Altered expression of mud loach (Misgurnus mizolepis;
Cypriniformes) hepcidin mRNA during experimental challenge with non-pathogenic or pathogenic bacterial species
Sang Yoon Lee, Dong Soo Kim and Yoon Kwon Nam†10)
Department of Marine Bio-Materials and Aquaculture, Pukyong National University, Busan 608-737, Korea
Transcriptional response patterns of mud loach (Misgurnus mizolepis; Cypriniformes) hepcidin, a potential ortholog to human hamp1, in response to experimental challenges with non-pathogenic and pathogenic bacterial species were analyzed based on the semi-quantitative reverse transcription-PCR assay. Mud loach hepcidin transcripts were much more preferentially induced by pathogenic bacterial species (Edwardsiella tarda and Vibrio anguillarum) causing apparent pathological symptoms than by non-pathogenic species (Escherichia coli and Bacillus thuringiensis) displaying neither clinical signs nor mortality. However in overall, the induced amounts of hepcidin transcripts were positively related with the number of bacterial cells delivered in both pathogenic and non-pathogenic bacterial species.
Inducibility of hepcidin transcripts were variable among three tissues examined (liver, kidney and spleen) in which kidney and spleen were more responsive to the bacterial challenge than liver. Time course expression patterns of hepcidin mRNAs after challenge were different between groups challenged with pathogenic and non-pathogenic species, although the overall pattern of hepcidin expression was in accordance with that generally observed in battery genes appeared during early phase of inflammation. Fish challenged with E. coli (non-pathogenic) showed the significant induction of hepcidin transcripts within 24 hr post injection (hpi) but the level was rapidly declined to the basal level either at 48 or 96 hpi. On the other hand, hepcidin transcript levels in E. tarda (pathogenic)-challenged fish were continuously elevated until 48 hpi, then downregulated at 96 hpi, although the level at 96 hpi was still significantly higher than control level observed in non-challenged fish. This expression pattern was consistent in all the three tissues examined. Taken together, our data indicate that hepcidin is tightly in relation with pathological and/or inflammation status during bacterial challenge, consequently providing useful basis to extend knowledge on the host defensive roles of hepcidin under infectious conditions in bony fish.
Key words :Bacterial challenge, Gene expression, Hepcidin, Metal exposure, Misgurnus mizolepis
Hepcidin is a central modulator for the cross-talk between innate immunity and iron regulation (Verga-Falzacappa and Muckenthaler, 2005; Viatte and Vaulont, 2009).
Although a number of previous studies have shown that the expression of fish hepcidin genes is tightly associated
†Corresponding author : Yoon Kwon Nam Tel : +82-51-629-5918 E-mail: [email protected]
with inflammation and iron overloading, several recent studies have reported that the fish hepcidin gene expressions are also modulated by other biotic and abiotic factors including estradiol-17, benzo[a]pyrene (BaP) and tetrodotoxin (Chen et al., 2008; Robertson et al., 2009; Wang et al., 2009; Matsumoto et al., 2011). Further, even the bacterial challenge-induced expression pattern of
hepcidin has been reported to differ among fish species.
Many post-mortem studies have demonstrated that hepcidin mRNAs would be highly inducible by various microbes, in which the hepcidin gene have often been activated by non-pathogenic microbes including model Escherichia coli strain as well as by identified pathogens in the host species (Huang et al., 2007; Cuesta et al., 2008; Cho et al., 2009). On the other hand, several other studies shown that hepcidin gene expression was only significantly inducible during bacterial challenge only when the challenged individuals represented clinical signs of infection (Shike et al., 2004), and also that certain bacterial challenge unexpectedly suppressed the expression of hepcidin in fish tissues (Lee et al., 2011).
Taken together, it suggests that different fish species belonging to distantly related taxonomic positions may represent functional diversification or neofunctionalization of hepcidin (or a certain isoform of multiple hepcidin copies) in their evolutionary history, although the basic dual roles of hepcidin (e.g., antimicrobial activity and iron regulation) are conserved in teleost group (Tennessen, 2005; Padhi and Verghese, 2007). Lineage-specific variation in the hepcidin gene copy numbers in teleost genomes (e.g., gene duplication or amplification of hepcidin copies only in the suborder Acanthopterygii) is also, at least in part, supportive of the potential subfunctionalization of hepcidin or hepcidin isoforms (Hilton and Lambert, 2008).
The mud loach Misgurnus mizolepis is an aquaculture-relevant, commercially important food fish in Korea with the gradually increasing market demand.
This species also has many attractive merits as a candidate experimental model organism due to its
easiness of laboratory rearing, relatively short generation time, high fecundity and well-established genetic manipulation techniques (Nam et al., 2000). Based on the hypothesis by Hilton and Lambert (2008), this cypriniform species belongs to non-Acanthopterygii superorder and hence may possess only a single copy of hepcidin that has an orthology to human hamp1.
Consequently the mud loach hamp1 is proposed to acquire multivalent or multifunctional functions as similarly with human ortholog. In line with our long-term goal to develop mud loach as a comparative model system for studying hepcidin function in vertebrates, the objective of this study is to examine the differential expression pattern of hepcidin depending on the pathogenicity of selected bacterial species used for experimental challenges.
Materials and Methods
Fish maintenance and hepcidin cDNA
Mud loach individuals used in this study were laboratory-bred strains. Artificial insemination according to the method described by Nam et al. (2000) was performed and reared in water-recirculating system.
Detailed conditions for fish rearing and maintenance can be referred to Nam et al. (2000). All the bacterial challenge experiments in this study were conducted at 25 °C using 10-μm filtered tap water. Fish were not fed during bacterial challenge. Mud loach hepcidin cDNA was isolated from the mud loach expressed sequence database (unpublished data) and its full-length sequence encoding a polypeptide of 91 amino acids was confirmed by direct sequencing of RT-PCR product.
The sequence was deposited in NCBI GenBank under the accession number JN230708.
Experimental bacterial challenge
Preliminary experiments to test potential pathogenicity of different bacterial species in mud loach were performed by intraperitoneal injection (5 × 106 cells/g body weight) followed by the examination of any clinical signs for infectious diseases (such as hemorrhage, swelling anus and/or impaired swimming ability) and occurrence of mortality until 2 weeks post injection.
Bacterial strains tested were Bacillus thuringiensis (Gram +; ATCC 33679), Lactococcus garvieae (Gram +;
ATCC 49156), Streptococcus iniae (Gram +; JSL0108), or S. parauberis (Gram +; KCTC 3651), Escherichia coli (Gram-; XL1 Blue MRFʹ; Stratagene, USA), Edwardsiella tarda (Gram-; FSW910410), E. tarda (Gram-; direct isolate from Paralichthys olivaceus), Vibrio anguillarum (Gram-; KFCC-11377P), V. harveyi (Gram-; ATCC 14126) and V. vulnificus (Gram-; ATCC 27562). Based on the examinations of clinical signs and mortality, the bacterial strain showing most potent pathogenicity was proven to be E. tarda (FSW910410) which cause the hemorrhage sign in most individuals injected as well as the cumulative mortality higher than 50% until 1 week after injection. Injection of V.
anguillarum also resulted significant mortality (up to 30% at 1 week post injection) with clinical signs in many individuals, although the severity was not as high as in E. tarda-injected individuals. On the other hand, the individuals E. coli, B. thuringiensis and S. iniae showed neither clinical sign nor mortality. Remaining other bacterial strains result in variable degrees of signs
but was difficult to be clearly classified into distinct groups in a quantitative manner. From this preliminary observation, the E. tarda and V. anguillarum were chosen as model strains of pathogenic microbes in mud loach, while E. coli and B. thuringiensis were selected as models of non-pathogenic microbes.
Firstly, the four selected bacterial species (two pathogenic and two non-pathogenic) were freshly grown in appropriate medium as reported previously and healthy mud loach individuals (n = 12; average body weight = 14.6 ± 2 g) were re-challenged with one of the four bacterial species at the dose level of 5 × 106 cells/g body weight. Non-challenged control (n = 12;
same-sized above) was prepared by injecting only a suspension medium (0.85% NaCl). Injection volume was adjusted to be 150 μl. After injection, each group was transferred to 60 L tank containing 50 L tap water at 25 °C. At 48 h post injection, liver, kidney and spleen were surgically removed from five-randomly chosen individual for RT-PCR assay in order to examine if the apparent pathogenicity of each bacterial species is in relation with the inducibility of hepcidin gene expression in different organs. Unsampled individuals in each group kept to estimate the cumulative mortality until 1 week. Two replicate examinations were made for each group.
Secondly, based on the result of the first experiment, the effects of different bacterial burdens of non-pathogenic and pathogenic bacterial strains on the induced expression of hepcidin were examined. In this challenge, E. coli and E. tarda were selected as non-pathogenic and pathogenic model strains, respectively. For each bacterial species, three bacterial doses (1 × 105 cells/g,
1 × 106 cells/g and 1 × 107 cells/g) were delivered to mud loach individuals (n = 8; same-sized above) via intraperitoneal injection. Non-challenged control was also prepared using a saline-injection, and the tank conditions were the same as described above. Twenty-four hours after injection, the three organs (liver, kidney and spleen) were obtained for RT-PCR assay.
Thirdly, time course of hepcidin expression was compared between two groups challenged with either E. coli (non-pathogenic) or E. tarda (pathogenic) strain.
Eight individuals were intraperitoneally injected with a 150 μl of bacterial suspension containing E. coli or E. tarda. The bacterial number was fixed to be 1 × 106 cells/g body weight for both bacterial species.
Non-challenged control was prepared by injection of the equal volume of saline alone. Again, the liver, kidney and spleen were sampled from each individual at 24, 48 and 96 h post injection for RT-PCR analysis.
Semi-quantitative RT-PCR analysis
Total RNA was extracted and purified using the RNeasy Midi Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The integrity of total RNA in each sample was checked using 28S : 18S rRNA ratios in ethidium bromide-stained agarose gel. Two μg of total RNA aliquot from each sample was reverse transcribed into cDNA using the Omniscript® Reverse Transcription Kit (Qiagen) with an with an oligo-d(T)20
primer (1 M final concentration) and a mud loach 18S rRNA reverse primer (ML 18S-RV; 5ʹ -CAAGAATTTCACCTCTAGCGGC-3ʹ; 0.1 M). Aliquots (2 L) of diluted cDNA (diluted fourfold for hamp1 and 20-fold for 18S rRNA) were subjected to thermal
amplification. Based on the preliminary experiment to optimize the cycling numbers of hepcidin and 18S rRNA genes in different tissues, the hepcidin cDNA fragment (263 bp) and 18S rRNA fragment (274 bp) was amplified with oligonucleotide primer pairs qMLHAMP1 1F (5ʹ -CAAACTGCTGCGTCTCCATT-3ʹ) / qMLHAMP1 1R (5ʹ -CAACTTGGGAAAAGCAGCTC-3ʹ) and qML18S 1F (5ʹ -ACCCATTGGAGGGCAAGTCT-3ʹ) / qML18S 1R (5ʹ -CCTAGCTGAGATATTCAGGC-3ʹ), respectively. Thermal conditions for both genes were as follow: 94 °C for 20 s, 58 °C for 20 s and 72 °C for 20 s with an initial denaturation step at 94 °C for 2 min. Cycle numbers of hepcidin were 28 for kidney and spleen, while 24 for liver. On the other hand, the cycle number of 18S rRNA was fixed to be 20 irrespective of tissue types.
Five μL of amplified product was separated in 1.5%
agarose gel and visualized by ethidium bromide staining.
Three independent RT-PCR assays were performed and the expression levels were determined by densitometry using Quantity-OneTM software implemented in the VersaDoc 4000 (Bio-Rad, Hercules, CA, USA).
Statistical evaluation
The hepcidin mRNA level in each sample was pre-normalized against its own level of 18S rRNA control, then the expression levels of hepcidin in bacterial challenged groups relative to non-challenged (i.e., saline-injected) controls were estimated as fold change, according to the method described previously (Schmittgen and Livak, 2008). Triplicate assays were performed in an independent fashion. The hepcidin expression levels between or among groups were assessed by ANOVA followed by Duncan’s multiple range tests. Difference
were considered to be significant when p < 0.05.
Results and Discussion
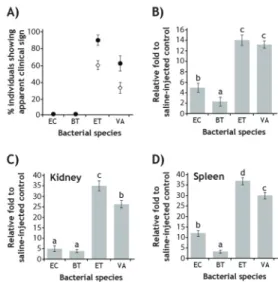
Based on the semi-quantitative RT-PCR assay, the hepcidin expression was proven to be significantly affected by the potential pathogenicity of the bacterial species in mud loaches (Fig. 1). Experimental challenges with the two presumed non-pathogenic bacterial species (E. coli and B. thuringiensis) did not show any apparent clinical signs of infectious disease and there was no mortality up to 1 week post-challenge. However, the two potential pathogens (E. tarda and V. anguillarum) resulted in diverse signs of infections, which could be detectable at least 80%
and 60% of individuals challenged. Considerable portions of individuals showing clinical signs were ultimately dead within 1 week. Regardless of the three tissues (liver, kidney and spleen) examined, the induced amounts of hepcidin transcripts were always higher in the groups challenged with E. tarda or V. anguillarum than in the groups with E. coli or B. thuringiensis. Moreover, the group challenged with E. tarda, of which viability and physiology were most adversely influenced during the challenge, exhibited more i nduct i on of hepci di n t ranscri pt s t h a n V . anguillarum-injected fish did. It suggests that the inducibility of hepcidin gene expression is closely associated with the degree of pathological progress and/or inflammation status (Rodrigues et al., 2006; Huang et al., 2007). In comparison among three tissues, the induced folds of hepcidin transcripts in groups injected with E.
tarda or V. anguillarum were significantly greater in kidney and spleen than in liver, which is congruent with previously proposed explanation that hepcidin expression in the liver
would play major role for negative iron regulation while hepcidin expression in other tissues support mainly for the innate immunity and host defense (Rodrigues et al., 2006; Cho et al., 2009). Meanwhile, the inducibility of hepcidin is higher in E. coli-challenged than in B.thuringiensis-challenged groups, suggesting the possibility that mud loach hepcidin might also be differentially responsive to bacterial Gram (-) or Gram (+)
Fig. 1. Effects of bacterial pathogenicity on the mRNA expression of hepcidin in mud loach tissues. (A) Incidence (%) of clinically affected individuals (closed circles) and cumulative mortality (open diamonds) assessed at 1 week post bacterial challenge using non-pathogenic [Escherichia coli (EC) and Bacillus thuringiensis (BT)] or pathogenic [Edwardsiella tarda (ET) and Vibrio anguillarum (VA)].
(B-D) Differential expression of hepcidin transcripts in liver (B), kidney (C) and spleen (D) during bacterial challenges as judged by semi-quantitative RT-PCR assays in triplicates.
Expression levels of hepcidin mRNAs in bacterial challenged groups are expressed as fold induction relative to that of saline-injected control, after prenormalization against 18S rRNA standard. Mean ± SDs with the same letters are not significantly different based on ANOVA followed Duncan’s multiple range tests (p < 0.05)
characteristics. Our finding on the relationship between hepcidin induction and potential pathogenicity is in concordance with to previous data made with zebrafish hepcidin, in which significant elevation of hepcidin mRNA was only observed in clinically affected individuals but not in healthy individuals after bacterial challenge (Shike et al., 2004).
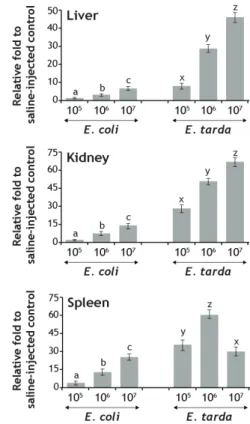
Different bacterial burdens resulted in differential amount of induced hepcidin transcripts in mud loach tissues wherein the overall pattern was toward upregulation with increasing bacterial numbers (Fig. 2).
It is consistent with not only pathogenic bacterial species (E. tarda) but also non-pathogenic species (E. coli).
However as expected, the inducibility of hepcidin transcripts was significantly higher in the fish injected with E. tarda than those with E. coli. Clear dose dependency of hepcidin expression was observable in both liver and kidney irrespective of the pathogenicity of the bacterial species. On the other hand, the fish injected with the highest number of E. tarda showed the least inducibility of hepcidin transcripts in the spleen, which is notably different from the patterns observed in other two tissues. Although suppressive regulation of hepcidin gene during bacterial challenge has not been frequently reported, our finding is similar with a recent observation on the significant downregulation of redbanded seabream (Pagrus auriga) hepcidin in the spleen by LPS challenge (Martin-Antonio et al., 2009) as well as the examination on the suppression of one of two hepcidin isoforms in Java ricefish (Oryzias javanicus) liver when the fish were exposed to E. tarda via immersion (Lee et al., 2011). Previous and present study may propose that high bacterial burdens beyond
a certain level may interfere or evade the host defense system along with severe cytotoxic effects and cell death, consequently resulting in the decreased expression of hose defense genes (Rodriguez et al., 2008; Cho et al., 2009).
Fig. 2. Effects of bacterial cell numbers on the mRNA expression of mud loach hepcidin in liver, kidney and spleen. Non-pathogenic Escherichia coli and pathogenic Edwardsiella tarda were delivered into fish in varying doses (1 × 105, 1 × 106 and 1 × 107 cells/fish with an average body weight of 15 g) and hepcidin expression was assayed by semi-quantitative RT-PCR at 24 hr post injection.
Mean ± SDs with the same letters (a-c for E. coli and x-z for E. tarda) are not significantly different based on ANOVA followed Duncan’s multiple range tests (p < 0.05).
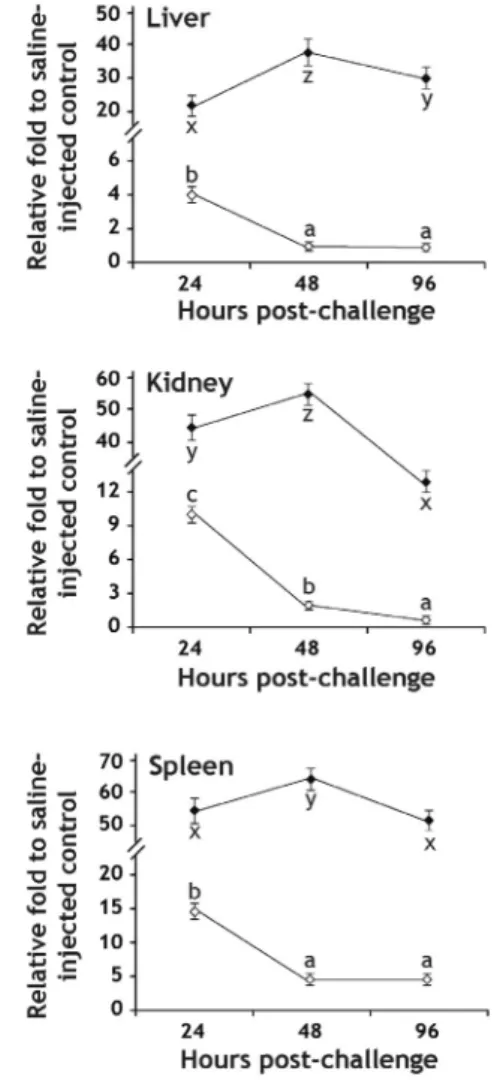
Time course expression assay of hepcidin transcripts after injection of E. coli or E. tarda represented the
expression pattern, which is in accordance with that of early battery genes during immune stimulation (Zhou et al., 2011) (Fig. 3). However the expression pattern was not identical between E. coli-and E. tarda-challenged groups. Generally in the group injected with E. coli, the significant amount of induced hepcidin transcripts at 24-h post injection was rapidly downregulated at 48-h post injection and then stabilized or further decreased at 96-h post injection. Except in the spleen, the expression levels of hepcidin in the liver and kidney of the E. coli-injected fish at these detection points were not significantly different from those in non-challenged fishes, suggesting that the non-pathogenic bacteria could elicit very early phase response but the effect is immediately diminished soon. However unlike E.
coli-injected groups, the hepcidin transcripts in the E.
tarda-challenged group were further elevated at 48-h post injection and then decreased down at 96-h post injection, although hepcidin transcript levels at 96-h were still significantly higher than the basal expression levels observed in non-challenged fishes. This pattern is consistent with all of three tissues examined. Data from this experimental challenge suggest that inflammatory status might proceed and/or retain at least until four days after challenge in the E. tarda-injected group, and also that hepcidin expression shows a close interrelationship with pathological progress and inflammation-mediated stress in fish. To gain a deeper insight into the regulation of hepcidin gene, genomic information on the 5ʹ-regulatory region of mud loach hepcidin, particularly regarding the transcription factor binding motifs, should be deciphered (Cho et al., 2009;
Darshan and Anderson, 2009). Also the comparative
assay of hepcidin mRNA along with cytokine gene expressions may be valuable for validating the inflammation-mediated regulation of the hepcidin in different mud loach tissues.
Fig. 3. Time course expression of hepcidin transcripts in mud loach tissues up to 96 hr after bacterial challenge with either Escherichia coli (open diamonds) or Edwardsiella tarda (closed diamonds), as determined by semi-quantitative RT-PCR assay. Mean ± SDs with the same letters (a-c for E. coli and x-z for E. tarda) are not significantly different based on ANOVA followed Duncan’s multiple range tests (p < 0.05).
In summary, mRNA expression patterns of mud loach hepcidin, a potential ortholog to human hamp1, were examined in response to experimental challenges with potentially non-pathogenic or pathogenic bacterial species. Mud loach hepcidin gene is much more responsive to the pathogenic bacterial invaders causing apparent pathological symptoms than to non-pathogenic bacterial species representing neither significant clinical sign nor mortality. Induced expression pattern of hepcidin is well in relation with the bacterial burden for both pathogenic and non-pathogenic species. Time course pattern of hepcidin expression after challenge is closely associated also with pathological progress and inflammation status depending on pathogenicity of the bacterial species. Data form this study would be a useful basis for extending our knowledge on the hose defensive roles of hepcidins in pathogen-infected fish.
Acknowledgment
This study was supported by a research fund from the Ministry of Land, Transport and Maritime Affairs, Republic of Korea (Project #20088033-1).
References
Chen, J., Shi, Y.H. and Li, M.Y.: Changes in transferrin and hepcidin genes expression in the liver of the fish Pseudosciaena crocea following exposure to cadmium. Arch. Toxicol., 82:525-530, 2008.
Cho, Y.S., Lee, S.Y., Kim, K.H., Kim, S.K., Kim, D.S.
and Nam, Y.K.: Gene structure and differential modulation of multiple rockbream (Oplegnathus
fasciatus) hepcidin isoforms resulting from different biological stimulations. Dev. Comp.
Immunol., 33:46-58, 2009.
Cuesta, A., Meseguer, J. and Esteban, M.Á.: The antimicrobial peptide hepcidin exerts an important role in the innate immunity against bacteria in the bony fish gilthead seabream. Mol. Immunol., 45:2333-2342, 2008.
Darshan, D. and Anderson, G.J.: Interacting signals in the control of hepcidin expression. Biometals, 22:77-87, 2009.
Hilton, K.B. and Lambert, L.A.: Molecular evolution and characterization of hepcidin gene products in vertebrates. Gene, 415:40-48, 2008.
Huang, P.H., Chen, J.Y. and Kuo, C.M.: Three different hepcidins from tilapia, Oreochromis mossambicus:
Analysis of their expressions and biological functions. Mol. Immunol., 44:1922-1934, 2007.
Lee, S.Y., Kim, B.S., Kim, D.S. and Nam, Y.K.: Isolation of two hepcidin paralogs, hamp1 and hamp2 from a euryhaline Javanese ricefish (Oryzias javanicus:
Beloniformes). Fish. Aquat. Sci., 14:93-104, 2011.
Martin-Antonio, B., Jiménez-Cantizano,R.M., Salas-Leiton, E., Infante, C. and Manchado, M.: Genomic characterization and gene expression analysis of four hepcidin genes in the redbanded seabream (Pagrus auriga). Fish Shellfish Immunol., 26:483-491, 2009.
Matsumoto, T., Ishizaki, S. and Nagashima, Y.: Differential gene expression profile in the liver of the marine puffer fish Takifugu rubripes induced by intramuscular administration of tetrodotoxin.
Toxicon, 57:304-310, 2011.
Nam, Y.K., Cho, Y.S. and Kim, D.S.: Isogenic transgenic
homozygous fish induced by artificial parthenogenesis.
Transgenic Res., 9:463-469, 2000.
Padhi, A. and Verghese, B.: Evidence for positive Darwinian selection on the hepcidin gene of perciform and pleuronectiform fishes. Mol. Divers., 11:119-130, 2007.
Robertson, L.S., Iwanowicz, L.R. and Marranca, J.M.:
Identification of centrarchid hepcidins and evidence that 17β-estradiol disrupts constitutive expression of hepcidin-1 and inducible expression of hepcidin-2 in largemouth bass (Micropterus salmoides). Fish Shellfish Immunol., 26:898-907, 2009.
Rodrigues, P.N., Vázquez-Dorado, S., Neves, J.V. and Wilson, J.M.: Dual function of fish hepcidin: Response to experimental iron overload and bacterial infection in sea bass (Dicentrarchus labrax). Dev. Comp.
Immunol., 30:1156-1167, 2006.
Rodriguez, I., Novoa, B. and Figueras, A.: Immune response of zebrafish (Danio rerio) against a newly isolated bacterial pathogen Aeromonas hydrophila. Fish Shellfish Immunol., 25:239-249, 2008.
Schmittgen, T.D. and Livak, K.J.: Analyzing real-time PCR data by the comparative CT method. Nat. Protoc., 3:1101-1108, 2008.
Shike, H., Shimizu, C., Lauth, X. and Burns, J.C.: Organization and expression analysis of the zebrafish hepcidin
gene, an antimicrobial peptide gene conserved among vertebrates. Dev. Comp. Immunol., 28:747-754, 2004.
Tennessen, J.A.: Molecular evolution of animal antimicrobial peptides: widespread moderate positive selection.
J. Evol. Biol., 18:1387-1394, 2005.
Verga-Falzacappa, M.V. and Muckenthaler, M.U.: Hepcidin:
Iron-hormone and anti-microbial peptide. Gene, 364:37-44, 2005.
Viatte, L. and Vaulont, S.: Hepcidin, the iron watcher.
Biochimie, 91:1223-1228, 2009.
Wang, K.J., Cai, J.J., Cai, L., Qu, H.D., Yang, M. and Zhang, M.: Cloning and expression of a hepcidin gene from a marine fish (Psedosciaena crocea) and the antimicrobial activity of its synthetic peptide. Peptides, 30:638-646, 2009.
Zhou, J.G., Wei, J.G., Xu, D., Cui, H.C., Yan, Y., Ou-Yang, Z.L. et al. Molecular cloning and characterization of two novel hepcidins from orange-spotted grouper, Epinephelus coioides. Fish Shellfish Immunol., 30:559-568, 2011.
Manuscript Received : September 7, 2011 Revised : December 4, 2011 Accepted : December 5, 2011