, T., Igarashi, T., Sakai, F., Shi- bata, D., Takano, A., Masuda, T., Shioi, Y., and Takamiya, K. (1996) Induction of a novel cytochrome P450 (CYP93 family) by methyl jasmonate in soybean suspension-cul- tured cells. FEBS Lett. 383, 83-86.
14. Jansen, M. A. K., Gaba, V., and Greenberg, B. M. (1998) Higher plants and UV-B radiation: balancing damage, repair and acclimation. Trends Plant Sci. 3, 131-135.
15. Logemann, E., Tavernaro, A., Schulz, W., Sommich, I.E., and Hahlbrock, K. (2000) UV light selectively coinduces supply pathways from primary metabolism and flavonoid secondary product formation in parsley. Pro. Natl. Acad.
Sci. 97, 1903-1907.
16. Ryan, K. G., Swinny, E. E., Markham, K. R., and Winde- filed, C. (2002) Flavonoid gene expression and UV photo-
protection in transgenic and mutant Petunia leaves.
Phytochem. 59, 23-32.
17. Schoch, G., Goepfert, S., Morant, M., Hehn, A., Meyer, D., Ullmann, P., and Werck-Reichhart, D. (2001) CYP98A3 from Arabidopsis thaliana is a 3'-hydroxylase of phenolic esters, a missing link in the phenylpropanoid pathway. J.
Biol. Chem. 276, 36566-36574.
18. Franke, R., Hemm, M.R., Denault, J.W., Ruegger, M.O., Humphreys, J.M., and Chapple, C. (2002a) Changes in sec- ondary metabolism and deposition of an unusual lignin in the ref8 mutant of Arabidopsis. Plant J. 30, 47-59.
19. Franke, R., Humphreys, J. M., Hemm, M. R., Denault, J.
W., Ruegger, M. O., Cusumano, J. C., and Chapple, C.
(2002b) The Arabidopsis REF8 gene encodes the 3-hydrox- ylase of phenylpropanoid metabolism. Plant J. 30, 33-45.
20. Holton, T. A., Brugliera, F., Lester, D. R., Tanaka, Y., Hyland, C. D., Menting, J. G., Lu, C. Y., Farcy, E., Steven- son, T. W., and Cornish, E. C. (1993) Cloning and expres- sion of cytochrome P450 genes controlling flower colour.
Nature 366, 276-279.
21. Benveniste, I., Tijet, N., Adas, F., Philipps, G., Salaun, J.P., and Durst, F. (1998) CYP86A1 from Arabidopsis thaliana encodes a cytochrome P450-dependent fatty acid omega- hydroxylase. Biochem. Biophys. Res. Com. 243, 688-693.
22. Wellesen, K., Durst, F., Pinot, F., Benveniste, I., Nettesheim, K., Wisman, E., Steiner-Lange, S., Saedler, H., and Yephre- mov, A. (2001) Functional analysis of the LACERATA gene of Arabidopsis provides evidence for different roles of fatty acid omega-hydroxylation in development. Proc. Natl.
Acad. Sci. USA. 98, 9694-9699.
23. So, E. H., Kuh, J. H., Park, K. Y., and Lee Y. H. (2001) Varietal difference of isoflavone content and antioxidant activity in soybean. Korean J. Breed. 33, 35-39.
24. Akashi, T., Aoki, T., and Ayabe, S.-I. (1999) Cloning and functional expression of a cytochrome P450 cDNA encod- ing 2-hydroxyisoflavanone synthase involved in biosynthe- sis of the isoflavonoid skeleton in licorice. Plant Physiol.
121, 821-828.
25. Latunde-Dada, A. O., Cabello-Hurtado, F., Czittrich, N., Didierjean, L., Schopfer, C., Hertkorn, N., Werck-Reich- hart, D., and Ebel, J. (2001) Flavonoid 6-hydroxylase from soybean (Glycine max L.), a novel plant P-450 monooxyge- nase. J. Biol. Chem. 276, 1688-1695.
26. Siminszky, B., Corbin, F., T., Ward, E. R., Fleischmann, T.
J., and Dewey, R. E. (1999)Expression of a soybean cyto- chrome P450 monooxygenase cDNA in yeast and tobacco enhances the metabolism of phenylurea herbicides Pro.
Natl. Acad. Sci. 96, 1750-1755.